不同作物品種耐鋁毒基因型篩選試驗(yàn)
2017-10-28 23:15:27邢承華劉仲書蔣紅英
現(xiàn)代農(nóng)業(yè)科技 2017年18期
邢承華+劉仲書+蔣紅英


摘要 采用營養(yǎng)液培養(yǎng)的方法,對水稻23個(gè)品種、大豆14個(gè)品種和苜蓿12個(gè)品種進(jìn)行了耐鋁性篩選試驗(yàn),并對各品種的耐鋁性進(jìn)行了評價(jià)和對各個(gè)品種根部和株高生長狀況進(jìn)行了比較,得出耐鋁性和鋁敏感性品種。結(jié)果表明,耐鋁基因型具有植株相對生物量較高、種子根相對伸長率、蘇木精著色程度較低的共同特點(diǎn)。種子根相對伸長率、蘇木精染色指數(shù)與植株的耐鋁性具有顯著的相關(guān)性。鋁脅迫下耐鋁性不同的典型植物的根系形態(tài)具有明顯差異,耐鋁基因型明顯優(yōu)于鋁敏感基因型。
關(guān)鍵詞 水稻;苜蓿;大豆;品種;耐鋁性評價(jià);根系形態(tài)
中圖分類號 S511;S557.1;S565.1 文獻(xiàn)標(biāo)識碼 A 文章編號 1007-5739(2017)17-0034-05
Al-tolerance Genotype Screening About Different Plants
XING Cheng-hua 1 LIU Zhong-shu 2 JIANG Hong-ying 1
(1 Agricultural and Bioengineering Institute,Jinhua Polytechnic,Jinhua Zhejiang 321007; 2 College of Chemistry and Life Sciences,
Zhejiang Normal University)
Abstract Using nutrition training methods,Al-tolerance test of 23 rice varieties,14 soybean varieties and 12 alfalfa varieties were did to evaluate the various species of Al-tolerance and contraste their root growth and plant height.The results showed that the Al-tolerant lines had lower hexnatoxylin dye index,higher RSRL and higher relative biomass in comparison with Al-sensitive lines.There were significant correlations between the relative seminal root length(RSRL),hematoxylin dye index,aril the Al-tolerance of the plant. There were significant difference of morphological and anatomical characteristics different patterns typical of the root system,Al-tolerant genotypes were distinctly better than Al-sensitive ones.
Key words rice;alfalfa;soybean;varieties;Al-tolerance estimation;root morphological
據(jù)估計(jì),酸性土壤占世界可耕地的40%,主要分布在發(fā)展中國家集中的熱帶和亞熱帶。我國酸性土壤遍及南方15個(gè)省(區(qū)),總面積達(dá)2 030萬km2。在酸性土壤上影響作物生長的因素包括金屬陽離子濃度的降低和吸收的抑制(Mg、Ca、K的缺乏)、氫離子、鋁毒、錳毒、磷和鋁可溶性的降低、水分缺乏和淋溶作用增強(qiáng)(養(yǎng)分缺乏)。而鋁毒被認(rèn)為是限制酸性上壤上作物生長的主要因素。鋁是地殼中含量最豐富的金屬元素。在大多數(shù)情況下,土壤中的鋁是無毒性的。然而,在酸性土壤條件下,對大多數(shù)植物都會(huì)產(chǎn)生毒害作用。鋁對生物體顯示出特殊的毒性是與鋁存在的化學(xué)形態(tài)緊密相連的。一般認(rèn)為,配合態(tài)鋁和固定態(tài)的鋁(稱低活性鋁)對植物和環(huán)境沒有毒害,只有離子態(tài)的鋁(僅需在微摩爾濃度下)才對環(huán)境和植物產(chǎn)生影響。其中,無機(jī)離子態(tài)鋁如Al3+和Al(OH)2+(稱高活性鋁)對植物根系的毒害作用最大,有機(jī)和無機(jī)鋁化復(fù)合物如Al(OH)3、Al(OH)4-、磷酸鋁復(fù)合體、氧化鋁復(fù)合體、有機(jī)酸鋁等,其毒性較小。許多人也注意到,環(huán)境中鋁的形態(tài)對植物根的吸收有很大影響。當(dāng)含鋁的溶液與強(qiáng)堿發(fā)生中和反應(yīng)時(shí)溶液中會(huì)產(chǎn)生有劇毒的Al3+聚合物。因此,研究植物耐鋁毒的機(jī)制已成為生物科學(xué)界一項(xiàng)重要而熱門的問題。
目前有關(guān)植物耐鋁毒的機(jī)理已有報(bào)道[1],其尤其在酸雨降沉頻率較高、人們長期施用酸性化肥的地區(qū),土壤中與氧和硅結(jié)合的鋁易轉(zhuǎn)變?yōu)橛卸镜碾x子態(tài)Al3+ [2],且Al3+很容易在土壤中發(fā)生遷移,造成大面積土壤酸化[3]。鋁通常是以難溶性硅酸鹽或氧化鋁的形式存在,對植物沒有毒害[4],但pH<5時(shí),這些難溶性的物質(zhì)溶解,可溶性的鋁(主要是Al3+)對大多數(shù)植物都會(huì)產(chǎn)生毒害[5]。鋁毒的最初癥狀是抑制根系伸長,影響植株的物質(zhì)和能量代謝[6]。重復(fù)施用含鋁量較高的等化學(xué)肥料,可以導(dǎo)致土壤鋁積累,土壤被鋁污染后可引起作物尤其是植物體內(nèi)鋁的積累。稻米、大豆作為人類食物的直接來源,是人體攝入鋁的主要途徑,高含鋁量的稻米、大豆或者苜蓿通過食物鏈會(huì)嚴(yán)重危害人體健康,引起中毒或老年癡呆癥等癥狀。隨著我國經(jīng)濟(jì)的不斷發(fā)展,污水灌溉、污泥、農(nóng)藥及肥料的施用等導(dǎo)致土壤重金屬污染日趨嚴(yán)重,解決植物的污染問題迫在眉睫。鋁的污染也不例外。鋁的積累性很高,受鋁毒害的人越來越多,攝入過多的鋁會(huì)破壞某些酶的活性,引起消化功能紊亂,破壞正常鈣磷比。在鋁污染土壤上通過適當(dāng)?shù)姆椒ㄏ拗其X從土壤進(jìn)入植物根系、控制植物對鋁的吸收是十分必要的。本課題主要研究同一植物種類不同品種間的耐鋁能力存在明顯的基因型差異。
本課題以苜蓿、大豆、水稻的多個(gè)品種為材料,研究其在生長過程中受鋁的脅迫導(dǎo)致生理形態(tài)的變化情況,以及受鋁脅迫后植物根尖鋁的含量,并選出同類植物中耐鋁能力較好的植物品種,為有效解決酸性土壤中的鋁毒害提供有效途徑,并為后續(xù)的通過轉(zhuǎn)基因技術(shù)培育耐鋁品種奠定基礎(chǔ)。
1 材料與方法
1.1 供試作物品種及來源
試驗(yàn)中共篩選23個(gè)水稻品種、12個(gè)苜蓿品種、14個(gè)大豆品種。
供試水稻品種:Ⅱ優(yōu)92、菲優(yōu)多系1號、Ⅱ優(yōu)838、德龍2000、Ⅱ優(yōu)3027、富優(yōu)1號、Ⅱ優(yōu)2070、協(xié)優(yōu)92、Ⅱ優(yōu)6216、Ⅱ優(yōu)8220、明優(yōu)98、Ⅱ優(yōu)46、協(xié)優(yōu)9019、Ⅱ優(yōu)63、協(xié)優(yōu)982、協(xié)優(yōu)9308、金優(yōu)987、e富豐優(yōu)11、紅良優(yōu)166、協(xié)優(yōu)63、Ⅱ優(yōu)8006、Ⅱ航優(yōu)148、協(xié)優(yōu)46。
供試苜蓿品種:阿爾岡金、巨人201、金皇后、WL-323HQ、WL-525HQ、周產(chǎn)紫花苜蓿、皇冠、飛馬、馴虎、WL-232HQ、胖多、獵人河。
供試大豆品種:矮腳青8901、引豆9707、春豐早、臺灣毛豆、日本青、科源1號、遼新1號、科源3號、028-289、浙秋2號、0290-236、029-15、029-238、029-290。水稻和大豆從金華種子公司購買,苜蓿從北京種子公司購買。
1.2 育苗培養(yǎng)與處理
種子用1%次氯酸鈉浸泡消毒30 min后,蒸餾水清洗6次,每次清洗5 min,以除去漂浮、膨脹、軟化和被污染的種子,然后在蒸餾水中浸泡(苜蓿24 h、大豆4 h、水稻24 h),將種子轉(zhuǎn)入培養(yǎng)皿內(nèi),倒掉蒸餾水,但要使種子保持濕潤,讓種子繼續(xù)催芽。待種子根長長到1 cm左右時(shí),繼續(xù)使種子生長,待種子根長長到2 cm左右時(shí),可以將其移入盆中對照培養(yǎng)。本試驗(yàn)是在浙江師范大學(xué)的實(shí)驗(yàn)室培養(yǎng)箱中培養(yǎng),水稻、苜蓿、大豆的培養(yǎng)條件都有所不同。其中水稻的培養(yǎng)條件為白天14 h、30 ℃,夜間10 h、25 ℃,濕度為70%,苜蓿的培養(yǎng)條件為白天14 h、25 ℃,夜間10 h、20 ℃,濕度為70%,而大豆的培養(yǎng)條件為黑夜24 h、25 ℃,濕度為70%。選取根長相近的幼苗在外面包有2層黑色塑料薄膜的盆中進(jìn)行隔離培養(yǎng)處理,盆內(nèi)培養(yǎng)24株幼苗,溶液體積為1 L,在盆上放一泡沫,在每個(gè)泡沫上打24個(gè)洞,每個(gè)洞內(nèi)移植1株幼苗。對照組的溶液氯化鈣的濃度為0.5 mmol/L,而試驗(yàn)組的溶液氯化鋁的濃度和氯化鈣的濃度分別為0.05 mmol/L和0.5 mmol/L。所有溶液的pH值用HCl調(diào)到4.5(由于苜蓿本身營養(yǎng)不是很豐富,故苜蓿用營養(yǎng)液培養(yǎng),其中試驗(yàn)組要另外加氯化鋁,營養(yǎng)液中氯化鋁的含量為0.05 mmol/L)。營養(yǎng)液配制如表1、2所示。
1.3 測定內(nèi)容與方法
1.3.1 植株生長量測定。植株生長2 d后測定對照組和Al處理的試驗(yàn)組植株的株高和根長度變化,以及株高、根長的增加量。并計(jì)算相對根伸長量和相對株高伸長量。根系相對伸長量可用于估計(jì)耐鋁的基因型差異。計(jì)算方法如下:
根相對伸長量(%)=試驗(yàn)組的根伸長量/對照組的根伸長量×100;
株高相對伸長量(%)=試驗(yàn)組的株高伸長量/對照組得株高伸長量×100;
相對Al含量(%)=試驗(yàn)組的OD值/對照組OD值×100。
1.3.2 根尖相對鋁含量的測定。經(jīng)過培養(yǎng)48 h后,試驗(yàn)組用氯化鈣溶液沖洗,去除植物根部表面上的Al,再用蒸餾水清洗,對照組直接清洗,洗后切下2 cm的根尖,用0.5 mmol/L 0.2%的蘇木精和0.02%的KIO3混合液染色15 min,用蒸餾水洗脫15 min,對照組和試驗(yàn)組的根各分3組(每組8根),經(jīng)3 mL 1 mol/L的HCl脫色1 h后,用UV-7504紫外可見分光光度計(jì)在490 nm下測植物根尖的分光光度值(這步非常重要,也是估計(jì)耐鋁的基因型差異的關(guān)鍵一步)。
2 結(jié)果與分析
2.1 對水稻幼苗根長和株高及根尖相對鋁含量的影響
水稻幼苗經(jīng)50 μmol/L的鋁處理后,鋁對水稻株高、根長的影響不同。在Al濃度為50 μmol/L時(shí),對水稻幼苗株高和根長都有影響。對根長的抑制作用則更大(圖1)。孔繁翔等[7]、郝魯寧等[8]認(rèn)為,鋁對植物毒害作用最明顯的特征是抑制了根尖細(xì)胞伸長和細(xì)胞分裂,根比地上部分對鋁更敏感。本試驗(yàn)也反映低濃度鋁處理時(shí),水稻幼苗受鋁的影響,根較植株受鋁的影響要更顯著,鋁對根有較大的抑制。然而有極少數(shù)品種在低濃度下,株高呈現(xiàn)刺激植物的生長。秦瑞君等[9]指出,少量鋁的存在可刺激植物的生長。本試驗(yàn)也證實(shí)了這一觀點(diǎn)(圖2)。
由圖1可知,富優(yōu)1號、Ⅱ優(yōu)63、協(xié)優(yōu)9308和紅良優(yōu)166這幾個(gè)品種的根相對伸長率比較低,說明這幾個(gè)品種受Al的影響比較大,受Al毒的抑制造成的,而根相對伸長率比較高的品種則是菲優(yōu)多系1號和Ⅱ優(yōu)838,說明這2個(gè)品種受Al毒的影響比較小,這也正是本試驗(yàn)篩選所要的品種。通過Spass分析可知,水稻各品種的根相對伸長率存在顯著性差異。
由圖2可知,株高相對的伸長率基本都在60%以上波動(dòng),只有Ⅱ優(yōu)3027、富優(yōu)1號品種的株高受Al毒的影響比較大,而Ⅱ優(yōu)8220、明優(yōu)98和Ⅱ優(yōu)8006的株高相對伸長率則出現(xiàn)大于100%,可能是由于低濃度的Al反而刺激了株高的伸長。結(jié)合圖1和圖2可知,富優(yōu)1號受Al毒的影響很大,對其株高和根的抑制都很大,該水稻品種則不適于在酸性土壤種植,且該水稻品種中的Al含量可能大于其他品種的水稻。
試驗(yàn)對23個(gè)水稻品種的根部染色,測得它們的相對Al含量,如圖3所示。菲優(yōu)多系1號、Ⅱ優(yōu)838、協(xié)優(yōu)92、Ⅱ優(yōu)6216、明優(yōu)98、協(xié)優(yōu)9019這5個(gè)品種的相對Al含量最小,說明它們根尖的鋁含量較低,是比較耐鋁的品種,而富優(yōu)1號、Ⅱ優(yōu)46、協(xié)優(yōu)9308和e富豐優(yōu)11的相對Al含量較大,則說明試驗(yàn)的較對照的易吸收鋁,試驗(yàn)較對照根尖的鋁含量多,因此它們是不耐鋁的品種,即不是所要選擇的品種。富優(yōu)1號、Ⅱ優(yōu)46、協(xié)優(yōu)9308和e富豐優(yōu)11的相對Al含量都是Ⅱ優(yōu)838的4倍左右,因此這2個(gè)品種水稻不適于種植在酸性的土壤中,農(nóng)民盡量不要種這種水稻,因?yàn)檫@種水稻含有很多的鋁,對人的身體有害,長時(shí)間食用鋁導(dǎo)致其在人身體內(nèi)積累,也許會(huì)破壞人體的內(nèi)分泌系統(tǒng),損傷神經(jīng),甚至?xí)?dǎo)致老年癡呆癥。結(jié)合圖1、圖2和圖3所得出的結(jié)論,可知菲優(yōu)多系1號、Ⅱ優(yōu)838是所有篩選品種中耐鋁性最好的,這為廣大的農(nóng)民種植何種水稻提供了有效的參考。通過Spass分析可知,水稻各品種的株高相對伸長率和根尖的相對Al含量存在顯著性差異。
2.2 對苜蓿幼苗根長和株高及根尖相對鋁含量的影響
苜蓿幼苗經(jīng)過處理后各個(gè)品種對照和試驗(yàn)的根長都有一定的抑制作用,但抑制作用不是很大且這種影響相比水稻來說要小一點(diǎn),這說明Al毒對苜蓿的影響不太強(qiáng),表明苜蓿是一種耐鋁性較強(qiáng)的植物。因此,應(yīng)該大力推廣這種植物作為動(dòng)物的飼料。由圖4可知,12個(gè)苜蓿品種除了阿爾岡金和巨人201之外,其他品種的根相對伸長率都在60%以上,其中金皇后、WL-323HQ、馴虎、WL-232HQ、胖多、獵人河這幾個(gè)品種根相對伸長率都在80%以上,受Al毒的影響比較小,而阿爾岡金和巨人201受Al毒的影響比較大都在50%以下,這2個(gè)品種在酸性土壤中則根部含有的Al含量較多,對動(dòng)物來說食用這種植物對它們的身體是有害的,另外通過食物鏈的作用對人類也是有一定影響的。因此,應(yīng)該大力發(fā)展金皇后、WL-323HQ、馴虎、WL-232HQ、胖多、獵人河等這些耐鋁的苜蓿品種。通過分析可知,苜蓿各品種的根相對伸長率存在顯著性差異。
苜蓿的株高受Al毒的影響相比根受Al毒的影響來說還要小,一方面說明苜蓿是耐鋁的,另一方面更說明Al毒對植物的作用主要體現(xiàn)在植物的根部,而不是地上部分,對地上部分的影響很小。由圖5可知,所有苜蓿品種的株高相對伸長率都在70%以上,WL-323HQ、皇冠和獵人河3個(gè)品種株高相對伸長率都接近100%,說明Al對這3個(gè)品種的株高幾乎沒有影響,此外金皇后的株高相對伸長率也在90%以上,受Al的影響也十分小。結(jié)合圖4和圖5可知,金皇后、WL-323HQ和獵人河這3個(gè)品種最優(yōu),一是因?yàn)樗鼈兊南鄬Ω扉L率都比較高,受鋁的影響小;二是因?yàn)樗鼈兊闹旮呦鄬ι扉L率也都在90%以上,受鋁的影響十分小,再加上因?yàn)橹旮叩拇笮【褪沁@種植物提供給動(dòng)物食物的多少,結(jié)合這幾點(diǎn)可認(rèn)為金皇后、WL-323HQ和獵人河這3個(gè)品種是最優(yōu)的,是耐鋁性品種。另外,通過對相對Al含量的測定來進(jìn)一步確認(rèn)耐鋁性苜蓿品種。通過分析可知,苜蓿各品種的株高相對伸長率存在顯著性差異。
苜蓿的株高受Al毒的影響相比根受Al毒的影響來說還要小,一方面說明苜蓿是耐鋁的,另一方面說明Al毒對植物的作用主要體現(xiàn)在植物根部,而不是地上部分,對地上部分的影響很小(圖5)。
由圖6可知,金皇后的相對Al含量最小,接近100%,也就是說對照和試驗(yàn)的金皇后根尖吸收的鋁含量相差不多,WL-323HQ次之,相對Al含量最大的是巨人201,為330%左右,是金皇后的3倍多、WL-323HQ的2倍多。大部分品種的相對Al含量范圍在150%左右。由圖4、圖5、圖6可知,最耐鋁的苜蓿品種是金皇后和WL-323HQ,敏感性品種是巨人201。通過分析可知,苜蓿各品種的相對Al含量存在顯著性差異。
2.3 對大豆幼苗根長和株高及根尖相對鋁含量的影響
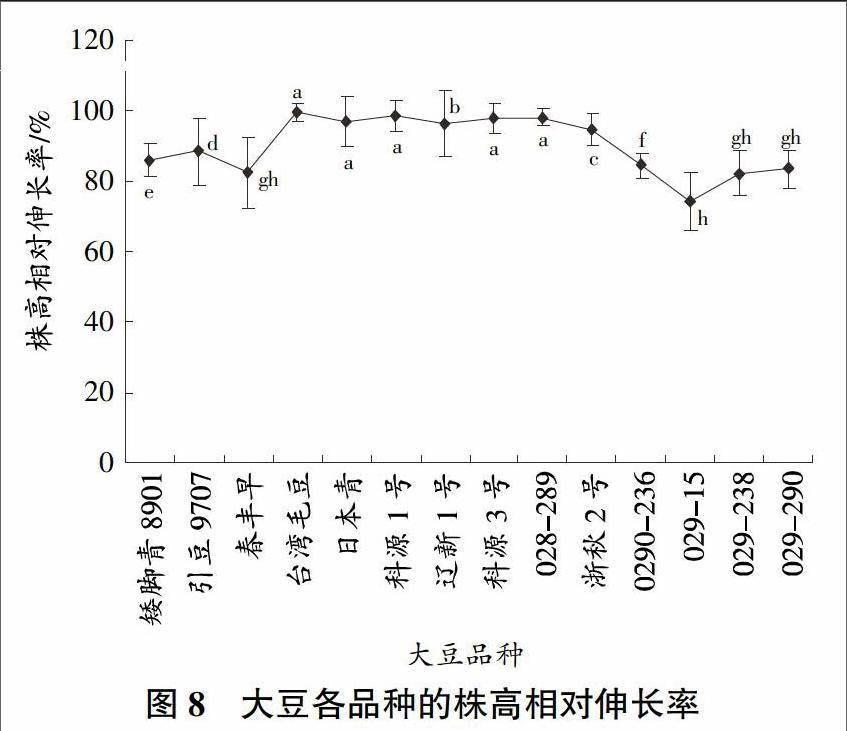
本試驗(yàn)結(jié)果表明,經(jīng)過鋁處理的大豆的根長要明顯矮于未經(jīng)過鋁處理的大豆,而且經(jīng)過鋁處理后的大豆的根部都呈現(xiàn)一定的紅色,可見鋁毒能夠抑制大豆根部的生長,影響大豆植株對營養(yǎng)成分的吸收。大豆的根相對伸長率基本分布在70%以下,這說明鋁毒對大豆根部的影響還是比較大的,根部受影響最大的品種是遼新1號,根相對伸長率在40%以下,相對影響最小的品種是029-236,它的根相對伸長率在75%左右,大約是遼新1號根相對伸長率的2倍,其次是029-15、029-238、029-290和矮腳青,它們的根相對伸長率也都在65%左右。因此,在這些大豆品種中根部比較耐鋁的品種是029-236、029-15、029-238、029-290和矮腳青(圖7)。大豆根系迅速吸收和積累A1但地上部分鋁累積并沒有明顯的變化,受鋁毒的影響和地上部分相差很大,它們的相對株高伸長率基本上都在80%以上(圖8),其中臺灣毛豆、科源1號、科源3號和029-289的株高相對伸長率都接近100%,幾乎不受任何影響,日本青和遼新1號也都在90%以上,影響最大的是春豐早、029-15、029-238和029-290,也都在75%以上。大豆株高相對伸長率都比較接近,相差不多,差距最大的是臺灣毛豆和029-15,相差25%以上,株高相對伸長率相對根長的伸長率的波動(dòng)要小得多,可見鋁毒對株高的影響相比根要小得多。大豆的根部是吸收鋁毒的主要部位。從植物的株高來看,臺灣毛豆、科源1號、科源3號和029-289這4個(gè)品種最好,日本青和遼新1號2個(gè)品種次之,株高相對伸長率較好的是029-15、029-238和029-290。在綜合考慮中要把大豆根相對伸長率因素放在第1位。通過分析,大豆各品種的根相對伸長率和株高相對伸長率存在顯著性差異。
大豆品種受鋁毒的影響和水稻、苜蓿基本相同,都是根部的鋁含量明顯大于地上部分。根部是植物吸收營養(yǎng)最主要的部位,植物根部的鋁含量多少對植物的影響很大。在篩選的大豆品種中,相對Al含量最少的品種是遼新1號,僅為130%左右,較大的品種是引豆9701和春豐早,分別為500%和400%以上,分別是遼新1號的4倍、3倍左右。除遼新1號外,029-289、浙秋2號和029-15也都在250%以下,029-238和029-290較大,它們的相對Al含量在350%左右,相對Al含量越大,說明根部鋁含量越大,越不耐鋁,反之則越耐鋁。引豆9701和春豐早的根部相對Al含量最大,029-289、浙秋2號、029-15和029-290較小,遼新1號最小(圖9)。綜上考慮,結(jié)合圖7和圖8可知,029-15是耐鋁品種,是試驗(yàn)最終所要的品種。敏感性品種是春豐早。通過Spass分析,大豆各品種的相對Al含量存在顯著性差異。
3 結(jié)論與討論
本試驗(yàn)結(jié)果表明,在50 μmol/L的鋁溶液處理下,鋁毒被植物根部吸收明顯大于地上部分的吸收,根據(jù)各個(gè)品種對鋁毒的吸收情況,來篩選出耐鋁性好的品種,對提高它們的質(zhì)量起到了非常重要的參考價(jià)值。鋁離子是酸性土壤上作物生長最主要的限制因子[10]。不同的作物、同一作物的不同品種間,其耐鋁性可能存在差異。在玉米、大豆、水稻、大麥、小麥、苜蓿、馬鈴薯、番茄等作物中都觀察到耐鋁性的基因型差異[11-12],在本試驗(yàn)中也發(fā)現(xiàn),不同品種的水稻、苜蓿和大豆對鋁的耐性具有基因型差異[13-14]。肉眼可見的鋁毒癥狀包括根伸長生長的抑制[15]、根尖膨大[16]、表皮脫落[17]等。由于根尖比成熟的根部組織積累更多的鋁,因此根尖被認(rèn)為是鋁毒的最初作用部位[18-19]。在含鋁營養(yǎng)液中。種子根凈增長度和種子根相對伸長長率可用于玉米耐鋁性的早期鑒定[20-11]。蘇木精染色廣泛用于細(xì)胞遺傳研究,也可用少水稻、苜蓿和大豆早期無損傷性的耐鋁性鑒定[21-22]。本文研究結(jié)果顯示,所用篩選材料對鋁脅迫的反應(yīng)具有基因型差異。耐鋁基因型具有植株相對生物量較高、根系生長受抑制程度相對較低和的蘇木精著色程度較低等共同特點(diǎn)。
根部性狀作為耐鋁性的評價(jià)指標(biāo)為大量研究所采用。測定溶液培養(yǎng)中根長(主根)對Al3+的反應(yīng)已成為植物耐鋁性鑒定的通用方法。Magnavaca研究認(rèn)為,在營養(yǎng)液培養(yǎng)中,采用根長法評價(jià)玉米自交系耐鋁性的最佳鋁濃度為6 mg/L,目前已發(fā)展成為熱帶玉米種質(zhì)耐鋁性篩選的有效方法,且與其他篩選技術(shù)具有很好的相關(guān)性[11]。在本試驗(yàn)中0.05 mmol/L Al3+為適宜的鋁處理濃度。在這種脅迫條件下,種子根凈增長度(NSRL)和相對伸長率(RSRL)變幅較大。本文將水稻、苜蓿和大豆種子根相對伸長率(RSRL)定義為鋁脅迫與對照的種子根凈增長之比,由于考慮了各品種根系生長的自身特性,因此較種子根凈增長度(RSRL)更適宜于耐鋁性的評價(jià)。在本研究中RSRL與植株地上部及整株相對生物量之間相關(guān)顯著,各品種的耐鋁性評價(jià)結(jié)果與以前土培試驗(yàn)中的耐酸性篩選結(jié)果[13]基木一致。而RSRL與植株相對生物量相關(guān)不顯著,這與Magnavaca等的研究結(jié)果一致,他們的分析表明,在鋁(0.185 mmol/L,Al3+)處理?xiàng)l件下,最初(鋁處理前)種子根長(RSRL)與NSRL存在極顯著的相關(guān)關(guān)系,但與RSRL相關(guān)不顯著[19]。因此,測定RSRL可作為一種快速簡便、準(zhǔn)確可靠的耐鋁性鑒定方法。關(guān)于高等植物鋁毒的機(jī)理目前知之甚少,許多方面尚停留在假說階段[23-24]。一般認(rèn)為,鋁毒癥狀是由于Al3+與根尖的相互作用導(dǎo)致根的發(fā)育被抑制所引起的。Morinmla等[19]用1 mol/L AlCl3處理洋蔥根尖,2 h后即可觀察到細(xì)胞分裂受阻,8 h后細(xì)胞分裂完全停止。本研究結(jié)果顯示,鋁處理2 d后,根尖明顯膨大,根系表皮脫落,而耐鋁基因型根系受抑制程度則明顯較輕。李德華、賀立源等研究發(fā)現(xiàn),耐鋁品種在鋁處理3 d后的根尖有膠狀物質(zhì),估計(jì)這是植物耐鋁的機(jī)制之一。也許附著在植物根尖上膠狀物質(zhì)可與上壤中的鋁結(jié)合,減少根尖對鋁的接觸機(jī)會(huì)[25]。研究發(fā)現(xiàn),有膠狀物質(zhì)附著的根經(jīng)過鋁處理后分別測定保留和除去膠狀物質(zhì)的根尖含鋁量,結(jié)果前者大于后者。這說明根系分泌的膠狀物質(zhì)可以和鋁結(jié)合[26]。
根際環(huán)境中鋁的生態(tài)化學(xué)行為、根系分泌物和微生物的生態(tài)毒理效應(yīng)受多種因素影響,且這此因素之間又相互影響、相互制約。本試驗(yàn)沒有對這些進(jìn)行深入研究。隨著學(xué)科的發(fā)展以及研究技術(shù)手段的進(jìn)步,可望從宏觀向微觀乃至分子、基因水平在以下幾方面開展深入研究:一是完善根系分泌物的收集、分離和鑒定技術(shù),建立根系分泌物原位收集方法。二是運(yùn)用生物工程技術(shù),揭開鋁脅迫對植物根細(xì)胞膜酶活性的變化規(guī)律,耐鋁酶活性與根系分泌物種類和濃度之間的關(guān)系,根尖釋放的分泌物與根尖細(xì)胞膜選擇透性之間的關(guān)系,酶在根系分泌物與鋁絡(luò)合反應(yīng)中的催化機(jī)理等。三是運(yùn)用基因工程技術(shù),加強(qiáng)植物抗鋁基因反應(yīng)機(jī)制以及植物遭受鋁毒時(shí)基因組變化的研究,積極運(yùn)用抗鋁基因的定位克隆和轉(zhuǎn)基因等技術(shù)選育抗鋁性強(qiáng)的植物新品種。
4 參考文獻(xiàn)
[1] WANGATSUMA T,YAMASAKN K.Relationship between differential aluminum tolerance and plant induced pH change of medium among barley cultivars[J].Soil Science Plant Nutrition,1987,31(4):521-535.
[2] 楊志敏,汪瑾.植物耐鋁的生物化學(xué)與分子機(jī)理[J].植物生理與分子生物學(xué)學(xué)報(bào),2003,29(5):361-366.
[3] 肖鳳娟,張欣杰.鋁、鎂、鋅、錳等金屬離子與鈣調(diào)素相互作用研究進(jìn)展[J].河北省科學(xué)院學(xué)報(bào),2003,20(2):163-167.
[4] 應(yīng)小鵬,劉鵬,徐根娣.土壤中的鋁及其植物效應(yīng)的研究進(jìn)展[J].生態(tài)環(huán)境,2003,12(2):237-239.
[5] LARSEN P B,STENZLER L M,TAI CY,et a1.Molecular and physiological analysis of Ambidopsis uzutnuts exhibiting altered sensitivities to aluminum[J].Plant and Soil,1997,192:3-7.
[6] LAZOF D B,GOLDSM IN JG,RUFTH T W.Rapid up-take of aluminum into cells of intact soybean root tips Amicroanalytical study using sceondary in mass species-toxicity[J].Plant Physiology,1994,106(3):1107-1114.
[7] 孔繁翔,桑偉蓮,蔣新,等.鋁對植物毒害及植物抗鋁作用機(jī)理[J].生態(tài)學(xué)報(bào),2000,20(5):855-861.
[8] 赫魯寧,劉濘田.鋁對水稻幼苗的生理影響[J].植物學(xué)報(bào),1989,27(11):847-853.
[9] 秦瑞君,陳福興.湘南紅壤作物苗期鋁中毒的研究[J].植物營養(yǎng)與肥料學(xué)報(bào),1999,5(1):50-55.
[10] MA J F,RYAN P R,DELHAIZE E.Aluminum tolerance in plants and the complexing role of organic acids[J].Trends in Plant Science,2001,6:273-278.
[11] CANCADO G M A,LOGUERCIO L L,MARTINS P R,et al.Hematoxylin staining as a phenotypic index for aluminum tolerance selection in tropical maize(Zea mays L.)[J].Theor Appl Genet,1999,99:747-754.
[12] 黃邦全,自景華,薛小橋.植物鋁毒害及遺傳育種研究進(jìn)展[J].植物學(xué)通報(bào),2001(18):385-395.
[13] 賀立源,徐尚忠,李建生.玉米自交系苗期耐酸的生物學(xué)和營養(yǎng)學(xué)特性[J].作物學(xué)報(bào),2000(26):205-209.
[14] 李德華,賀立源,劉武定.耐鋁的和對鋁敏感的玉米自交系根系的有機(jī)酸分泌[J].植物生理與分子生物學(xué)學(xué)報(bào),2003,29(2):114-120.
[15] DELHAIZ E,ROAN P R.Aluminum oxicity and tolerance in plants[J].Plant Physiol,1995,107:315-321.
[16] JONES D L,KOCHIAN L V.Aluminum inhibition of 1,4,5-trisphosphate signal transduction pathway in wheat roots:a role in aluminum[J].Toxicity Plant Cell,1995:1913-1922.
[17] LARSEN P B,DEGENHARDT J,TAI C Y,et al.ArabodopsisMut-ants with increased aluninum resistance exhibit altered patterns of aluminumaccumulation and organic:acid release from roots[J].Plant Phaysiol,1998,117:9-17.
[18] DELHAIZE E,RYAN P R.Aluminum toxicity and tolerance in plants[J].Plant,1995,107:315-321.
[19] MORINMLA S,TAKAHASHI E,MATSUMOTO H.Association of alum-inum with nuclei and inhibition of cell division in onion(Allium cepa)roots[J].Z. Pflarcenphysiol,1978,88:395-401.
[20] MAGNAVACA R.Genetics Aspects of Plant Mineral Nutrition[J].Martinus Nitrition off Publishers,1987,88:201-212.
[21] RYAN P R,DITOMASO J M,KOCHIAN L V.Aluminum toxicity in roots:an investigation of spatial sensitivity and role of the cap[J].J Exy Bot,1993,44:437-446.
[22] JORGE R A,ARRUDA P.Aluminum induced organic acids exudation by roots of an aluminum-tolerant tropical maize[J].Phytochenaistry,1997,45:675-681.
[23] TAYLOR G J,MCDONALD-STEPHENS J L,HUNTER D B.Direct Measure ment of aluminum uptake and distribution in single cell of Chara corallina[J].Plant Plysiol,2000,123:987-996.
[24] YAMAMOTO Y,KOHAYASHI Y,MATSUMOTO H. Lipid peroxidation is an early symptom triggered by aluminum,but not the primary cause of elongation inhibition in pea roots[J].Plant Physiol,2001,125:199-208.
[25] HENDERSON M,OWNBY J D.The role of root cap mucilage secration in aluminum tolerane in wheat[J].Curr Top Plant Biochem Plysiol,1991,10:134-141.
[26] 黎曉峰,馬建鋒,松木英明.玉米根冠粘膠和鋁的結(jié)合及有機(jī)酸累積[J].植物生理與分子生物學(xué)學(xué)報(bào),2002(28):121-126.
猜你喜歡
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
