ClC-3基因敲除小鼠的構建及鑒定
2017-09-23 06:12:56余杰毛建文徐彬
世界復合醫學 2017年1期
關鍵詞:小鼠
余杰,毛建文,徐彬
1.廣東藥科大學生命科學與生物制藥學院,廣東廣州 510006;2.廣東藥科大學基礎學院,廣東廣州 510006
ClC-3基因敲除小鼠的構建及鑒定
余杰1,毛建文2,徐彬1
1.廣東藥科大學生命科學與生物制藥學院,廣東廣州 510006;2.廣東藥科大學基礎學院,廣東廣州 510006
目的 構建并鑒定ClC-3基因敲除小鼠,為研究ClC-3氯通道蛋白的生物學功能提供動物模型。 方法 將從2015年9月—2016年9月選取的6只賽業生物科技有限公司 (廣州)構建的ClC-3flox/+小鼠與2只Jackson實驗室引進的Ubc-Cre小鼠進行雜交繁殖,得到基因型為ClC-3flox/flox-Cre+及ClC-3flox/+-Cre+的小鼠。取鼠尾DNA,通過PCR法鑒定子代小鼠的基因型。結果 子代小鼠基因組DNA結果顯示在357 bp與100 bp處均有條帶,符合預期結果。 結論 該研究基于Cre/Loxp系統,利用loxp轉基因的ClC-3flox/+小鼠和在全身表達 Cre重組酶的Ubc-Cre小鼠,成功構建并鑒定了ClC-3基因敲除小鼠。
ClC-3;基因敲除;Cre/Loxp系統
課題組自 2000年開始從事 ClC-3蛋白的生物學功能研究,致力于獲得 ClC-3氯通道蛋白參與某些疾病的實驗證據。有研究表明ClC-3氯離子通道參與了細胞的轉運[1]、增殖[2-3]、遷移[4-8]、凋亡[9]等過程,另外,一些疾病的發生發展也與ClC-3有著密不可分的聯系[10-17],例如其在腫瘤組織中高表達[18-19],且在不同病理分級之間存在明顯差異[20]。因此,成功構建ClC-3基因敲除小鼠,為研究ClC-3基因與某些疾病的相關性提供動物模型具有重要的意義。現已有實驗室成功構建組織特異性ClC-3敲除小鼠模型用于疾病研究,例如,腸特異性敲除ClC-3會加重炎癥性腸病[20],心臟特異性敲除 ClC-3基因會導致成年鼠心肌肥厚。但目前還沒有實驗室構建條件性ClC-3敲除小鼠,該課題組從2015年9月—2016年9月,利用6只loxp轉基因的ClC-3flox/+小鼠和2只在全身表達 Cre重組酶的Ubc-Cre小鼠,成功繁育出條件性ClC-3敲除小鼠21只,為進一步研究ClC-3與某些疾病的相關性提供實驗條件。
1 材料與方法
1.1 實驗動物
實驗動物 Ubc-Cre工具鼠由美國Jackson實驗室引進 (Stock Number:007179,StrainName:129S.Cg-Tg(UBC-cre/ERT2)1Ejb/J,品系背景為C57BL/6J,雄性2只,基因型為雜合子(+/-)。ClC-3基因敲除小鼠委托賽業生物科技有限公司(廣州)構建,編號為001_mClcn3_1H12,品系背景為C57BL/6J,雌雄各3只,基因型為雜合子(+/-)。同時自中山大學實驗動物中心購進野生型C57BL/6J鼠雌性15只、雄性1只用于保種擴群。
1.2 實驗材料
鼠尾基因組DNA提取試劑盒 (目錄號:CW2094)購自康為世紀有限公司,Taq酶 (DRR037S)、DNA marker DL1000(D526A)、瓊脂糖(BIOWEST)購自大連寶生物工程有限公司,EB替代物溶液購自北京普利萊基因技術有限公司。小鼠鑒定引物由上海Invitrogen公司合成,見表1。
1.3 敲除小鼠的繁殖、信息記錄
3月齡雜合flox小鼠、Cre工具小鼠及野生型小鼠于SPF環境下飼養[SYXK(粵)2012-0125],紫外2 h/d,溫度22~28℃,濕度 40%~60%,明暗循環12 h/d,自由飲食和飲水。小鼠飼料及墊料均購自廣東藥科大學實驗動物中心,籠盒、墊料、飲用水均經高溫高壓滅菌處理。飼養過程中,進入動物房1次/d補充飼料和飲用水,觀察小鼠生長情況,墊料更換3次/周。小鼠2個月后性能力發育成熟,將其雌雄3:1同籠合養,定期給予滅菌葵花籽以增強其繁殖能力。雌性小鼠的妊娠期和哺乳期為3周左右,記錄其分娩時間、仔鼠數量,并于哺乳期后將小鼠與母鼠分開,雌雄分開飼養,待小鼠成年用于鑒定。flox雜合小鼠(ClC-3flox/+)自交所得F1代有flox純合小鼠(ClC-3flox/flox)、flox雜合小鼠(ClC-3flox/+)及野生型小鼠(ClC-3+/+),據孟德爾定律,概率分別為1/4、1/2、1/4。同時由flox雜合小鼠(ClC-3flox/+)與Cre工具小鼠(Cre+)交配獲得雜交系F1代有flo x雜合伴Cre陽性小鼠 (ClC-3flox/+-Cre+),flox雜合小鼠(ClC-3flox/+),Cre工具小鼠(Cre+),野生型小鼠(ClC-3+/+),概率分別為1/4。最后由所得的flox純合小鼠(ClC-3flox/flox)與 flox雜合伴 Cre陽性小鼠(ClC-3flox/+-Cre+)雜交得到flox純合伴 Cre陽性小鼠(ClC-3flox/flox-Cre+),flox雜合伴Cre陽性小鼠 (ClC-3flox/+-Cre+),flox純合小鼠(ClC-3flox/flox),flox雜合小鼠(ClC-3flox/+),概率分別為1/4。其中flox純合伴Cre陽性小鼠(ClC-3flox/flox-Cre+),flox雜合伴 Cre陽性小鼠(ClC-3flox/+-Cre+)即實驗所需。
1.4 小鼠的基因型鑒定
1.4.1 小鼠組織DNA的提取 將小鼠尾部組織小段 (長0.5~1.0 cm)置入1.5 mL Eppendolf管,按照TailGen DNA Kit鼠尾基因組DNA提取試劑盒 (CW2094)說明進行操作,提取DNA。
1.4.2 PCR反應 利用ClC-3基因敲除鼠和Ubc-Cre工具鼠特異性引物(如表1)進行PCR擴增。ClC-3基因敲除鼠PCR體系總體積25 μL,分別加入Premix Taq酶12.5 μL,PCR引物各0.8 μL,模板DNA1.5 μL,ddH2O 9.4 μL。注:引物濃度為10 μM。反應條件為:94℃預變性3 min,94℃變性30 s,退火60℃ 35 s,延伸72℃35 s,循環32次,72℃延伸5min,4℃保存。Ubc-Cre工具鼠PCR體系總體積12 μL,分別加入Premix Taq酶4.81 μL,目的基因及內參引物各1.2 μL,模板DNA2 μL,ddH2O 2.79 μL。注:引物濃度為10 μM。反應條件為:94℃預變性3 min,94℃變性30 s,退火51.7℃1 min,延伸72℃1 min,循環35次,72℃延伸2 min,4℃保存。
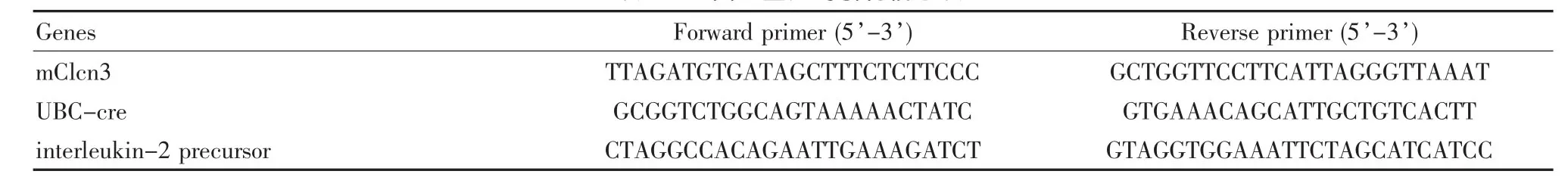
表1 基因型鑒定的引物信息表
1.4.3 瓊脂糖電泳及成像分析 取10 μLPCR產物進行3%的瓊脂糖凝膠電泳,凝膠成像系統分析結果。
2 結果
2.1 小鼠繁殖情況
成年ClC-3 flox母鼠與表達Cre重組酶的母鼠成功繁殖出子代幼鼠。每只母鼠孕期為19~21 d,哺乳期18~23 d,產后3周離乳。每胎平均產6~10只幼鼠,成活率>95%,新生小鼠呈肉色,無毛,閉眼,尾短,四肢柔弱,平均斷乳期21 d。成年小鼠成熟期:雄性10周,雌性8周,體重、活力相對C57BL/6J沒有較大差別,但繁殖能力相對弱,容易出現食仔、脫毛等現象。12個月共繁育小鼠2代/次,得到F2代小鼠39只,經基因型鑒定,獲得flox純合伴Cre陽性小鼠 (ClC-3flox/flox-Cre+)9只,flox雜合伴Cre陽性小鼠 (ClC-3flox/+-Cre+)12只,flox純合小鼠 (ClC-3flox/flox)8只,flox雜合小鼠(ClC-3flox/+)10只,純合小鼠比例為23%,基本符合孟德爾遺傳定律。
2.2 ClC-3基因敲除鼠鑒定結果
F2代基因敲除小鼠有 flox純合伴 Cre陽性小鼠(ClC-3flox/flox-Cre+),flox雜合伴 Cre陽性小鼠 (ClC-3flox/+-Cre+),flox純合小鼠 (ClC-3flox/flox),flox雜合小鼠(ClC-3flox/+)4種基因型。Loxp位點驗證,如圖 1所示,1、2、3、4、6號小鼠電泳條帶位于357 bp和287 bp處為雜合子;5、8號小鼠電泳條帶位于357 bp處為純合子;7號小鼠電泳條帶位于287 bp處為野生型。Cre位點驗證,如圖2所示,3、4、5、6、7號小鼠電泳條帶位于100 bp是為有Cre位點。綜上,實驗所需flox純合伴Cre陽性小鼠(ClC-3flox/flox-Cre+)為5號,flox雜合伴Cre陽性小鼠(ClC-3flox/+-Cre+)為3、4、6號。
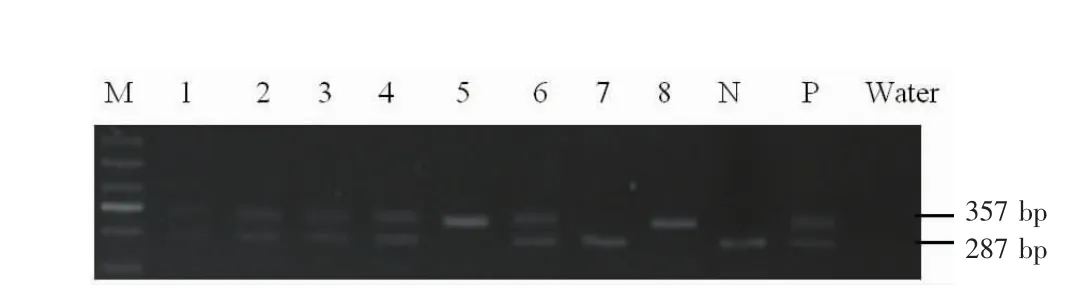
圖1 ClC-3 flox小鼠基因型鑒定結果

圖2 表達UBC-Cre重組酶的小鼠基因型的鑒定結果
2.3 小鼠生長發育情況
flox純合伴Cre陽性小鼠 (ClC-3flox/flox-Cre+),flox雜合伴Cre陽性小鼠(ClC-3flox/+-Cre+)毛色、體重、活力相對野生型C57BL/6 J整體沒有較大差別,見圖3。病理學檢查結果顯示,部分組織如腦、腎臟、脾臟的形態結構,flox純合伴 Cre陽性小鼠 (ClC-3flox/flox-Cre+)相對C57BL/6J未見明顯異常,見圖4。

圖3 不同基因型小鼠形態對比
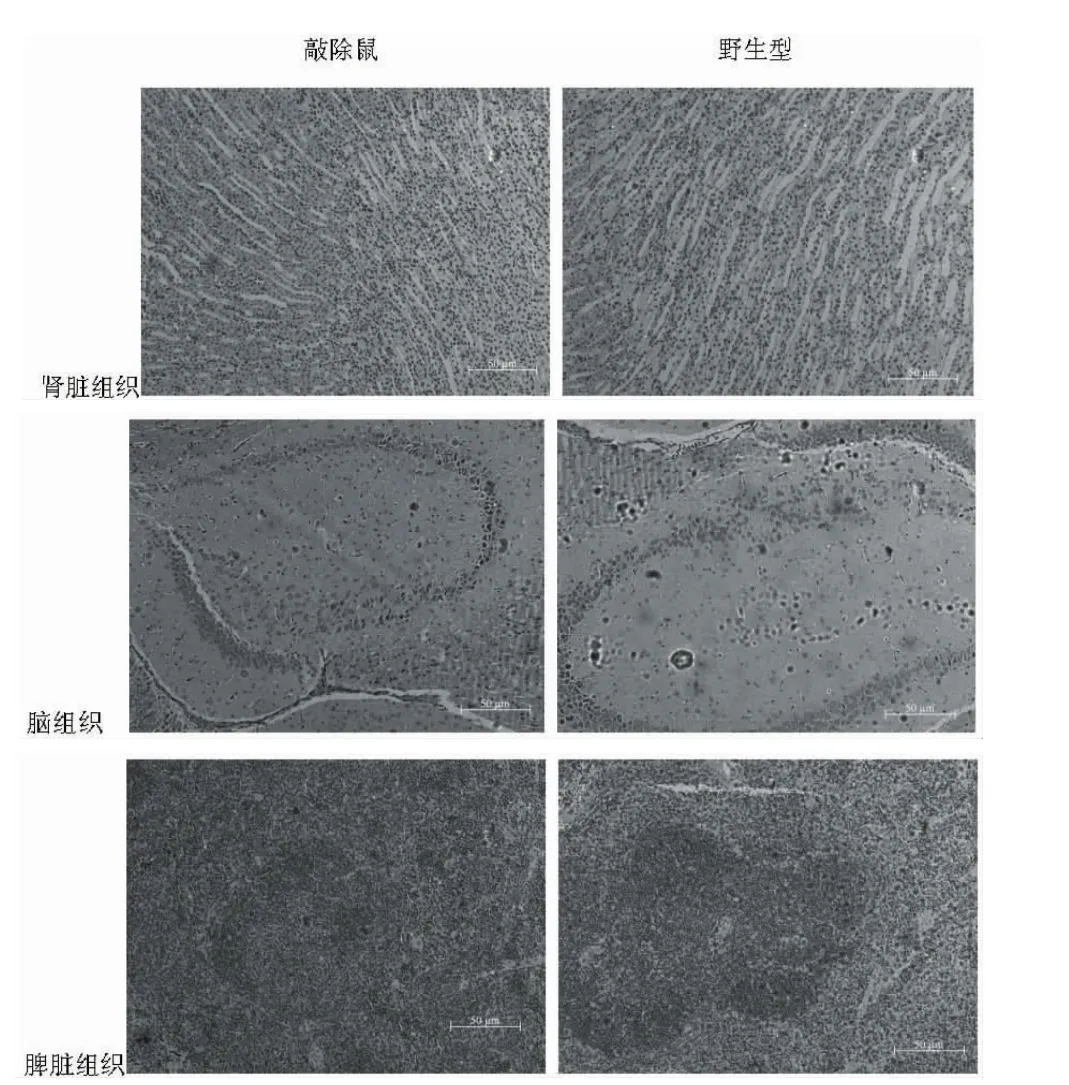
圖4 不同基因型小鼠的部分組織比較(HE,×100)
3 討論
隨著基因組學研究的深入,基因敲除技術日漸成熟,Zhen等[21]通過 RNA干擾技術成功抑制豬顆粒細胞中CEBPβ基因表達。2012年,余華等[22]利用雙臂同源重組法成功對銅綠假單胞菌內彈性蛋白基因進行了敲除。Liu等[23]采用 TALENs技術對對果蠅染色體上的基因進行了修飾。該文所述的條件性基因敲除是利用Cre/Loxp重組系統介導基因打靶的方法。Cre重組酶由Cre基因編碼,可識別特異DNA序列,如Loxp位點,切掉Loxp位點之間的靶基因和一個Loxp位點,從而達到靶基因敲除的目的。通過顯微注射法把兩個同向排列的LoxP位點插入到胚胎干細胞基因組靶位點的一個或幾個重要外顯子的兩端以制備出floxed小鼠,即文中所提ClC-3flox/+鼠,基因組DNA結果顯示在357 bp處有條帶,該floxed小鼠在與表達Cre重組酶小鼠雜交之前,該目的基因表達完全正常,若floxed小鼠與組織特異性表達Cre酶的小鼠進行雜交,可以在特定的組織或細胞中敲除該基因,而該基因在其它組織或細胞表達正常,該實驗所雜交對象為全身表達Cre重組酶的Ubc-Cre工具小鼠,DNA結果顯示在100bp處有條帶,可對ClC-3進行全身敲除。
ClC-3,基因符號為CLCN3,作為ClC電壓門控性氯離子通道家族中的重要一員,對于保持膜腔內電中性調節細胞體積有重要作用。有研究表明,心臟特異性敲除ClC-3基因,可以導致心肌肥大甚至心衰[24],此外,ClC-3在胃腸道黏膜保護、血管重塑[25]、成骨細胞的礦化和骨形成過程中也起到重要作用[26-27],也有研究表明某些基因的完全性敲除小鼠致死率極高[28],難以進行該基因功能的相關性研究。因此,構建全身敲除ClC-3基因的小鼠并于成年后誘導敲除對于進一步深入研究ClC-3的功能極為重要,該實驗構建的敲除小鼠未經他莫昔芬誘導時生長發育良好,且病理檢查未見異常,能夠用于后續實驗。根據Cre/Loxp條件性基因敲除技術原理,只有基因型為ClC-3 flox/flox-Cre+的純合小鼠于成年后經他莫西芬誘導ClC-3基因完全敲除,而其他類型的小鼠體內ClC-3基因則正常存在。例如基因型為ClC-3flox/+-Cre+的雜合子敲除小鼠仍有可能對某些細胞的功能產生影響。本實驗成功構建ClC-3基因敲除純合子、雜合子小鼠用于深入研究ClC-3的結構、功能及與某些疾病的相關性。
綜上所述,基于Cre/Loxp系統,利用loxp轉基因的ClC-3flox/+小鼠和在全身表達 Cre重組酶的Ubc-Cre小鼠,本實驗室成功構建ClC-3基因全身敲除小鼠,并通過PCR技術從基因表達水平上完成了鑒定。ClC-3全身敲除小鼠在生長速度、繁殖能力等方面與野生型小鼠無明顯差異。到目前為止培育雜交子代小鼠共20多只,下一步待小鼠發育成熟后通過他莫西芬誘導其ClC-3全身敲除,并利用該敲除鼠,探討局部給藥誘導實現組織特異性敲除,觀察ClC-3敲除對高表達ClC-3的組織功能是否有影響。
綜述所述,條件性ClC-3基因全身敲小鼠的成功構建,為ClC-3在組織器官生理病理的功能研究提供了良好的實驗材料基礎。
(
)
[1]Guzman RE,Miranda-Laferte E,Franzen A,et al.Neuronal ClC-3 splice variants differ in subcellular localizations,but mediate identicaltransportfunctions [J].JournalOf Biological Chemistry,2015,290(43):25851-25862.
[2]Mao J,Li X,Chen W,et al.Cell cycle-dependent subce llular distribution of ClC-3 in HeLa cells[J].Histochemistry And Cell Biology,2012,137(6):763-776.
[3]Liang W,Huang L,Zhao D,et al.Swelling-activated Cl currents and intracellular CLC-3 are involved in proliferation of human pulmonary artery smooth muscle cells[J].Journal Of Hypertension,2014,32(2):318-330.
[4]Guo R,Pan F,Tian Y,et al.Down-Regulation of ClC-3 Expression Reduces Epidermal Stem Cell Migration by Inhibiting Volume-Activated Chloride Currents[J].The Journal Of Membrane Biology,2016:1-12.
[5]Xu B,Jin X,Min L,et al.Chloride channel-3 promotes tumor metastasis by regulating membrane ruffling and is associated with poor survival[J].Oncotarget,2015,6(4):2434-2450.
[6]Guan Y,Huang Y,Wu J,et al.Overexpression of chloride channel-3 is associated with the increased migration and invasion ability of ectopic endometrial cells from patients with endometriosis[J].Human Reproduction,2016:dew034.
[7]Ma MM,Lin CX,Liu CZ,et al.Threonine532 phosphorylation in ClC-3 channels is required for angiotensin II-induced Cl?current and migration in cultured vascular smooth muscle cells[J].British Journal Of Pharmacology,2016,173 (3):529-544.
[8]Gaurav R,Bewtra AK,Agrawal DK.Chloride Channel 3 Channelsinthe Activationand MigrationofHumanBlood Eosi nophils in Allergic Asthma[J].American Journal Of Respir atory cell And Molecular Biology,2015,53(2):235-245.
[9] Liu J,Zhang FF,Li L,et al.ClC-3 deficiency prevents apoptosis induced by angiotensin II in endothelialprogenitorcellsvia inhibition ofNADPH oxidase[J]. Apoptosis,2013,18(10):1262-1273.
[10]Guzman RE,Alekov AK,Filippov M,et al.Involvement of ClC -3 chloride/proton exchangers in controlling glutamatergic synaptic strength in cultured hippocampal neurons[J].Frontiers In Cellular Neuroscience,2014:8.
[11]Zhang Y,Guo X,Guo J,et al.Lactobacillus casei reduces susceptibility to type 2 diabetes via microbiota-mediated body chloride ion influx[J].Scientific Reports,2014(4): 5654.
[12]Xiang N,Liu J,Liao Y,et al.Abrogating ClC-3 Inhibits LPS-induced Inflammation via Blocking the TLR4/NF-κB Pathway[J].Scientific Reports,2016:6.
[13] Liu SW,Li Y,Zou LL,et al.Chloride channels are involved in sperm motility and are downregulated in spermatozoa from patients with asthenozoospermia[J].Asian Journal Of Andrology,2016.
[14]Tao J,Liu C Z,Yang J,et al.ClC-3 deficiency prevents atherosclerotic lesion development in ApoE mice[J].Journal Of Molecular And Cellular Cardiology,2015(87):237-247.
[15]Huang YY,Huang XQ,Zhao LY,et al.ClC-3 deficiency protects preadipocytes against apoptosis induced by palmi tate in vitro and in type 2 diabetes mice[J].Apoptosis, 2014,19(11):1559-1570.
[16]Fan F,Liu T,Wang X,et al.ClC-3 Expression and Its Association with Hyperglycemia Induced HT22 Hippoc ampal Neuronal Cell Apoptosis[J].Journal Of Diabetes Res earch,2016,2016.
[17]Yan Y,Ding Y,Ming B,et al.Increase in Hypotonic Stress-Induced Endocytic Activity in Macrophages via ClC-3[J].Molecules And Cells,2014,37(5):418.
[18]Zhang H,Zhu L,Zuo W,et al.The ClC-3 chloride channel protein is a downstream target of cyclin D1 in nasopharyngeal carcinoma cells[J].The International Journal Of Biochemistry&Cell Biology,2013,45(3):672-683.
[19]Ye D,Luo H,Lai Z,et al.ClC-3 Chloride Channel Proteins Regulate the Cell Cycle by Up-regulating cyclin D1-CDK4/6 through Suppressing p21/p27 Expression in Nasopharyngeal Carcinoma Cells[J].Scientific Reports,2016, 6.
[20]Huang L Y,He Q,Liang S J,et al.ClC-3 chloride channel/ antiporter defect contributes to inflammatory bowel disease in humans and mice[J].Gut,2014:gutjnl-2013-305168.
[21]Zhen YH,Wang L,Riaz H,et al.Knockdown of CEBPβ by RNAi in porcine granulosa cells resulted in S phase cell cycle arrest and decreased progesterone and estradiol synthesis[J].The Journal Of Steroid Biochemistry And Mol ecular Biology,2014(214):90-98.
[22]余華,熊浚智,何曉梅,等.采用 Red重組系統敲除銅綠假單胞菌彈性蛋白酶基因[J].中國人獸共患病學報,2013, 29(2):129-132.
[23]Liu J,Li C,Yu Z,et al.Efficient and specific modifications of the Drosophila genome by means of an easy TALEN strategy[J].Journal Of Genetics And Genomics,2012,39(5): 209-215.
[24]Yu Y,Ye L,Li YG,et al.Heart-specific overexpression of the human short CLC-3 chloride channel isoform limits myocardial ischemia-induced ERP and QT prolongation[J]. International Journal Of Cardiology,2016(214):218-224.
[25]Zeng JW,Wang XG,Ma MM,et al.Integrin β3 mediates cerebrovascular remodelling through Src/ClC‐3 volume‐regulated Cl channel signalling pathway[J].British Journal Of Pharmacology,2014,171(13):3158-3170.
[26]Larrouture QC,Nelson DJ,Robinson LJ,et al.Chloridehydrogen antiporters ClC‐3 and ClC‐5 drive osteoblast mineralization and regulate fine‐structure bone patterning in vitro[J].Physiological Reports,2015,3(11):e12607.
[27]Wang H,Wang R,Wang Z,et al.ClC-3 chloride channel functions as a mechanically sensitive channel in osteob lasts[J].Biochemistry And Cell Biology,2015,93(6):558-565.
[28]Morgado-Palacin L,Varetti G,Llanos S,et al.Partial loss of Rpl11 in adult mice recapitulates Diamond-Blackfan Anemia and promotes lymphomagenesis[J].Cell Reports, 2015,13(4):712-722.
Construction and Identification of ClC-3 Knockout Mouse
YU Jie1,MAO Jian-wen2,XU Bin1
1.College of Life Science and Pharmaceutical Engineering,Guangdong Pharmaceutical University,Guangzhou,Guangdong Province,510006 China;2.Academic College,Guangdong Pharmaceutical University,Guangzhou,Guangdong Province,510006 China
Objective To construct and identify the ClC-3 knockout mouse and provide the animal model for researching the biology function of ClC-3 chloride channel protein.Methods 6 ClC-3flox/+mice constructed by the Saiye biotechnology Co.Ltd (Guangzhou)and 2 Ubc-Cre mice introduced by Jackson laboratory were selected from September 2015 to September 2016 were selected for interbreeding thus obtaining the lC-3flox/flox-Cre+and ClC-3flox/+-Cre+mice,Results The generation mice gene was identified by PCR method.The DNA results in the general mice gene group showed that there were stripes at 357 bp and 100 bp,meeting the expected results.Conclusion The paper successfully constructs and identifies the ClC-3 knockout mouse by using the loxp transgenosis ClC-3flox/+mice and Ubc-Cre mice with cre recombinase in expression of the whole body based on the Cre/Loxp system.
ClC-3;Knockout;Cre/Loxp system
R4
A doi 10.11966/j.issn.2095-994X.2017.03.01.04
2016-12-15;
2017-01-07
國家自然科學基金:ClC-3氯通道蛋白非離子通道途徑調控膜皺褶和有絲分裂的研究(31371144);廣東省自然科學基金:氯通道阻斷劑NPPB抑制術后腹膜粘連的研究(2016A030313741)。This work was supported by the National Natural Science Foundation of China(31371144)and the?Natural Science Foundation of Guangdong Province(2016A030313741).
余杰(1992.7-),女,湖北宜昌人,碩士,研究方向:離子通道蛋白功能及其靶向藥物篩選。
徐彬(1975.11-),女,四川眉山人,博士,教授,研究方向:離子通道蛋白功能及其靶向藥物篩選,腫瘤分子生物學與基因治療。E-mail:xubin2003@163.com。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
