鹽脅迫對鹽芥硝酸鹽的吸收及根系生長發(fā)育的影響
2017-10-17 05:31:15姜娜娜崔鳳劉譯陽韓燕張荃李軍李國衛(wèi)
山東農(nóng)業(yè)科學 2017年9期
關鍵詞:植物
姜娜娜, 崔鳳,劉譯陽,韓燕,張荃,李軍,李國衛(wèi)
(1.山東省農(nóng)業(yè)科學院生物技術研究中心/山東省作物遺傳改良與生態(tài)生理重點實驗室,山東 濟南 250100; 2.山東種業(yè)集團股份有限公司,山東 濟南 250100;3. 山東師范大學生命科學學院,山東 濟南 250014)
鹽脅迫對鹽芥硝酸鹽的吸收及根系生長發(fā)育的影響
姜娜娜1,2, 崔鳳1,劉譯陽1,韓燕1,張荃3,李軍1,李國衛(wèi)1
(1.山東省農(nóng)業(yè)科學院生物技術研究中心/山東省作物遺傳改良與生態(tài)生理重點實驗室,山東 濟南 250100; 2.山東種業(yè)集團股份有限公司,山東 濟南 250100;3. 山東師范大學生命科學學院,山東 濟南 250014)
氮素是植物生長發(fā)育所必需的主要營養(yǎng)元素,硝酸鹽是植物吸收氮的主要形式。鹽脅迫是一類非生物脅迫,抑制植物的生長發(fā)育。為了探討鹽脅迫影響硝酸鹽吸收與植物生長發(fā)育的關系,本研究以耐鹽模式植物鹽芥為試驗材料,測定了鹽脅迫下鹽芥體內(nèi)硝酸鹽含量的變化,分析其與根系發(fā)育的關系,并利用RNA-seq技術分析了鹽脅迫條件下鹽芥中硝酸根轉運蛋白(nitrate transporter,NRT)基因的表達調(diào)控。結果顯示,鹽脅迫后鹽芥葉片和根中的硝酸鹽含量均顯著下降,且根系干重與硝酸鹽含量呈正相關關系,表明鹽脅迫抑制了鹽芥對硝酸鹽的吸收,并且影響根系的發(fā)育。RNA-seq共檢測到10個鹽芥硝酸根轉運蛋白基因的表達,大部分基因的表達受鹽脅迫的調(diào)控,可能參與了鹽脅迫條件下鹽芥體內(nèi)硝酸鹽的吸收和轉運。綜合上述結果,推測鹽脅迫抑制鹽芥對硝酸鹽的吸收,可能是導致鹽脅迫下植物生長發(fā)育受抑制的因素之一。
鹽芥;鹽脅迫;硝酸鹽吸收;硝酸根轉運蛋白
AbstractNitrogen is a major nutrient element necessary to plant growth and development, and nitrate is the main form of nitrogen absorbed by plants. Salt stress is one type of abiotic stress, which defects plant growth and development. In order to study the effects of salt stress on nitrate absorption and plant growth and development, we employed salt-tolerant model plant Eutrema salsuginea as materials. The changes of nitrate concentrations inEutremasalsugineaunder salt stress were measured, and its relationship with root development was analyzed. In addition, the expression levels of nitrate transporter (NRT) genes were analyzed using RNA-seq technology under salt stress conditions. The results showed that the nitrate contents in leaves and roots were both significantly decreased after salt stress treatment, and the root dry weight had a positive correlation with nitrate content. It implied that salt stress inhibited plant nitrate absorption, and further affected the growth of root system. Totally, ten NRTs were detected by RNA-seq analysis, and the expression of most of NRTs was regulated by salt stress. It suggested that these genes might be related with nitrate absorption and transport inEutremasalsugineaunder high salt environment. In conclusion, we presume that salt stress inhibits the nitrate absorption inEutremasalsuginea, which might be one of the factors limiting the growth and development of plants under salt stress.
KeywordsEutremasalsuginea; Salt stress; Nitrate absorption; Nitrate transporter
氮素是植物需求量最大的營養(yǎng)元素,其吸收和利用是影響植物生長和發(fā)育的主要因素[1],而硝酸鹽是植物吸收利用的主要氮素形式。硝酸鹽不僅是植物生長必需的營養(yǎng)物質,也可以作為一種信號調(diào)節(jié)根冠比和根的形態(tài)建成[2-4]。 液泡內(nèi)的硝態(tài)氮是重要的滲透調(diào)節(jié)物質,用于調(diào)節(jié)植物的生長發(fā)育[5]。
鹽芥是分布在鹽堿地中的一種鹽生植物,耐鹽性極強[6]。它與模式植物擬南芥的親緣關系非常近,對從鹽芥中克隆的數(shù)百個EST (expressed sequence tag)序列的分析說明,鹽芥的cDNA和氨基酸序列與擬南芥的相似度為90%~95%[7]。從全基因組測序的結果來看,鹽芥93.7%的基因家族與擬南芥相似,有研究者提出把鹽芥作為研究植物耐鹽性的模式物種[8-11]。
硝酸根轉運蛋白(nitrate transporter, NRT)是一類膜蛋白,參與植物根系對硝酸根的吸收和體內(nèi)分布。根據(jù)其對硝酸根親和力的差異,可以分為低親和力硝酸根轉運蛋白(low-affinity transport system, LATS)和高親和力硝酸根轉運蛋白(high-affinity transport system, HATS),大部分NRT1為低親和力硝酸根轉運蛋白,NRT2為高親和力硝酸根轉運蛋白[12]。其中,擬南芥中的AtNRT1.1(CHL1)根據(jù)環(huán)境中硝酸根濃度差異可以表現(xiàn)出對硝酸根的雙重親和性,當環(huán)境中硝酸根濃度較高時,AtNRT1.1表現(xiàn)為對硝酸根的低親和性;而當環(huán)境中的硝酸根濃度較低時,AtNRT1.1蛋白序列中的101位蘇氨酸發(fā)生磷酸化后,表現(xiàn)為對硝酸根的高親和性[13]。近年來,人們對鹽芥的耐鹽機理已經(jīng)有了較為深入的認識[8]。然而,鹽脅迫對鹽芥硝酸根吸收的影響以及鹽芥對硝酸根吸收與植物耐鹽性的關系尚未見相關報道。
本研究通過測定不同濃度鹽處理對鹽芥生長以及葉片和根系內(nèi)硝酸鹽含量的影響,明確了鹽脅迫條件下硝酸鹽吸收的減少可能是影響植物發(fā)育的重要因素之一,為植物耐鹽方面的研究提供新的思路,為植物耐鹽基因工程改造提供理論基礎。
1 材料與方法
1.1植物培養(yǎng)及鹽處理
鹽芥(山東生態(tài)型)由本實驗室保存。將鹽芥種子播種于1/2MS培養(yǎng)基上萌發(fā),17 d后轉移到盛有Hoagland 營養(yǎng)液的水培盒中進行水培。每個水培盒放6棵苗,水培18 d后,選生長一致的幼苗進行鹽處理。鹽處理濃度梯度為:0、50、100、150、200、300、400 mmol/L的NaCl,鹽處理時間為15 d。每個濃度處理6棵苗,每個處理重復3次。
1.2植株地上部和根干重測定
鹽芥鹽處理15 d時,將植株完整地從水培盒中取出,用蒸餾水將表面沖洗干凈,吸水紙吸干表面水分,將地上部分和根分開,用錫箔紙分別包好,置于80℃烘箱內(nèi)烘干至恒重,分別稱取每株地上部和根的干重。
1.3葉片和根系硝酸鹽含量的測定
采用還原法測定硝酸鹽含量。取0、0.1、0.2、0.3、0.5、1.0 mmol/L硝酸鉀溶液各10 μL作為標準樣品,加入90 μL硝酸還原酶,室溫反應30 min。然后加入60 μL N-1奈乙二胺鹽酸和對氨基苯磺酰胺(N-1奈乙二胺鹽酸∶對氨基苯磺酰胺=1∶1),2 h后,540 nm測定吸光值,制作標準曲線。
在每份烘干的葉片和根樣品中加入適量0.1 mol/L的HCl,過夜,溶解硝酸鹽。溶解液用水稀釋100倍,然后取10 μL稀釋液作為待測樣品,按照標準品的測定方法測定吸光值。根據(jù)標準曲線方程計算樣品的硝酸鹽濃度,最后再根據(jù)稀釋倍數(shù)和樣品重量計算硝酸鹽含量。
1.4鹽芥表達譜分析
鹽芥RNA提取、文庫構建、高通量測序參照Wang等[14]已發(fā)表論文。簡述如下,將培養(yǎng)18 d的鹽芥幼苗用200 mmol/L NaCl處理 48 h,處理后根和葉片分別取樣,利用RNAiso Reagent試劑盒提取RNA,并利用DNase Ⅰ去除基因組DNA污染。RNA質量檢測、建庫等步驟詳見文獻[14]。RNA-seq由北京基因組研究所(BGI)的Illumina HiSeq 2000 平臺完成,每組樣品設置兩次生物學重復。
2 結果與分析
2.1鹽脅迫對鹽芥生長的影響
試驗結果表明,水培培養(yǎng)液中含50~400 mmol/L NaCl條件下,鹽芥均可以完成其生活史。但鹽脅迫下,鹽芥的生長發(fā)育受到抑制,且鹽濃度越高受到的抑制越明顯。與對照相比,處理濃度為300、400 mmol/L的植株葉片出現(xiàn)黃化干枯現(xiàn)象(圖1)。
鹽脅迫下,鹽芥地上部和根干重的測定結果見圖2,結果顯示,NaCl濃度大于100 mmol/L時,鹽芥地上部分和根系干重均有不同程度下降。

圖1不同濃度鹽處理15 d鹽芥生長情況
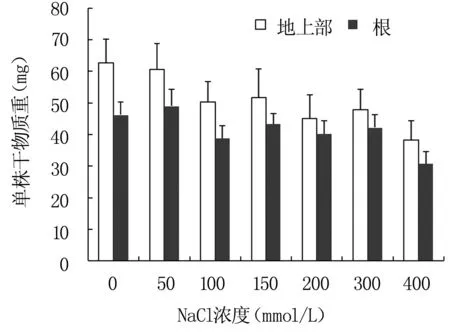
圖2不同濃度NaCl處理鹽芥地上部和根干重變化
2.2鹽脅迫對鹽芥葉片硝酸鹽含量的影響
鹽處理后,鹽芥葉片內(nèi)硝酸鹽含量均比對照明顯降低。在50~400 mmol/L NaCl各供試濃度下,鹽芥葉片硝酸鹽含量分別比對照降低53.4%、72.9%、83.9%、93.4%、100%和99.6%(圖3)。

圖3鹽芥在不同濃度鹽脅迫下葉片中硝酸鹽含量比較
2.3鹽脅迫對鹽芥根硝酸鹽含量的影響
鹽處理后,鹽芥根系的硝酸鹽含量也發(fā)生了明顯改變,各處理下的硝酸鹽含量均低于對照,其中300、400 mmol/L處理的下降最明顯。在50~400 mmol/L NaCl各處理濃度下,根系中硝酸鹽含量分別比對照降低49.3%、55.1%、53.5%、78.1%、99.4%和97.1%(圖4)。

圖4鹽芥在不同濃度鹽脅迫下根中硝酸鹽含量比較
2.4根干重與硝酸鹽含量的關系
比較鹽處理后植株根系硝酸鹽的總量與根干重發(fā)現(xiàn),根系中硝酸鹽的含量與根干重呈正相關關系(R2=0.62),即根系中的硝酸鹽含量越高,根干重越大(圖5)。這一結果說明鹽脅迫條件下根系的生長發(fā)育與硝酸鹽的吸收密切相關,硝酸鹽的含量降低可能是鹽脅迫抑制鹽芥根系發(fā)育的因素之一。

注:使用Pearson線性回歸分析。
圖5根系干重與硝酸鹽含量的關系
2.5鹽脅迫對硝酸根轉運蛋白的表達調(diào)控
鹽脅迫條件下,鹽芥根系和葉片硝酸鹽積累均顯著下降。因為鹽脅迫條件下植株生長受到抑制,硝酸鹽的利用減少,因此推測根系吸收硝酸鹽的能力下降。為此,我們利用表達譜分析了鹽芥硝酸根轉運蛋白(Nitrate Transporter,NRT)基因對200 mmol/L NaCl脅迫的響應。通過對表達譜數(shù)據(jù)的分析,發(fā)現(xiàn)10個表達豐度較高的NRT基因,聚類分析結果表明這些基因可以分為兩組:4個NRT1s和6個NRT2s,分別屬于低親和力和高親和力硝酸根轉運蛋白基因(圖6)。

注:鹽芥NRT基因序列信息來源于鹽芥基因組Eutrema salsugineum v1.0 (https://phytozome.jgi.doe.gov/pz/portal.html)。利用Clustal Omega 在線軟件(https://www.ebi.ac.uk/Tools/msa/clust-
alo/)對NRT蛋白序列進行多序列比對。
圖6鹽芥鹽脅迫響應NRT進化樹分析
在葉片中,鹽處理后只有Thhalv10023363m(NRT1)表達上升比較明顯,上升3.46倍。Thhalv10007157m(NRT1)和Thhalv10007338m(NRT2)的表達被抑制,分別只有正常生長條件下的35%和21%。Thhalv10007254m(NRT1)和Thhalv10013237m(NRT2)為鹽芥根特異表達基因,其表達均受鹽脅迫抑制,分別為正常條件下的20%和32%。Thhalv10007157m(NRT1)在根中的表達量僅為正常條件下的29%(圖7)。
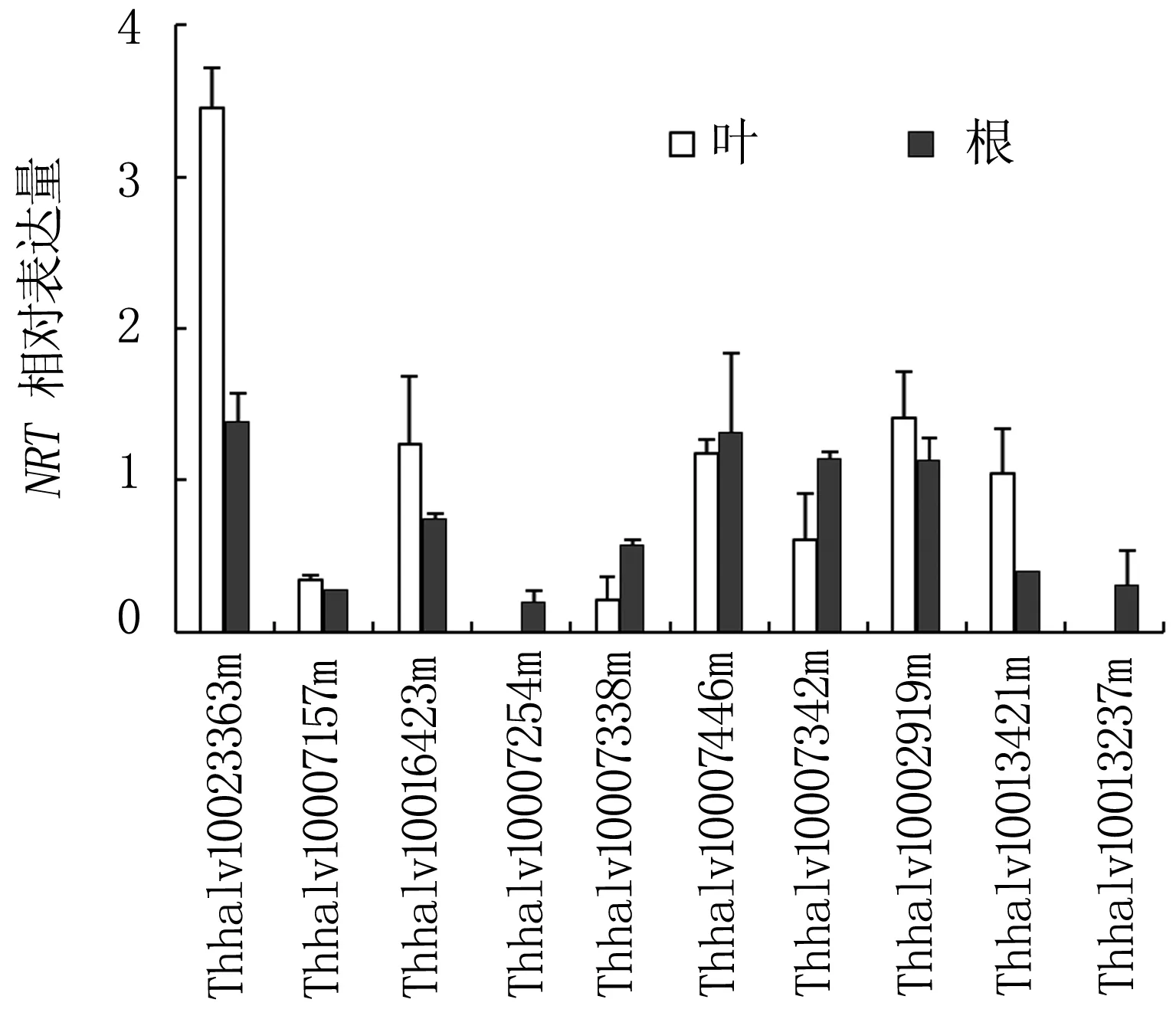
注:NRT相對表達量(NRT relative expression)為鹽脅迫條件下
3 討論與結論
硝酸根轉運蛋白(NRT)家族龐大,參與植物根系對硝酸鹽的吸收、轉運、分布和存儲過程,與植物生長發(fā)育等重要生理活動相關聯(lián)。除了參與硝酸根的轉運,擬南芥中的NRT1.1還參與了生長素等的轉運,與側根的發(fā)育等生命活動密切相關[15]。NRT1.8屬于NRT1亞族,參與硝酸根從木質部的轉運,還參與了擬南芥對重金屬鎘脅迫的響應,可以提高植物對鎘脅迫的抗性[16]。
本研究結果顯示,鹽脅迫使鹽芥的生長發(fā)育受到了抑制,并且鹽濃度越高抑制效果越明顯。在300 mmol/L和400 mmol/L NaCl處理15 d時,鹽芥出現(xiàn)了明顯的葉片干枯現(xiàn)象。鹽芥葉片和根系中硝酸鹽含量的測定結果顯示,葉片和根系中的硝酸鹽含量均下降,并且葉片下降的更為明顯,在300 mmol/L和400 mmol/L時,硝酸鹽含量接近于零。這與鹽脅迫下鹽芥生長發(fā)育受到抑制的情況相吻合,根系干重與其硝酸鹽含量呈線性正相關,推測鹽脅迫抑制根系對硝酸鹽的吸收和積累可能是鹽芥生長發(fā)育受到抑制的因素之一。
盡管鹽芥為鹽生植物,與正常Hoagland溶液相比,高于50 mmol/L NaCl仍然會抑制其生長發(fā)育。硝酸鹽在植物中的積累降低,利用也減少(因為生長受到抑制),說明鹽脅迫條件下根系對硝酸鹽的吸收能力下降。氮元素是構成蛋白質、核酸等生命大分子的重要成分,硝酸鹽的吸收利用與植物的生長發(fā)育密切相關。擬南芥地上部的生物量隨著葉片中硝酸鹽含量的增加而增大,硝酸鹽轉運蛋白突變體nrt2.1根系吸收硝酸鹽能力下降,同時植株的發(fā)育也延遲。此外,硝酸鹽吸收受到限制會降低植物對水分吸收的能力[17,18]。植物自身可以通過調(diào)控硝酸根轉運蛋白的表達調(diào)節(jié)根系對硝酸鹽的吸收能力[13]。硝酸根轉運蛋白基因是植物硝酸鹽吸收和轉運的關鍵基因,本研究表達譜數(shù)據(jù)顯示鹽芥中4個NRT1s和6個NRT2s表達豐度比較高。其中多個基因的表達明顯受到鹽脅迫的抑制,與鹽脅迫條件下鹽芥葉片和根系硝酸鹽含量下降的趨勢一致。推測這些基因表達豐度的下降可能是硝酸根積累下降的原因,Thhalv10023363m(NRT1)等基因表達上升或者維持不變,可能是鹽脅迫條件下鹽芥硝酸鹽吸收和轉運的關鍵基因。今后我們將進一步深入研究鹽芥NRT基因參與的硝酸鹽吸收調(diào)控的分子機理及其在提高植物耐鹽性中的作用。
[1] 李靜,張冰玉,蘇曉華,等.植物中的銨根及硝酸根轉運蛋白研究進展[J].南京林業(yè)大學學報,2012,36(4):133-139.
[2] Scheible W R, Gonzalez-Fontes A, Lauerer M, et al.Nitrate acts as a signal to induce organic acid metabolism and repress starch metabolism in tobacco [J].Plant Cell,1997,9(5):783-798.
[3] Zhang H, Forde B G.Regulation ofArabidopsisroot development by nitrate availability [J].J. Exp. Bot.,2000,51(342):51-59.
[4] Orsel M, Chopin F, Leleu O, et al.Characterization of a two component high-affinity nitrate uptake system inArabidopsisphysiology and protein-protein interaction [J]. Plant Physiol., 2006, 142:1304-1317.
[5] 王宇通,邵新慶,黃欣穎,等.植物根系氮吸收過程的研究進展[J].草業(yè)科學,2010,27(7):105-111.
[6] 趙昕,趙敏桂,譚會娟,等.NaCl脅迫對鹽芥和擬南芥K+、Na+吸收的影響[J].草業(yè)學報,2007,16(4):21-24.
[7] 劉愛榮,趙可夫.鹽脅迫下鹽芥滲透調(diào)節(jié)物質的積累及其滲透調(diào)節(jié)作用[J].植物生理與分子生物學學報,2005,31(4):389-395.
[8] Bressan R A, Zhang C Q, Zhang H, et al.Learning from theArabidopsisexperience, the next gene search paradigm[J].Plant Physiol.,2001,127:1354-1360.
[9] Zhu J K.Genetic analysis of plant salt tolerance usingArabidopsis[J].Plant Physiol., 2000, 124:941-948.
[10] Zhu J K. Plant salt tolerance [J]. Trends Plant Sci., 2001, 62:66-71.
[11] Volkov V, Wang B, Doming P J, et al.Thellungiellahalophile, a salt relative ofArabidopsisthaliana, possesses effective mechanisms to discriminate between potassium and sodium [J]. Plant Cell Environ., 2003, 27:1-14.
[12] Jose A O, Vega A, Bouguyon E, et al. Nitrate transport, sensing, and responses in plants [J]. Mol. Plant, 2016, 9(6): 837-856.
[13] Ho C H, Lin S H, Hu H C, et al. CHL1 functions as a nitrate sensor in plants [J]. Cell, 2009, 138(6): 1184-1194.
[14] Wang J, Zhang Q, Cui F, et al. Genome-wide analysis of gene expression provides new insights into cold responses inThellungiellasalsuginea[J]. Front Plant Sci., 2017, 8:713.
[15] Noguero M, Lacombe B. Transporters involved in root nitrate uptake and sensing byArabidopsis[J]. Front Plant Sci.,2016,7:1391.
[16] Li J Y, Fu Y L, Pike S M, et al. TheArabidopsisnitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance [J]. Plant Cell, 2010, 22(5):1633-1646.
[17] Remans T, Nacry P, Pervent M, et al. A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation inArabidopsis[J]. Plant Physiol., 2016, 140:909-921.
[18] Li G, Tillard P, Gojon A, et al. Dual regulation of root hydraulic conductivity and plasma membrane aquaporins by plant nitrate accumulation and high-affinity nitrate transporter NRT2.1[J]. Plant Cell Physiol., 2016, 57:733-742.
EffectsofSaltStressonAbsorptionofNitrateandRootGrowthandDevelopmentinEutremasalsuginea
Jiang Nana1,2, Cui Feng1, Liu Yiyang1, Han Yan1, Zhang Quan3, Li Jun1, Li Guowei1
(1.Bio-TechResearchCenter,ShandongAcademyofAgriculturalSciences/ShandongProvincialKeyLaboratoryofCropGeneticImprovement,EcologyandPhysiology,Jinan250100,China; 2.ShandongSeedGroupCo.,Ltd.,Jinan250100,China; 3.CollegeofLifeSciences,ShandongNormalUniversity,Jinan250014,China)
10.14083/j.issn.1001-4942.2017.09.006
2017-06-05
山東省現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術體系棉花創(chuàng)新團隊項目(SDAIT-03-03,SDAIT-03-05);山東農(nóng)業(yè)大學鹽堿地改良項目;國家自然科學基金項目(31601253);山東省農(nóng)業(yè)良種工程項目(2014LZ)
徐佳陵(1990—),男,碩士研究生,研究方向:棉花耐鹽分子機理。E-mail:1348573856@qq.com
宋憲亮(1972—),男,教授,博士,博士生導師,主要從事棉花遺傳育種研究。E-mail:songxl999@163.com 孫學振(1963—),男,教授,博士,博士生導師,主要從事棉花栽培生理研究。E-mail:sunxz@sdau.edu.cn
S580.1
A
1001-4942(2017)09-0037-05
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
