黃腐酸對(duì)Haematococcus pluvialis LUGU蝦青素積累和lcy基因表達(dá)量的影響
2017-09-26 07:31:48尚敏敏趙永騰徐軍偉余旭亞
食品與生物技術(shù)學(xué)報(bào) 2017年7期
尚敏敏, 趙永騰,趙 鵬, 李 濤,徐軍偉, 余旭亞
(昆明理工大學(xué) 生命科學(xué)與技術(shù)學(xué)院,云南 昆明 650500)
黃腐酸對(duì)Haematococcus pluvialis LUGU蝦青素積累和lcy基因表達(dá)量的影響
尚敏敏, 趙永騰,趙 鵬, 李 濤,徐軍偉, 余旭亞*
(昆明理工大學(xué) 生命科學(xué)與技術(shù)學(xué)院,云南 昆明 650500)
雨生紅球藻是蝦青素的主要來源,為探明黃腐酸 (FA)對(duì)雨生紅球藻Haematococcus pluvialis LUGU的影響,將不同質(zhì)量濃度的FA添加至對(duì)數(shù)生長期的藻液中置于脅迫條件下培養(yǎng),并分別對(duì)細(xì)胞生物量濃度、蝦青素積累質(zhì)量濃度以及番茄紅素β-環(huán)化酶(lcy)基因表達(dá)量進(jìn)行了測(cè)定。結(jié)果表明,黃腐酸質(zhì)量濃度為10 mg/L,藻細(xì)胞的生物量產(chǎn)率達(dá)到最大80.28 mg/(L· d),比對(duì)照組提高5.44%;而蝦青素最大質(zhì)量濃度為20.82 mg/L,是在FA質(zhì)量濃度為5 mg/L時(shí)測(cè)得,比對(duì)照組提高了86.89%。RT-PCR分析顯示,蝦青素合成的關(guān)鍵基因lcy的表達(dá)受FA的影響,當(dāng)添加5 mg/L和10 mg/L的黃腐酸時(shí)lcy基因最大的表達(dá)量分別為對(duì)照的1.2倍和0.7倍。實(shí)驗(yàn)證明FA誘導(dǎo)下的雨生紅球藻蝦青素的積累含量和lcy基因表達(dá)量呈正相關(guān)。
黃腐酸;誘導(dǎo);蝦青素;lcy基因;雨生紅球藻
雨生紅球藻是自然界中普遍存在的一種單細(xì)胞綠藻,屬于團(tuán)藻目,紅球藻科,紅球藻屬[1],在不利的環(huán)境條件下藻細(xì)胞逐漸變大并積累蝦青素[2],且其蝦青素的產(chǎn)量優(yōu)于其他已知的蝦青素來源[3-4]。雨生紅球藻細(xì)胞內(nèi)積累的蝦青素是生物體天然次級(jí)代謝產(chǎn)物和強(qiáng)抗氧化物質(zhì),已作為添加劑廣泛應(yīng)用于水產(chǎn)養(yǎng)殖和食品領(lǐng)域[5-7]。但目前蝦青素存在需求大,產(chǎn)量低、生產(chǎn)成本高等問題,因此,采取有效措施提高雨生紅球藻蝦青素的產(chǎn)量是解決這一問題的關(guān)鍵和研究熱點(diǎn)。
Steinbrenner等[8]將雨生紅球藻蝦青素合成途徑分為兩個(gè)階段:β-胡蘿卜素的形成為第一階段,蝦青素的合成為第二階段。番茄紅素是類胡蘿卜素中的一種,是類胡蘿卜素生物合成途徑中的重要的中間體,番茄紅素β-環(huán)化酶基因是在雨生紅球藻中蝦青素合成途徑中將番茄紅素環(huán)化生成β-胡蘿卜素的關(guān)鍵酶基因,β-胡蘿卜素進(jìn)而成為第二階段合成蝦青素的重要底物,lcy基因表達(dá)量的增加可以提高角黃質(zhì)轉(zhuǎn)化為蝦青素的效率。黃腐酸(Fulvic Acid,F(xiàn)A)是一類成分復(fù)雜的天然有機(jī)物質(zhì)[9]。陳玉玲等[10]在干旱條件下,冬小麥幼苗施加FA,發(fā)現(xiàn)其超氧物歧化酶SOD與過氧化物酶POD活性提高,植物抗干旱的能力增強(qiáng)。而FA作為誘導(dǎo)因子,對(duì)雨生紅球藻中蝦青素的積累影響和機(jī)理方面的研究還尚未見報(bào)道。
作者以雨生紅球藻 Haematococcus pluvialis LUGU為對(duì)象,研究FA對(duì)Haematococcus pluvialis LUGU生長和蝦青素積累的影響及FA脅迫對(duì)蝦青素合成關(guān)鍵酶基因lcy表達(dá)的作用,為優(yōu)化微藻培養(yǎng)條件、提高蝦青素的產(chǎn)量、探索FA誘導(dǎo)其合成機(jī)制提供理論基礎(chǔ)。
1 材料與方法
1.1 雨生紅球藻的培養(yǎng)
雨生紅球藻Haematococcus pluvialis LUGU為作者所在實(shí)驗(yàn)室篩選、保存。利用BG-11為基礎(chǔ)培養(yǎng)基,接種到3 L(內(nèi)置2 L培養(yǎng)基)光生物反應(yīng)器中,室內(nèi)恒溫(25±1)℃,光照強(qiáng)度2 800 lx,通入無菌空氣進(jìn)行高密度培養(yǎng),直至培養(yǎng)到生長對(duì)數(shù)期。
1.2 黃腐酸的配制
根據(jù)預(yù)試驗(yàn)結(jié)果,實(shí)驗(yàn)共設(shè)置0,5,10 mg/L 3組質(zhì)量濃度梯度的黃腐酸,每組設(shè)3個(gè)平行樣,將對(duì)數(shù)生長期的藻細(xì)胞收集后重新懸浮,以2×105個(gè)/ mL的接種量接入誘導(dǎo)培養(yǎng)基中。每隔一天取樣一次,測(cè)定H.pluvialis LUGU蝦青素質(zhì)量濃度。
1.3 蝦青素質(zhì)量濃度的測(cè)定
為了測(cè)定基礎(chǔ)培養(yǎng)基中培養(yǎng)的 H.pluvialis LUGU蝦青素的產(chǎn)量,采用Boussiba等[11]的方法稍加改進(jìn)測(cè)定其蝦青素含量。每隔一天定期取出5 mL處于誘導(dǎo)階段的藻液,5 000 r/min離心5 min,棄上清液。于收集的藻細(xì)胞沉淀中加入質(zhì)量分?jǐn)?shù)5% KOH和體積分?jǐn)?shù)30%甲醇混合液置于65°水浴鍋內(nèi)5 min以破壞葉綠素,5 000 r/min離心收集沉淀,加入3 mL二甲亞砜 (DMSO),利用超聲波破壁(20 s/5 s,輸出功率40 w),反復(fù)抽提直至藻體發(fā)白后,高速離心機(jī)10 000 r/min離心10 min,取上清液于490 nm下測(cè)定A值。按公式c(mg/L)=(4.5×A490× Va)/Vb計(jì)算蝦青素質(zhì)量濃度(A:吸光值,Va:DMSO的體積,Vb:藻液體積)。
1.4 雨生紅球藻lcy基因表達(dá)分析
本實(shí)驗(yàn)由Primer 5.0軟件設(shè)計(jì),上海生工生物工程技術(shù)服務(wù)有限公司合成lcy酶基因的上下游擴(kuò)增引物:5’-CTTCTTCTCCGCCTTCTTCA-3’與5’-GCATCCTACCGCTCAAAGAA-3’,擴(kuò)增長度為565bp。擴(kuò)增所得序列測(cè)序(上海生工)后BLAST比對(duì),并以此為模板設(shè)計(jì)熒光定量引物 lcyF(5’-GACTGGAGTGGGAAGAAC-3’) 與 lcyR (5’-CCTACCGCTCAAAGAAATA-3’),目標(biāo)產(chǎn)物為186bp。
Trizol法提取不同質(zhì)量濃度FA處理的雨生紅球藻RNA,利用逆轉(zhuǎn)錄試劑盒(TaKaRa)將RNA逆轉(zhuǎn)錄合成cDNA,以其為模板,以lcyF與lcyR分別為上下游引物進(jìn)行RT-PCR擴(kuò)增,檢測(cè)不同質(zhì)量濃度FA對(duì)雨生紅球藻lcy基因表達(dá)的影響。通過ABI 7500熒光定量儀對(duì)lcy基因的表達(dá)進(jìn)行定量,RT-PCR的數(shù)據(jù)結(jié)果用2-ΔΔCT的方法處理分析[12]。以18s(上游引物18sF:5’-CGGTCTGCCTCTGGTATG-3’,下游引物18sR:5’-GCTTGCTTTGAACACGCT-3’)基因作為內(nèi)標(biāo)以調(diào)節(jié)RNA的用量和循環(huán)數(shù),使內(nèi)標(biāo)基因在不同濃度誘導(dǎo)下的表達(dá)豐度一致。
進(jìn)行一元線性回歸分析(回歸方程中Y為蝦青素質(zhì)量濃度,x為lcy基因表達(dá)水平),研究不同質(zhì)量濃度FA誘導(dǎo)下雨生紅球藻蝦青素質(zhì)量濃度和lcy基因表達(dá)量之間的相關(guān)性。
2 結(jié)果與討論
2.1 FA對(duì)誘導(dǎo)培養(yǎng)微藻細(xì)胞生長的影響
圖1表明,不同質(zhì)量濃度的FA對(duì)誘導(dǎo)階段雨生紅球藻細(xì)胞生長的影響存在差異,微藻細(xì)胞的生長量隨著培養(yǎng)時(shí)間的延長而逐漸增加,培養(yǎng)至第3天后,添加FA的微藻細(xì)胞量都開始高于對(duì)照組。對(duì)照組和10 mg/L FA實(shí)驗(yàn)組在第7天生物量達(dá)到最大值;而FA質(zhì)量濃度為10 mg/L時(shí),生物量最大值提前出現(xiàn)在第5天。比較不同質(zhì)量濃度FA脅迫下H.pluvialis LUGU的最大生物量(圖2),結(jié)果顯示,添加10 mg/L FA,細(xì)胞生物量達(dá)到最大值0.562 g/L。可見在誘導(dǎo)藻細(xì)胞階段,適當(dāng)質(zhì)量濃度的FA能促進(jìn)藻細(xì)胞的生長,且當(dāng)黃腐酸的質(zhì)量濃度為10 mg/L時(shí),藻細(xì)胞的生物量產(chǎn)率達(dá)到最大80.28 mg/(L·d)。

圖1 不同質(zhì)量濃度黃腐酸對(duì)誘導(dǎo)階段雨生紅球藻LUGU藻細(xì)胞生長的影響Fig.1 Effect of different level of FA on the cell growth of H.pluvialis LUGU under induction period
2.2 FA對(duì)微藻總蝦青素產(chǎn)量的影響
圖3顯示,不同質(zhì)量濃度的FA對(duì)誘導(dǎo)階段H. pluvialis LUGU內(nèi)總蝦青素質(zhì)量濃度的影響存在差異,微藻總蝦青素的質(zhì)量濃度隨著培養(yǎng)時(shí)間的延長而逐漸增加。當(dāng)FA質(zhì)量濃度為0~10 mg/L時(shí),總蝦青素產(chǎn)量呈對(duì)數(shù)增長。比較不同質(zhì)量濃度FA脅迫下H.pluvialis LUGU總蝦青素最大產(chǎn)量的變化(圖4),結(jié)果發(fā)現(xiàn),在不添加FA的條件下,總蝦青素的產(chǎn)量為20.14 mg/g;當(dāng)添加5 mg/L的FA時(shí),總蝦青素的產(chǎn)量提高了1.86倍;當(dāng)FA質(zhì)量濃度的增加到10 mg/L時(shí),蝦青素的產(chǎn)量僅為對(duì)照組的1.09,可見10 mg/L質(zhì)量濃度的FA雖然促進(jìn)了藻細(xì)胞的生長但卻不利于蝦青素的合成,不同質(zhì)量濃度FA對(duì)蝦青素積累的影響存在顯著差異。
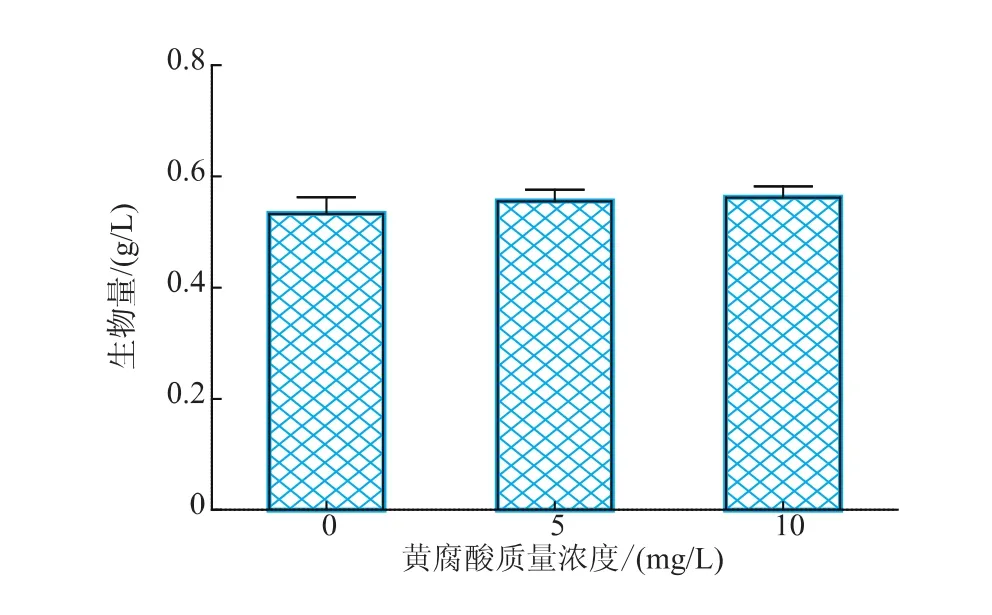
圖2 不同質(zhì)量濃度黃腐酸對(duì)雨生紅球藻的最大生物量的影響Fig.2 Effect of different level of FA on the maximum cell growth of H.pluvialis LUGU under induction period
2.3 FA誘導(dǎo)下微藻蝦青素產(chǎn)量和lcy基因表達(dá)量的關(guān)系
用RT-PCR的方法測(cè)定不同誘導(dǎo)時(shí)間內(nèi),不同濃度梯度FA誘導(dǎo)的H.pluvialis LUGU蝦青素相關(guān)合成基因lcy的表達(dá)水平(圖5),結(jié)合蝦青素產(chǎn)量變化趨勢(shì),F(xiàn)A質(zhì)量濃度為5 mg/L時(shí),lcy的表達(dá)量至第7天達(dá)到最大值,為對(duì)照組表達(dá)量的1.2倍,蝦青素產(chǎn)量至第7天達(dá)到最大值20.817 mg/g。FA質(zhì)量濃度為10 mg/L時(shí),蝦青素產(chǎn)量至第7天達(dá)到最大值12.231 mg/g,lcy基因的表達(dá)量亦于第7天呈現(xiàn)出最大值,為對(duì)照的0.7倍。從不同F(xiàn)A質(zhì)量濃度誘導(dǎo)H.pluvialis LUGU上來看,質(zhì)量濃度為5 mg/L時(shí),lcy基因表達(dá)量最大,這與該濃度下蝦青素產(chǎn)量最大的結(jié)果一致。
不同濃度FA誘導(dǎo)H.pluvialis LUGU蝦青素產(chǎn)量和lcy基因表達(dá)量之間的相關(guān)性表明,當(dāng)lcy表達(dá)水平低時(shí),微藻蝦青素產(chǎn)量隨其后表現(xiàn)出低產(chǎn)量;如lcy表達(dá)水平提高,蝦青素的產(chǎn)量隨之增加,兩者呈現(xiàn)線性相關(guān),回歸方程為:y=0.058 5x+0.004 7,R2=0.975 5(式中,y為H.pluvialis LUGU蝦青素的產(chǎn)量,x為lcy基因的表達(dá)水平,直線方程的相關(guān)系數(shù)為0.975 5)。
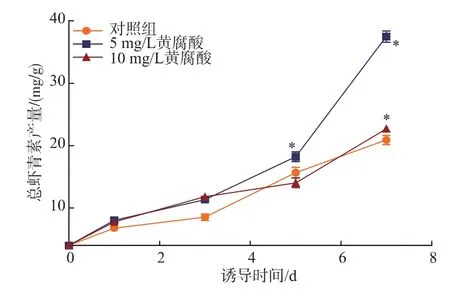
圖3 黃腐酸對(duì)雨生紅球藻總蝦青素產(chǎn)量的影響Fig.3 Effect of FA on the total astaxanthin production of H.pluvialis LUGU

圖5 不同質(zhì)量濃度的黃腐酸對(duì)lcy基因表達(dá)量的影響Fig.5 Effects of different level of FA on the transcript levels expression kinetics of lcy
3 結(jié)語
Raman等[13]利用不同的植物生長調(diào)節(jié)劑作為誘導(dǎo)子,誘導(dǎo)雨生紅球藻大量積累蝦青素。如將不同質(zhì)量濃度的茉莉酸甲酯等加入到處于生長對(duì)數(shù)期的藻液中,再轉(zhuǎn)入脅迫生長環(huán)境下誘導(dǎo)藻細(xì)胞積累蝦青素,發(fā)現(xiàn)50 mg/L的茉莉酸甲酯大大縮短了微藻合成蝦青素的時(shí)間,可有效促進(jìn)雨生紅球藻積累蝦青素,但高濃度的誘導(dǎo)劑可能對(duì)藻細(xì)胞有毒害作用。Gao和Lu等皆利用不同質(zhì)量濃度赤霉素A3誘導(dǎo)藻細(xì)胞積累蝦青素,結(jié)果表明,赤霉素A3作為誘導(dǎo)劑可以提高蝦青素的積累量,40 mg/L的赤霉素A3誘導(dǎo)藻細(xì)胞所積累的蝦青素,高于20 mg/L和2 mg/L的赤霉素A3誘導(dǎo)的積累量[14-15]。因此,探究合適的誘導(dǎo)子和恰當(dāng)?shù)膭┝渴强朔r青素產(chǎn)量低的有效途徑。Gao[16]研究了25 mg/L和50 mg/L的水楊酸分別對(duì)藻細(xì)胞積累蝦青素與相關(guān)基因表達(dá)量的影響,基因與蝦青素的積累正相關(guān)主要包括在轉(zhuǎn)錄前水平、轉(zhuǎn)錄水平及轉(zhuǎn)錄水平后;研究發(fā)現(xiàn)用25 mg/L誘導(dǎo)藻細(xì)胞時(shí),lcy基因的表達(dá)量在第3天達(dá)到最高,早與藻細(xì)胞開始快速積累蝦青素時(shí)間,表明這lcy在轉(zhuǎn)錄水平前對(duì)蝦青素生物合成起到了正調(diào)節(jié)的作用。因此,研究誘導(dǎo)過程中蝦青素代謝途徑中相關(guān)基因的表達(dá)水平,有助于了解在添加黃腐酸的條件下蝦青素產(chǎn)量提高的原因,為誘導(dǎo)蝦青素積累提供理論基礎(chǔ)。
黃腐酸是腐殖質(zhì)中水溶性的一種天然物質(zhì),含有很多植物所需要的微量元素和氨基酸等,所含有的大量官能團(tuán)生理活性比一般的植物生長激素生理能力強(qiáng),還可以絡(luò)合金屬離子[9],可作為植物生長因子促進(jìn)植物的生長發(fā)育[17]。作為植物的一種拮抗劑,已有研究指出黃腐酸對(duì)于植物的抗倒伏、抗旱具有一定的作用[18-19]。添加外源黃腐酸可激發(fā)植物防御基因的表達(dá),誘導(dǎo)植物的化學(xué)防御,促進(jìn)相關(guān)化合物的合成。蝦青素是雨生紅球藻在脅迫條件下產(chǎn)生和積累的次生代謝產(chǎn)物,因此研究FA對(duì)雨生紅球藻的生長以及蝦青素積累的影響有較好的前景。
適宜質(zhì)量濃度的FA有利于促進(jìn)雨生紅球藻H. pluvialis LUGU細(xì)胞的生長和蝦青素的積累。FA添加質(zhì)量濃度為10 mg/L,藻細(xì)胞的生物量產(chǎn)率達(dá)到最大80.28 mg/(L·d),比對(duì)照組提高5.44%。5 mg/L的FA,可誘導(dǎo)蝦青素產(chǎn)量提高了1.86倍;且lcy表達(dá)量達(dá)到最大值,為對(duì)照組的1.2倍,微藻蝦青素產(chǎn)量與lcy基因表達(dá)量成正相關(guān)。隨著FA誘導(dǎo)蝦青素生物合成的分子機(jī)制的進(jìn)一步研究,F(xiàn)A有望成為產(chǎn)蝦青素微藻的生物誘導(dǎo)子。
[1]MIKI W,HOSODA K,KONDO K,et al.Astaxanthin-containing Drink[J].Japanese Patent,1998,10155459.
[2]ZHUANG Huiru,CHEN Wenlie.The research of ultrastructure for different morphological cells of Haematococcus pluvialis[J]. Journal of Applied and Environmental Biology,2001,7(5):428-433.(in Chinese)
[3]QIAN Fei,LIU Haiying,GUO Shidong.The application of papain in astaxanthin extraction from crayfish shell[J].Journal of Food and Biotechnology,2010,29(2):237-243.(in Chinese)
[4]HU Jianzhong,GONG Jifan,DONG Qinglin.The mechanism of low nitrogen promoting astaxanthin biosynthesis in Phaffia rhodozyma[J].Journal of Food and Biotechnology,2009,28(1).(in Chinese)
[5]PAN C H,CHIEN Y H.Effects of dietary supplementation of alga Haematococcus pluvialis(Flotow),synthetic astaxanthin and β-carotene on survival,growth,and pigment distribution of red devil,Cichlasoma citrinellum (Günther)[J].Aquaculture Research,2009,40(8):871-879.
[6]KOP A,DURMAZ Y.The effect of synthetic and natural pigments on the colour of the cichlids(Cichlasoma severum sp.,Heckel 1840)[J].Aquaculture International,2008,16(2):117-122.
[7]GUERIN M,HUNTLEY M E,OLAIZOLA M.Haematococcus astaxanthin:applications for human health and nutrition[J]. Trends in Biotechnology,2003,21(5):210-216.
[8]STEINBRENNER J,LINDEN H.Light induction of carotenoid biosynthesis genes in the green alga Haematococcus pluvialis:regulation by photosynthetic redox control[J].Plant Molecular Biology,2003,52:343-356.
[9]LU Lingang.The yellow humic acid and its application in agriculture[J].Modernizing Agriculture,2001,5(9):10.(in Chinese)
[10]CHEN Yuling,ZHOU Xie.Effect of fulvic acid on ABA,IAA and activities of superoxide dismutase and peroxydase in winter wheat seedling under drought condition[J].Plant Physiology Communication,2000,36(4):311-314.(in Chinese)
[11]BOUSSIBA S,VONSHAK A.Astaxanthin accumulation in the green alga Haetoccus pluvialis[J].Plant Cell Physiol,1991(32):1077-1082.
[12]LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod [J].Methods,2001,25:402-408.
[13]RAMAN V,RAVI S.Effect of salicylic acid and methyl jasmonate on antioxidant systems of Haematococcus pluvialis[J].Acta Physiologiae Plantarum,2011,33(3):1043-1049.
[14]LU Y,JIANG P,LIU S,et al.Methyl jasmonate-or gibberellins A3-induced astaxanthin accumulation is associated with up-regulation of transcription of β-carotene ketolase genes(bkts)in microalga Haematococcus pluvialis[J].Bioresource Technology,2010,101(16):6468-6474.
[15]GAO Z,MENG C,GAO H,et al.Carotenoid genes transcriptional regulation for astaxanthin accumulation in fresh water unicellular alga Haematococcus pluvialis by gibberellin A3(GA3)[J].Indian Journal of Biochemistry&Biophysics,2013,50(6):548-553.
[16]GAO Z,MENG C,ZHANG X,et al.Induction of salicylic acid(SA)on transcriptional expression of eight carotenoid Xgenes and astaxanthin accumulation in Haematococcus pluvialis[J].Enzyme and Microbial Technology,2012,51(4):225-230.
[17]何立華.生物技術(shù)黃腐酸的研究和應(yīng)用[M].北京:化學(xué)工業(yè)出版社,1999.
[18]ZHOU Lina,SUN Lirong,MAO Hui,et al.Efects of drought-resistant fulvic acid liquid fertilizer on wheat and maize growth[J]. Agricultural Research in Arid Region,2012,30(1):154-158.(in Chinese)
[19]HUI Zhenlong,LI Chaozhou,SHI Wenxuan,et al.A study on the use of fulvic acid to improve growth and resistance in continuous cropping of potato[J].Acta Prataculturae Sinica,2013,22(4):130-13.(in Chinese)
Effects of Fulvic Acid on Astaxanthin Accumulation and the Transcript Levels Expression Kinetics of lcy Gene of Haematococcus pluvialis LUGU
SHANG Minmin, ZHAO Yongteng, ZHAO Peng, LI Tao, XU Junwei, YU Xuya*
(College of Life Science and Technology,Kunming University of Science and Technology,Kunming 650500,China)
To explore the effect of Fulvic acid (FA)on Haematococcus pluvialis,different concentration gradient of Fulvic acid(FA)was added in the culture of Haematococcus pluvialis LUGU.Meanwhile,cells growth,astaxanthin accumulation and the transcript levels expression kinetics of lcy gene was studied during induced time.The results show that the suitable concentration of FA not only promote the algae growth(when dealt with 10 mg L-1FA concentration,the algae biomass production reach the maximum 80.28 mg L-1d-1,increased by 5.44%than that of the control group),but also greatly improve the astaxanthin content(the astaxanthin content reach 20.82 mg L-1when dealt with 5 mg L-1FA concentration,is 86.89%higher than the control group).RT-PCR analysis shows that the transcriptional levels of lcy induced by 5 mg L-1and 10 mg L-1FA is 1.2 and0.7 times than that of control,respectively.The correlation analysis indicate that,the accumulation of astaxanthin content and the transcriptional levels of lcy gene inducted by FA is positively correlated.
fulvic acid,induce,astaxanthin,lcy gene,Haematococcus pluvialis LUGU
Q 81
:A
:1673—1689(2017)07—0702—05
2015-07-07
國家自然科學(xué)基金項(xiàng)目(21266013)。
*通信作者:余旭亞(1969—),男,云南昆明人,工學(xué)博士,教授,主要從事生物質(zhì)能工程研究。E-mail:xuya_yu@163.com
尚敏敏,趙永騰,趙鵬,等.黃腐酸對(duì)Haematococcus pluvialis LUGU蝦青素積累和lcy基因表達(dá)量的影響[J].食品與生物技術(shù)學(xué)報(bào),2017,36(07):702-706.
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國生殖健康(2019年2期)2019-08-23 08:12:08
聚氯乙烯(2018年9期)2018-02-18 01:11:34
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
