溫度對雌雄葎草花芽分化和色素含量及光合作用影響的性別差異
2017-09-03 08:59:08郭海燕段婧劉金平游明鴻謝瑞娟
草業學報 2017年8期
關鍵詞:影響
郭海燕,段婧,劉金平*,游明鴻,謝瑞娟
(1.西華師范大學西南野生動植物資源保護省部共建教育部重點實驗室,四川 南充 637009;2.四川省草原科學研究院,四川 成都 611731)
溫度對雌雄葎草花芽分化和色素含量及光合作用影響的性別差異
郭海燕1,段婧1,劉金平1*,游明鴻2,謝瑞娟1
(1.西華師范大學西南野生動植物資源保護省部共建教育部重點實驗室,四川 南充 637009;2.四川省草原科學研究院,四川 成都 611731)
以雌雄異株攀援草本植物葎草為材料,依據野生資源主要分布地域的平均溫度,設置3個溫度為變量,通過測定營養生長期和生長積溫、葉數量與性狀、光合色素含量及生物量分配的變化,分析溫度對葎草光合物質基礎、光合速率和光合效益的影響,研究雌雄株的營養生長持續期、葉性狀、光合參數及生物量分配對溫度響應的性別差異。結果表明,溫度與性別對營養生長持續期和生長積溫均有極顯著影響(P<0.01),3個溫度下雄株花芽分化均顯著早于雌株(P<0.05);花芽分化主要受溫度影響,生長積溫主要受性別影響,20 ℃利于花芽分化,營養生長期長短與積溫大小無關;溫度對葉性狀有顯著影響(P<0.05),受影響順序為葉片數/株>單葉面積>葉寬>葉長,25 ℃時葉片數/株和單葉面積最大;光合色素含量無性別差異(P>0.05),溫度顯著影響除葉綠素b外的其他色素含量(P<0.05),順序為類胡蘿卜素>葉綠素a+b>葉綠素a/b>葉綠素a>葉綠素b,20 ℃時光合色素含量顯著低于其他溫度。性別對Pn、Gs和Tr有顯著影響(P<0.05),溫度對光合參數有顯著影響(P<0.05),影響順序為Pn>Ci>Tr>Gs,Pn、Ci和Tr主要受溫度影響,Gs主要受性別影響;雌株生物量及根分配顯著大于雄株(P<0.05),溫度對生物量累積及分配有極顯著影響(P<0.01),隨溫度下降單株生物量顯著下降(P<0.05),溫度對生物量分配影響順序為根>葉>莖,25 ℃時優先向根和莖分配,20 ℃時顯著降低了莖分配,15 ℃時顯著優先向葉分配。
葎草;雌雄異株;光合色素;光合作用;生物量
雌雄異株攀緣草本植物,集雌雄異株植物的性別分化性、攀緣植物對外界支持物的依賴性、草本植物的短壽命性等特點于一身,面臨脅迫生境時,要求在相對短暫的生活史中,充分適應與利用生境條件,采取復雜、完善、高效、系統的生長策略,才能在自然選擇中保證物種的延續。在環境脅迫下,雌雄異株植物維持種群穩定性的能力較為薄弱,光合作用能力、生物量分配、水分利用效率以及凋落物降解等方面都易受環境脅迫的影響[1]。應對環境脅迫時,雌雄個體生長、形態、生殖、分布以及抗逆性等方面表現出明顯的性別差異[2],常引起種群性比發生變化,種群繁殖能力下降,最終導致種群所在的生態系統的組成、結構和功能發生改變。雌雄異株攀援草本植物的性別分化性、對生境的依賴性及短壽命性,要求雌雄株采取更為完善、同步、協調、匹配的生長策略,致使自然界中該類植物的存留物種極少。
桑科葎草屬植物葎草(Humulusscandens)是一種可作藥用、飼用及生態治理等功能的材料[3],為南北各省區分布最廣的野生雌雄異株草本攀緣植物,是研究該類植物生殖分配與繁殖策略的理想材料。目前,關于葎草的研究主要集中在營養成分、藥物成分、飼喂效果評價及對其他生物的化感作用方面[4-6],對雌雄異株特性導致的雌雄株形態、種群性比、生殖投入、種子活性等方面進行了較為淺顯的研究[7-9],對生境脅迫下形態塑性、繁殖策略及應激機理進行了初步的研究[10-12]。溫度是植物生長的主要生境因子之一,不僅影響光合、呼吸、蒸騰作用,也影響有機物的合成、運輸和貯存,從而影響植物生存、生長及生殖過程。溫度高低及積溫對植物構件形成、花芽分化、胚子活力及種子發育起信號誘導作用,若雌雄個體不能同時對溫度做出應激反應,同時調整和改變生長策略,將影響植物營養生長和生殖生長,導致物種面臨巨大的存續風險。關于溫度對雌雄植物生長發育、生理應激及生殖過程影響的研究極少。
本研究依據葎草主要分布地域的平均溫度,采用人工控制生境因子的辦法,設置3個溫度為變量,通過測定雌雄株營養生長期和生長積溫、葉數量與性狀、光合色素含量及生物量分配的變化,分析溫度對葎草光合物質基礎、光合速率和光合效益的影響,研究雌雄株的營養生長持續期、葉性狀、光合參數及生物量分配對溫度響應的性別差異,以期為溫度下葎草個體發育、種群形成、適應度及種群擴繁等研究奠定基礎,為雌雄異株植物資源保護利用、生態系統可持續發展及社會經濟發展提供科學依據。
1 材料與方法
1.1 試驗材料及設計
于2015年3月,以野生葎草種子為材料,采用高34 cm、口徑50 cm花盆,以河沙∶腐殖土1∶1均勻混合為基質,按10粒/盆播種,共30盆。在25 ℃下培養,幼苗1~2對真葉時,按3株/盆定苗。隨機10盆為1組,分別移入15、20、25 ℃的人工氣候培養箱,設光照12 h/黑暗12 h,光照強度1300 lx,相對濕度32%進行培養。待花芽分化現花蕾可識別雌雄時,進行相關測定。
1.2 測定項目與方法
營養生長持續期:從真葉形成到完成花芽分化,可識別雌雄所需要的時間。
營養生長積溫:按K=N(T-C)計算,其中:K為完成營養生長所需要的總熱量;N為真葉形成到花芽分化的發育歷期;T為發育溫度;C為發育閾溫度(10 ℃)。
葉片性狀:測單株葉片數,用普托YMJ-C型葉面積儀測由上而下第3葉的葉長、葉寬及單葉面積。
光合特征參數:利用LI-6400XT光合儀,用LED-紅藍光源葉室,設置有效輻射為850 μmol/(m2·s),相對濕度為40%~70%,胞外CO2濃度為385~420 μmol/mol,于上午10:00-11:30,選第3葉中部,隨機取3點測凈光合速率(net photosynthetic rate,Pn),氣孔導度(stomatal conductance,Gs),胞間CO2濃度(intercellular CO2concentration,Ci)和蒸騰速率(transpiration rate,Tr)等光合參數,每點連續采集3個值。
光合色素含量:采用乙醇丙酮混合提取法[13],測定成熟葉片中葉綠素a 、葉綠素b含量,計算類胡蘿卜素、總葉綠素含量及葉綠素a/b。
生物量:每組隨機選雌、雄各3株,按根、莖、葉分離,分別裝入信封,在105 ℃下烘至恒重后計算單株生物量及各構件生物量分配比。
1.3 數據分析
采用SPSS 17.0軟件,進行Duncan多重比較、SNK檢驗(即q檢驗)和雙因素方差分析。
2 結果與分析
2.1 溫度對營養生長持續期和生長積溫的影響
葎草雌株的營養生長持續期和生長積溫顯著長于雄株(P<0.05)(表1),3個溫度下雄株花芽分化均顯著早于雌株(P<0.05)。20 ℃下,葎草營養生長持續期顯著短于25與15 ℃(P<0.05),持續期僅為25 ℃時的50%左右;這表明20 ℃最有利于葎草的花芽分化轉變為生殖生長,過高或過低溫度延長其營養生長期。25 ℃下,葎草營養生長積溫顯著高于其他溫度(P<0.05),20與15 ℃下,完成營養生長所需積溫無顯著差異。方差分析表明,溫度與性別對營養生長持續期和生長積溫均有極顯著影響(P<0.01)(表2)。由F值可見,營養生長持續期主要受溫度影響,生長積溫主要受性別影響,溫度和性別互作對營養生長持續期影響大于對生長積溫的影響,致使葎草的營養生長持續期比生長積溫呈現出更大的差異。
2.2 溫度對葉性狀的影響
葎草雌株的葉長顯著大于雄株(P<0.05)(表1),雌雄株的葉片數/株、葉寬、單葉面積均無顯著差異(P>0.05)。溫度對葎草葉性狀有顯著影響(P<0.05),25 ℃下葎草的葉片數/株、葉長、葉寬、單葉面積均顯著大于其他溫度,葉性狀在20與15 ℃間無顯著差異。進一步方差分析(表2)表明,性別間葉長有極顯著差異(P<0.01)(表2),溫度間葉片數/株、葉寬、單葉面積有極顯著差異(P<0.01)。F值表明,溫度對葉性狀影響順序為:葉片數/株>單葉面積>葉寬>葉長。性別與溫度共同作用使葉性狀呈現出極顯著差異(P<0.01)。
2.3 溫度對光合色素含量影響
葎草葉片內光合色素含量在雌雄性別間無顯著差異(P>0.05)(表3),3個溫度下雄株葉片中葉綠素b(Chl b)和類胡蘿卜素(carotenoid,Caro)含量均略大于雌株。溫度對葉綠素a(Chl a)和Caro含量有顯著影響(P<0.05),對Chl b含量 無顯著影響,而Caro含量隨溫度降低而顯著下降(P<0.05)。20 ℃下,葉片中Chl a、Chl (a+b)含量及Chl a/Chl b顯著低于其他溫度。方差分析表明,性別間光合色素含量差異較小,溫度間光合色素含量除Chl b含量外,均有顯著差異(表4)。溫度對光合色素影響順序為Caro>Chl (a+b)>Chl a/b>Chl a>Chl b。性別與溫度互作對葎草光合色素含量無顯著影響。
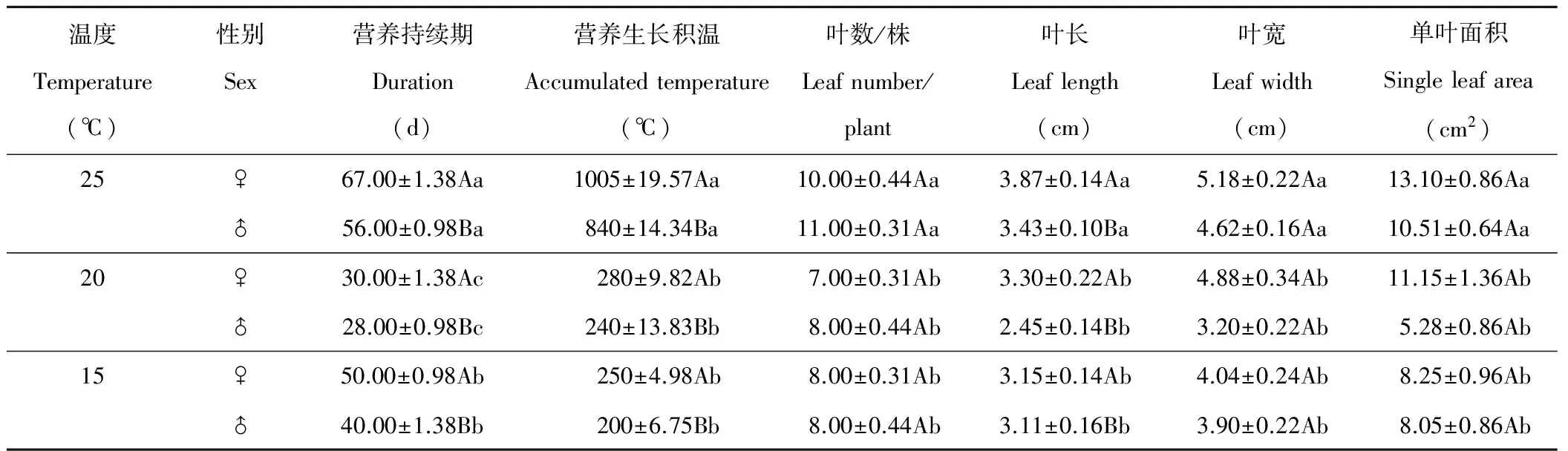
表1 溫度對營養期和葉性狀影響的SNK檢驗
♀:雌株Female;♂:雄株Male.同列不同大寫字母表示性別間差異顯著(P<0.05),不同小寫字母表示溫度間差異顯著(P<0.05)。下同。Different capital letters in the same column indicate significant differences between genders atP<0.05 level and different lowercase letters within the same column indicate significant differences among temperatures atP<0.05 level. The same below.
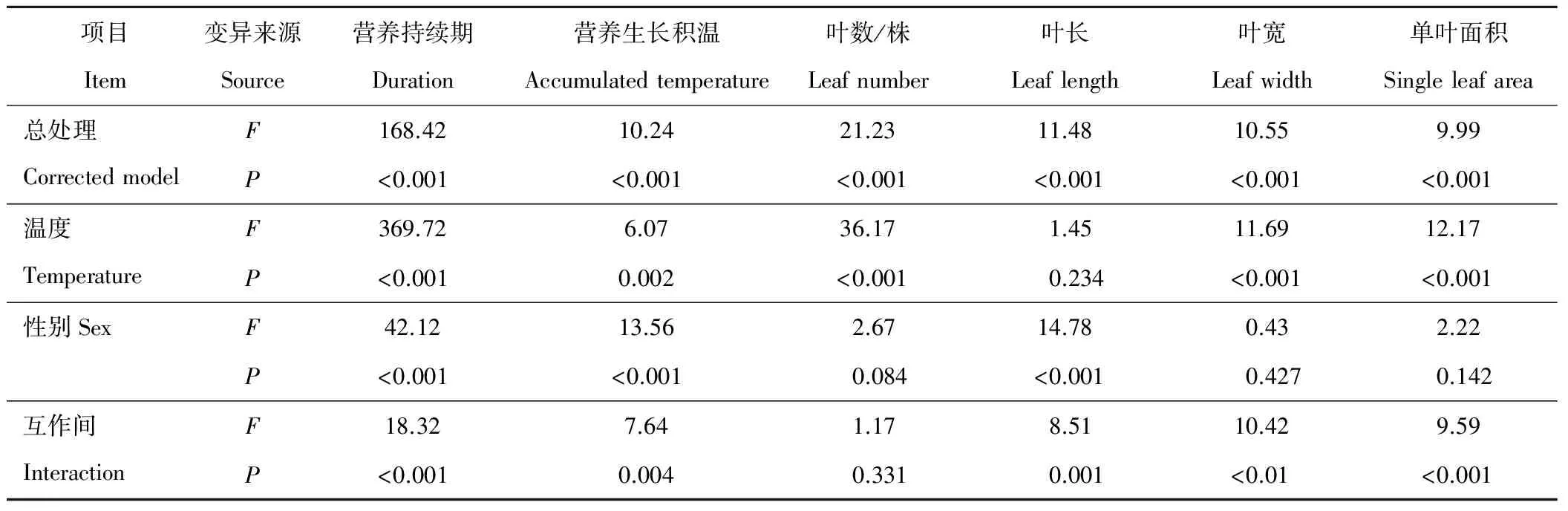
表2 營養生長期和葉性狀差異的雙因子方差分析
注:F值表示F檢驗的顯著性,F越大表示越顯著,P值表示概率值。下同。
Note:Fvalue indicates the significance ofFtest, with greaterFvalue meaning more significant.Pvalue indicates the probability value. The same below.
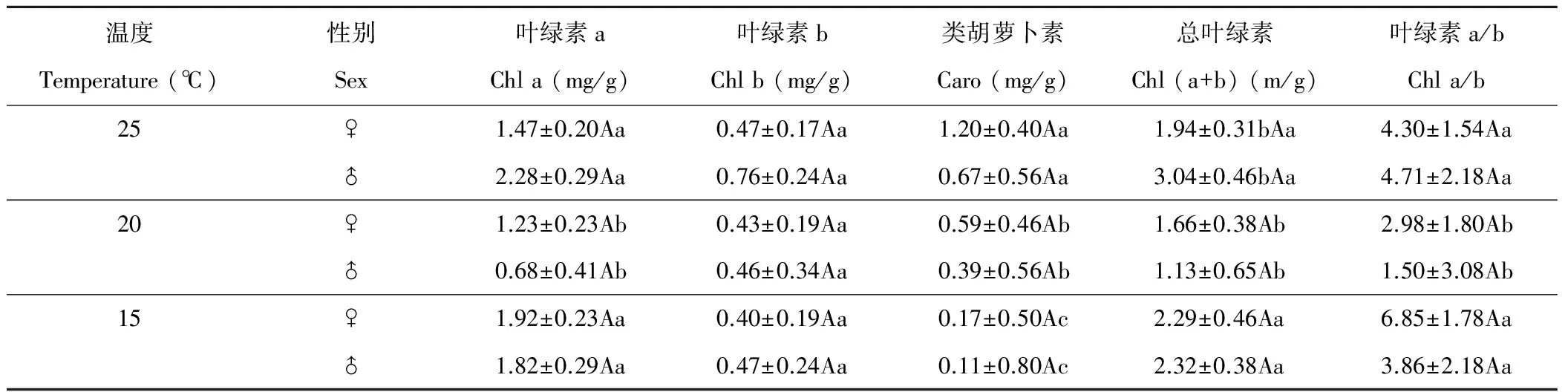
表3 溫度對光合色素含量影響的SNK檢驗
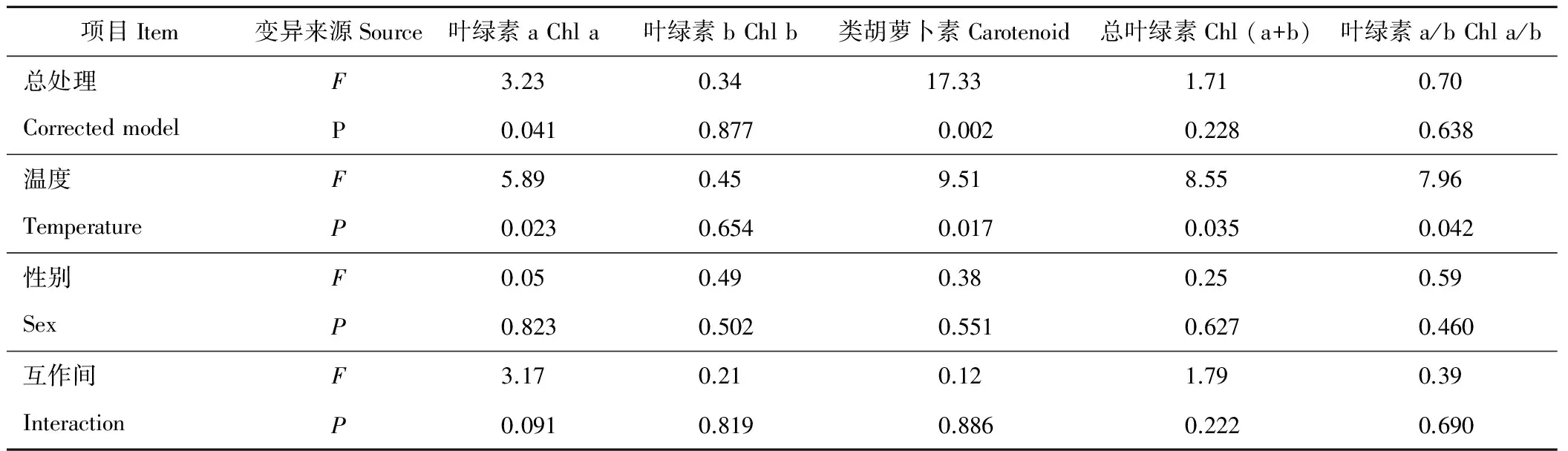
表4 光合色素含量差異的雙因子方差分析
2.4 溫度對光合參數影響
雌株葉片的凈光合速率(Pn)、氣孔導度(Gs)和蒸騰速率(Tr)顯著大于雄株(P<0.05)(表5),雌雄株葉片的胞間CO2濃度(Ci)無顯著差異。溫度對Pn、Gs、Ci和Tr均有顯著影響(P<0.05),20 ℃時,Pn、Gs和Tr值顯著低于其他溫度。Pn在15 ℃達最大值,為20 ℃時的2倍左右。Gs、Ci和Tr在25 ℃均達最大值,隨溫度下降呈現出不同的變化趨勢,Ci隨溫度下降而逐步下降,Gs和Tr則先顯著下降再顯著增加(P<0.05)。
方差分析表明,性別間Pn、Gs和Tr有顯著差異(P<0.05)(表6),差異大小為Tr>Pn>Gs。溫度間Pn、Ci和Tr有極顯著差異(P<0.01),Gs有顯著差異(P<0.05)。溫度對光合參數的影響順序為Pn>Ci>Tr>Gs。光合參數在性別與溫度互作間均有顯著差異(P<0.05)。葎草Pn、Ci和Tr受溫度的影響大于受性別的影響,Gs受性別影響大于溫度影響,而Ci僅受溫度影響。
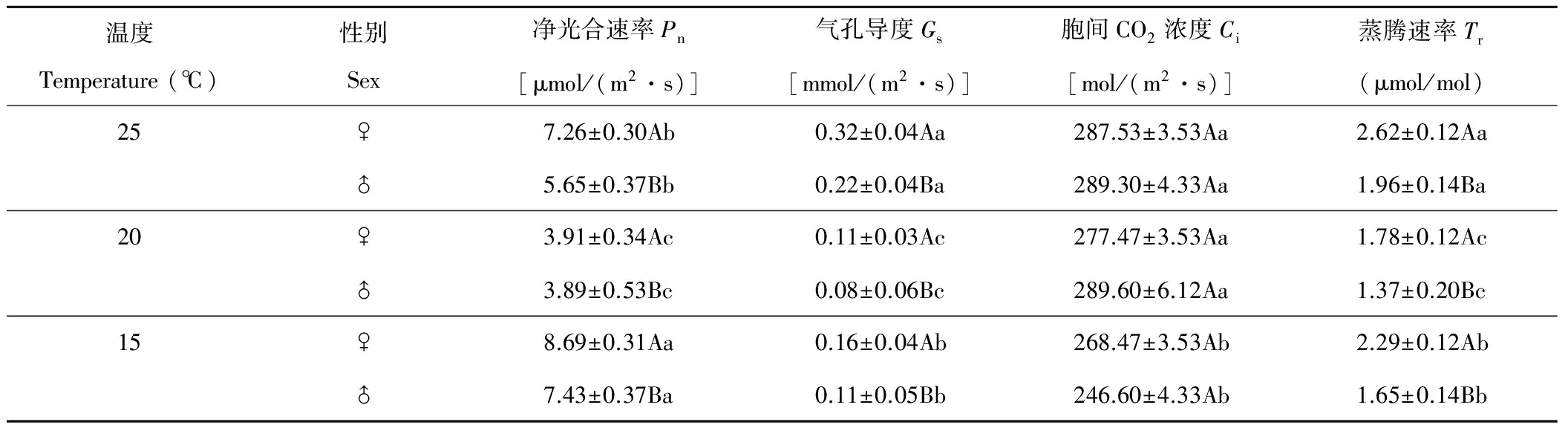
表5 溫度對光合參數影響的SNK檢驗
Pn: Net photosynthetic rate;Gs: Stomatal conductance;Ci: Intercellular CO2concentration;Tr: Transpiration rate. 下同 The same below.
2.5 溫度對生物量的影響
雌株的生物量顯著大于雄株(P<0.05)(表7),雌株生物量根分配顯著大于雄株(P<0.05),莖、葉分配受性別影響較少(P>0.05)。隨溫度下降單株生物量顯著下降(P<0.05),15 ℃時單株生物量比25 ℃時下降了55%左右。溫度對生物量分配有顯著影響,25 ℃時生物量優先向根和莖分配,20 ℃時顯著降低了莖分配,15 ℃時顯著降低根、莖生物量分配而優先向葉分配。
方差分析(表8)表明,性別間單株生物量和根分配有極顯著差異(P<0.01),溫度對生物量累積及分配的影響大于性別。溫度間單株生物量及構件分配比均有極顯著差異(P<0.01),溫度對生物量分配的影響順序為根>葉>莖。性別與溫度互作對生物量分配有極顯著影響,葉分配受互作間影響最大。
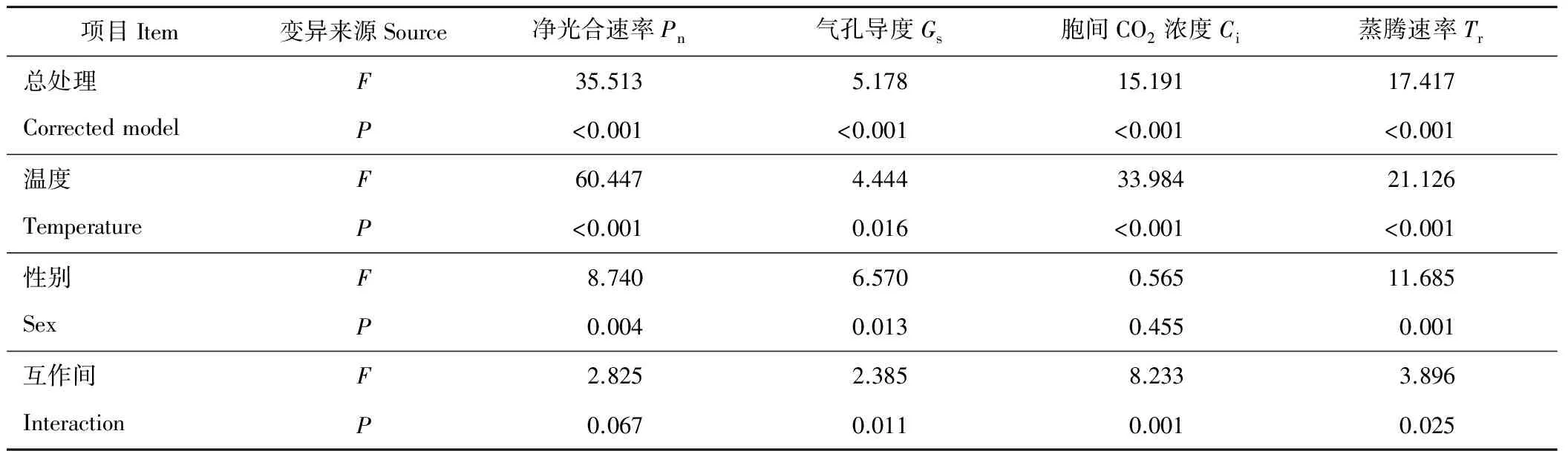
表6 光合參數差異的雙因子方差分析
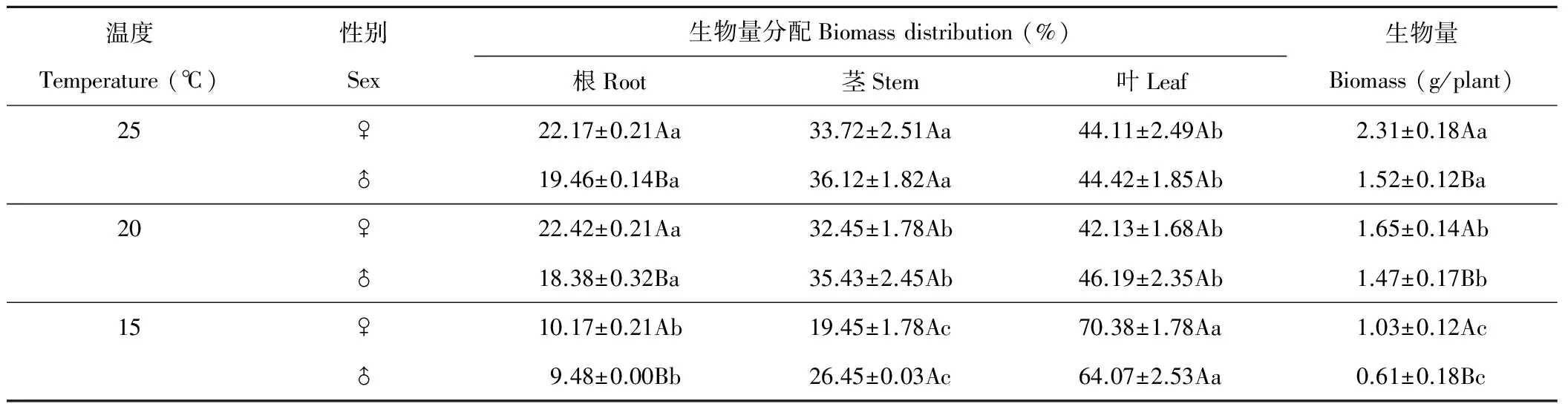
表7 溫度對生物量影響的SNK檢驗
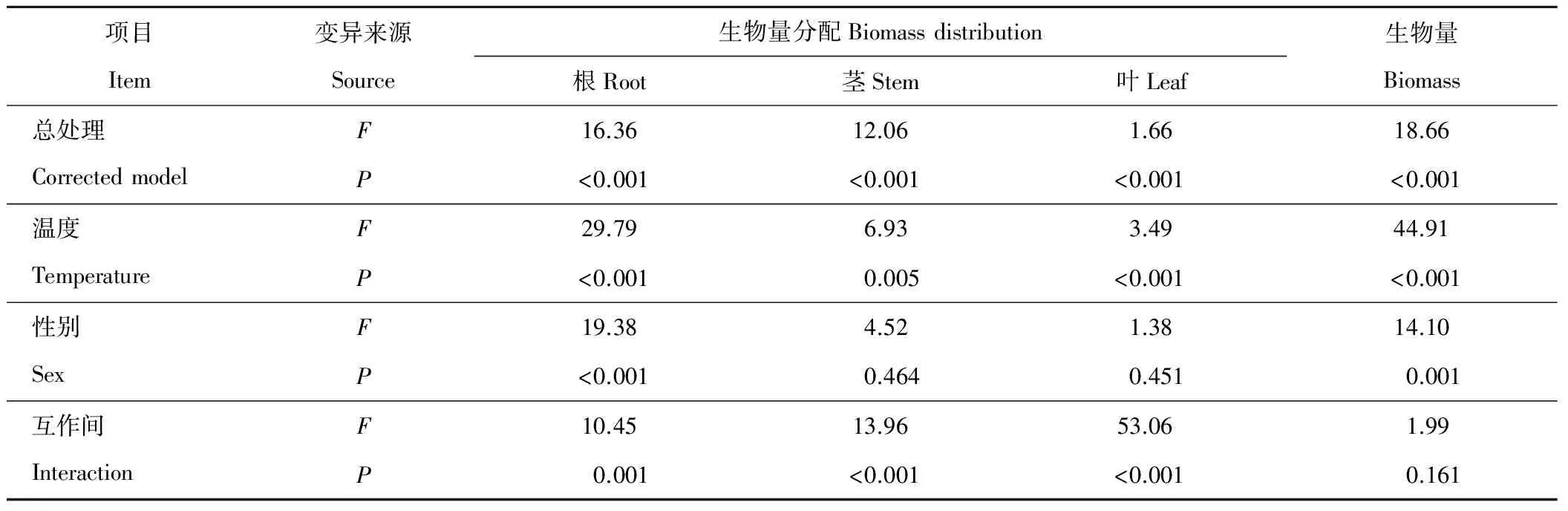
表8 生物量差異的雙因子方差分析
3 討論
3.1 溫度對花芽分化的影響
溫度是影響植物生長發育的主要生境因子之一,既可通過改變植物體內生理活性物質(酶、激素、維生素)的含量與活性,影響同化、異化等新陳代謝過程,導致構件數量和質量性狀、生物量累積與分配發生改變;還可通過影響土溫、氣溫,改變植物對水、肥的吸收和輸導效率,從而影響植物生長發育。尤其三基點溫度直接決定植物的分布區域、生長發育速度、光合潛力與產量潛力。野生葎草能在高寒地區之外的廣大區域分布,表明具有極強的溫度適應能力,≥10 ℃年積溫830 ℃即可完成生活史,但對其三基點溫度未見詳細報道。花芽分化是營養生長向生殖生長的轉變的生理和形態標志,而溫度是控制植物由營養生長向生殖生長轉變的臨界因子。大量事實表明,植物的成花有一定溫度需求。按有效積溫法則,完成營養生長期應該達到某一有效積溫。同一品種植物經歷的低溫時數越長, 達到開花所需的有效積溫越少[14]。本試驗中,不同溫度下葎草花芽分化需要的有效積溫差異極大,25 ℃積溫達1000 ℃是15 ℃積溫的近4倍,或許葎草完成營養生長所需積溫依據生境條件發生變化。野外觀察發現,生境良好的種群出苗當年只進行營養生長,而生境不良種群,營養生長期很短、生殖生長持續150 d左右[15]。但3個溫度下,雌株花芽分化需要積溫顯著高于雄株,或許雌株比雄株有更高的繁殖成本[16],致使雌株通過延長營養生長期累積更多的物質貯藏為生殖成功提供保障。一般認為花芽分化需要低溫的春化作用,本試驗在3個恒溫條件下,葎草雌雄株都順利完成了花芽分化,且野外觀察發現葎草在南充地區為一年生或多年生,2月中旬出苗或恢復生長,5月初開花,11下旬仍有開花,5月中旬到11月上旬均有果實成熟,表明葎草開花無需春化作用,同時對光周期不敏感。本研究中20 ℃時葎草營養生長期最短,花芽分化積溫較低。葎草營養生長持續期和花芽分化積溫對生境溫度的適應性調節,尤其雄株花芽分化所需積溫和時間顯著短于雌株,使雄株先于雌株進入生殖生長,高位的無限花序持續性地提供大量花粉,為傳花授粉奠定了基礎,雌株開花便能獲取足夠的受精機會[17],這對性比極易失衡的雌雄異株植物完成生殖過程及延續種群具有重要的意義。
3.2 溫度對光合物質基礎的影響
葉片是光合作用的主要器官,葉片數量、葉面積及光合色素含量是光合作用的物質基礎。光合作用實質是植物在系統性酶等生理活性物質促進下,經過復雜的系列反應把光能轉變為化學能的同化過程,最終體現為植物生物量的積累和分配。本研究中溫度對葎草葉性狀有顯著影響,葉片數/株、單葉面積隨溫度降低而減少,降到20 ℃后葉性狀下降幅度放緩。可見葉片形成率、葉面積擴展速度和葉片生長周期長均受溫度影響,25 ℃時利于葉片分化和擴展。葉長在性別間有極顯著差異,3個溫度下雌株葉長均顯著大于雄株,加之營養生長期長于雄株,使其葉片光合葉基礎及光合時期大于雄株。雖葉片內光合色素密度在性別間無顯著差異,而雌株葉片的鮮重和干重顯著大于雄株[18],使雌株光合作用所需有效色素含量顯著高于雄株,與干旱[15]和鉻脅迫[19]下的研究結果一致。溫度對光合色素含量有顯著影響,不同種類色素隨溫度降低含量變化的趨勢不同,吸收光能的葉綠素a含量先減后增,傳遞光能的葉綠素b含量相對穩定,吸收藍紫光的類胡蘿卜素則逐步下降,20 ℃時Chl a、Chl (a+b)含量及Chl a/Chl b顯著低于其他溫度。有報道認為在低溫環境脅迫下,由于葉片細胞內酶活性降低,葉綠素合成受抑制并導致葉綠素降解[20]使葉綠素含量降低。葎草15 ℃時葉綠素含量顯著高于20 ℃,可能是15 ℃未達導致葉綠素降解的低溫強度,同時15 ℃限制了葉面積拓展,而葉綠體等細胞器已經形成,造成葉綠素密度增加。15 ℃時類胡蘿卜素含量僅為25 ℃的15%左右,一方面可能類胡蘿卜素合成需要較高溫度,低溫引起類胡蘿卜素分解,另一方面可能類胡蘿卜素參與抵制低溫產生自由基的抗氧化活動,導致類胡蘿卜素含量隨溫度降低而顯著下降。葉綠素含量隨溫度變化無顯著性別差異,在本試驗中雄株的Chl a、Chl b、Chl (a+b)含量變異系數大于雌株。需擴大試驗溫度范圍,進一步研究雌雄株光合物質基礎對溫度脅迫的性別差異。
3.3 溫度對光合參數的影響
溫度對光合作用的影響較為復雜,主要通過酶的活性和氣孔開合度,影響酶促反應速度和氣體通量,致使光合作用底物濃度和反應速度發生改變。本試驗控制光照強度、CO2濃度,溫度作為變量下進行,表明溫度對葎草光合作用有顯著影響。常通過胞間CO2濃度(Ci)、氣孔導度(Gs)數值變化一致性,判斷氣孔是否參與光合速率的調節[21]。葎草在20~25 ℃溫度范圍內,胞間CO2濃度(Ci)、氣孔導度(Gs)和蒸騰速率(Tr)隨溫度而降低,20 ℃時Pn、Gs和Tr達最小值,說明該溫度區間氣孔因素對光合作用有調節作用。在15~20 ℃,Ci值下降而Gs值上升,推測在此溫度區間光合作用速率與氣孔因素無關而與葉肉細胞光合作用有關。按理說在適合植物生長的溫度范圍內,隨溫度增高酶活性提高和酶促反應速度應該加快,光合作用速率應該上升,溫度上升10 ℃,光合速率可提高一倍左右。本試驗中,15 ℃時凈光合速率(Pn)竟達最大值,或許此溫度下葉綠素濃度高是原因之一。葎草長期在低溫環境下,或許形成了更為復雜系統的光合策略,是否光合測定時所用的人工光源,暫時刺激了15 ℃下葎草的光合補償機制或光合潛力,其單株生物量僅為25 ℃時的50%左右。3個溫度下凈光合速率(Pn)、氣孔導度(Gs)和蒸騰速率(Tr)均有顯著性別差異,表明雌雄株光合作用對溫度變化響應方式不同。諸多研究表明,良好生境下雌株光合能力強于雄株[22-23],而在不良生境下雄株表現出更強的光合適應性[15,19,24-25]。3個溫度下雌株的光合速率總高于雄株,雌株的生物量總大于雄株,雌株根分配總多于雄株,致使其同化能力、貯存能力及抗逆能力提高,為雌株生殖生長及孕育種實提供了基礎。關于溫度對生殖投入、生殖過程及生殖收益待進一步研究。
4 結論
葎草對溫度具有極強的適應能力,在較低的有效積溫就可完成營養生長(約200 ℃),無需春化作用就可花芽分化;溫度對營養生長持續期有顯著影響,20 ℃利于花芽分化,但營養生長期長短與積溫大小無關;3個溫度下雄株都先于雌株花芽分化,為盡早提供雄胚子奠定基礎,雌株則通過延長營養生長期累積更多的物質貯藏為生殖成功提供保障;葎草可通過控制葉片數/株和單葉面積適應溫度條件,同時改變光合色素含量適應不同溫度下的光合需求;雌株光合葉面積、光合色素含量及氣孔導度高于雄株,其光合效率、生物量累積大于雄株;雌雄個體能依據溫度變化,協調一致的調整和改變生長策略,順利完成營養生長向生殖生長轉變,為生殖生長、種子形成及物種延續奠定基礎。
References:
[1] Xu X, Yang F, Yin C Y,etal. Research advances in sex-specific responses of dioecious plants to environmental stresses. Chinese Journal of Applied Ecology, 2007, 18(11): 2626-2631. 胥曉, 揚帆, 尹春英, 等. 雌雄異株植物對環境脅迫響應的性別差異研究進展. 應用生態學報, 2007, 18(11): 2626-2631.
[2] Yin C Y, Li C Y. Gender differences of dioecious plants related sex ratio recent advances and future prospects. Chinese Journal of Applied & Environmental Biology, 2007, 13(3): 419-425. 尹春英, 李春陽. 雌雄異株植物與性別比例相關的性別差異研究現狀與展望. 應用與環境生物學報, 2007, 13(3): 419-425.
[3] Sun Y C, Xu T B, Wang Y L. The comprehensive development and utilization ofhumulusscandens. Quarterly of Forest By-product and Speciality in China, 2003, (3): 57. 孫躍春, 徐彤寶, 王艷玲. 葎草的綜合開發利用. 中國林副特產, 2003, (3): 57.
[4] Zhang J, Zhang K, Liu X G,etal. Identification of crude drugs from genusLeonurus. Journal of Chinese Medicinal Materials, 1998, 21(12): 605-609. 張劫, 張鯤, 劉湘桂, 等. 葎草的形態組織鑒定. 中藥材, 1998, 21(12): 605-609.
[5] Li S Y, Ren X Q. Can be fed to livestock and poultry ofHumulusscandens. Animal Science Abroad, 2003, 30(6): 48-49. 李守陽, 任相泉. 葎草飼喂畜禽. 中國畜牧獸醫, 2003, 30(6): 48-49.
[6] Lei Q J, Diao Z M, Zhang Z G,etal. The development and utilization ofHumulusscandensand countermeasures research. Anhui Agricultural Science Bulletin, 2005, 11(7): 68-69. 雷青娟, 刁治民, 張正國, 等. 葎草的開發利用及防治對策的研究. 安徽農學通報, 2005, 11(7): 68-69.
[7] Liu J P, Wang Y. Asexual reproduction component and its diversity of wildHumulusscandensin Nanchong. Pratacultural Science, 2009, 26(8): 68-71. 劉金平, 王艷. 南充地區野生葎草構件性狀多樣性分析. 草業科學, 2009, 26(8): 68-71.
[8] Liu J P, Kang J L. Variation analysis of sexual reproductive investment for wildHumulusscandens(lour) merr population. Southwest China Journal of Agricultural Sciences, 2010, 23(3): 872-875. 劉金平, 康軍利. 野生葎草種群有性生殖投入與收益差異性分析. 西南農業學報, 2010, 23(3): 872-875.
[9] Liu J P, Ouyang M. Effect of maturity degree and storage time on the seed germinating ability of wildHumulusscandens. Journal of Anhui Agricultural Sciences, 2009, 38(4): 1817-1818. 劉金平, 歐陽梅. 成熟度與貯藏期對野生葎草種子發芽力的影響. 安徽農業科學, 2009, 38(4): 1817-1818.
[10] Liu J P, You M H, Zhang L H,etal. External supports affect the photosynthetic characteristics and biomass allocation of the climbing plantHumulusscandens. Acta Ecologica Sinica, 2015, 35(18): 6032-6040. 劉金平, 游明鴻, 張麗慧, 等. 不同支持物對攀援植物—葎草雌雄株光合特性及生物量結構的影響. 生態學報, 2015, 35(18): 6032-6040.
[11] Liu J P, Duan J.Humulusscandensgender differences in response to water stress in the vegetative growth stage. Acta Prataculturae Sinica, 2013, 22(2): 243-249. 劉金平, 段婧. 營養生長期雌雄葎草表觀性狀對水分脅迫響應的性別差異. 草業學報, 2013, 22(2): 243-249.
[12] Liu J P, You M H, Duan J,etal. Plasticity of reproductive strategy of dioecious humulus scandens in response to variation in water deficit stress. Acta Prataculturae Sinica, 2015, 24(3): 226-232. 劉金平, 游明鴻, 段婧, 等. 水分脅迫下雌雄異株植物葎草繁殖策略的可塑性調節. 草業學報, 2015, 24(3): 226-232.
[13] Xiong Q E. Plant Physiology Experiment Teaching Materials[M]. Chengdu: Sichuan Science and Technology Publishing House, 2003. 熊慶娥. 植物生理實驗教材[M]. 成都: 四川科學技術出版社, 2003.
[14] Zhang G R. Study on the effect of availably accumulative temperature on flower forcing ofChaenomelesjaponica. Northern Horticulture, 2008, (12): 123-124. 張桂榮. 有效積溫對日本海棠催花的影響. 北方園藝, 2008, (12): 123-124.
[15] Duan J, Liu J P. Gender differences ofHumulusscandensin response to drought stress during the vegetative growth stage. Hubei Agricultural Sciences, 2014, 52(13): 3078-3082. 段婧, 劉金平. 營養生長期葎草對干旱脅迫應對機理的性別差異分析. 湖北農業科學, 2014, 52(13): 3078-3082.
[16] Fan X, Wang D W, Liu J P. Gender differences in morphological plasticity and reproductive strategy of Humulus scan-dens in response to different external supports. Chinese Journal of Ecology, 2015, 34(6): 1559-1564. 樊星, 王大偉, 劉金平. 支持物對攀援葎草分枝形態塑性和繁殖對策影響的性別差異. 生態學雜志, 2015, 34(6): 1559-1564.
[17] Fan X, Wang D W, Fan X,etal. Effect of external support on the morphological plasticity and reproductive strategies of climbingHumulusscandens. Pratacultural Science, 2015, 32(9): 1457-1462. 樊星, 王大偉, 范宣, 等. 支持物改變葎草形態可塑性和繁殖特性. 草業科學, 2015, 32(9): 1457-1462.
[18] Duan J, Liu J P. Gender differences analysis of apparent traits ofHumulusscandensin response to temperature in the vegetative growth stage. Pratacultural Science, 2013, 30(3): 418-422. 段婧, 劉金平. 不同溫度下雌雄葎草營養生長期的生長特性. 草業科學, 2013, 30(3): 418-422.
[19] Wang B X, Xiao J, Feng X,etal. Effects of chromium stress on physiological and ecophysiological characteristics ofHumulusscandens. Acta Prataculturae Sinica, 2016, 25(7): 131-139. 王碧霞, 肖娟, 馮旭, 等. 鉻脅迫對葎草雌雄植株光合生理特性的不同影響. 草業學報, 2016, 25(7): 131-139.
[20] Miao W, Wang G J, Ma D R,etal. Physiological responses of weedy rice to cold stress at seeding in Liaoning Province, China. Chinese Journal of Rice Science, 2011, 25(6): 639-644. 苗微, 王國驕, 馬殿榮, 等. 遼寧省雜草稻幼苗對低溫脅迫的生理響應. 中國水稻科學, 2011, 25(6): 639-644.
[21] Xu D Q, Zhang Y Z. Photoinhibition of photosynthesis in plants. Plant Physiology Communications, 1992, 28(4): 237-243. 許大權, 張玉忠. 植物光合作用的光抑制. 植物生理學通訊, 1992, 28(4): 237-243.
[22] Chen J, Li C Y. Sex-specific responses to environmental stresses and sexual competition of dioecious plants. Chinese Journal of Applied and Environmental Biology, 2014, 20(4): 743-750. 陳娟, 李春陽. 環境脅迫下雌雄異株植物的性別響應差異及競爭關系. 應用與環境生物學報, 2014, 20(4): 743-750.
[23] He M, Meng M, Shi D W,etal. On gender difference of dioecious plant in response to drought stress. Journal of Plant Resources and Environment, 2015, 24(1): 99-106. 何梅, 孟明, 施大偉, 等.雌雄異株植物對干旱脅迫響應的性別差異.植物資源與環境學報, 2015, 24(1): 99-106.
[24] Xu X, Li Y, Wang B,etal. Salt stress induced sex-related spatial heterogeneity of gas exchange rates over the leaf surface inPopuluscathayanaRehd. Acta Physiologiae Plantarum, 2015, 37(1): 1-10.
[25] Chen L, Han Y, Jiang H,etal. Nitrogen nutrient status induces sexual differences in responses to cadmium inPopulusyunnanensis. Journal of Experimental Botany, 2011, 62(14): 5037-5050.
Effects of temperature on flower bud differentiation, pigment contents, and photosynthesis of male and femaleHumulusscandens
GUO Hai-Yan1, DUAN Jing1, LIU Jin-Ping1*, YOU Ming-Hong2, XIE Rui-Juan1
1.ChinaWestNormalUniversityandKeyLaboratoryofEducationonSouthwestChinaWildlifeResourecesConservation,Nanchong637009,China; 2.AcademyofSichuanGrasslandScience,Chengdu611731,China
In this study, we evaluated the effects of temperature on the physiology of male and female plants of the dioecious climbing herb,Humulusscandens. Seeds ofH.scandenswere sown in pots, and the pots were moved to artificial climate incubators set at 15, 20, or 25 ℃ when the seedlings had one to two true leaves. These temperatures were selected based on the average temperatures of the areas where the wild resource are concentrated. We determined the duration of vegetative growth, growth accumulated temperature, number and characteristics of leaves, pigment contents, biomass, and photosynthetic parameters of the male and female seedlings. The effects of temperature on photosynthetic production, net photosynthetic rate (Pn), photosynthetic efficiency, transpiration rate (Tr), intercellular CO2concentration (Ci), and gas exchange (Gs) were also analyzed. The results showed that, 1) temperature and gender had significant effects on growth duration and accumulated temperature (P<0.01). Flower buds differentiated earlier in male plants than in female plants at all three temperatures (P<0.05). Flower bud differentiation was affected by temperature and was optimal at 20 ℃. Growth accumulated temperature differed between males and females, but was not related to growth duration. Temperature significantly affected all of the measured leaf traits (P<0.05), with the strongest effect on leaf number per plant>leaf area per leaf>leaf width>leaf length. The pigment contents did not differ significantly between males and females (P>0.05). Temperature significantly affected the contents of all pigments except for chlorophyll b (P<0.05) with the strongest effect on carotenoids>chlorophyll (a+b)>chlorophyll a/b>chlorophyll a>chlorophyll b. Pigment contents were significantly lower at 20 ℃ than at the other temperatures. Photosynthetic parameters differed significantly between males and females and among temperatures (P<0.05). The temperature affected photosynthetic parameters in the orderPn>Ci>Tr>Gs. The parametersPn,Ci, andTrwere mainly affected by temperature, butGswas mainly affected by gender. The biomass and root biomass distribution were significantly higher in female plants than in male plants (P<0.05). Temperature significantly affected biomass accumulation and allocation in the following order: root>leaf>stem (P<0.01). Biomass per plant significantly decreased as the temperature decreased (P<0.05). The highest biomass allocation to the root and stem was at 25 ℃ and the highest biomass allocation to leaves was at 15 ℃. Key words:Humulusscandens; dioecious; photosynthetic pigment; photosynthesis; biomass
10.11686/cyxb2016425
http://cyxb.lzu.edu.cn
2016-11-14;改回日期:2016-12-15
西華師范大學英才科研基金(17YC337)和四川省科技支撐計劃項目(2011NZ0064)資助。
郭海燕(1993-),女,重慶合川人,在讀碩士.E-mail:1217482599@qq.com*通信作者Corresponding author. E-mail:jpgg2000@163.com
郭海燕, 段婧, 劉金平, 游明鴻, 謝瑞娟. 溫度對雌雄葎草花芽分化和色素含量及光合作用影響的性別差異. 草業學報, 2017, 26(8): 104-112.
GUO Hai-Yan, DUAN Jing, LIU Jin-Ping, YOU Ming-Hong, XIE Rui-Juan. Effects of temperature on flower bud differentiation, pigment contents, and photosynthesis of male and femaleHumulusscandens. Acta Prataculturae Sinica, 2017, 26(8): 104-112.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
