煙臺牟平海洋牧場夏季大型底棲動物群落特征
2017-09-03 08:37:46周政權張建設劉甜甜李曉靜艾冰花李寶泉陳琳琳
海洋科學 2017年5期
關鍵詞:物種
楊 東, 周政權, 張建設, 劉甜甜, 李曉靜, 艾冰花, 李寶泉, 陳琳琳
?
煙臺牟平海洋牧場夏季大型底棲動物群落特征
楊 東1, 周政權2, 3, 張建設1, 劉甜甜1, 李曉靜2, 3, 艾冰花3, 李寶泉3, 陳琳琳3
(1. 浙江海洋大學國家海洋設施養殖工程技術研究中心, 浙江舟山 316000; 2. 中國科學院大學, 北京 100049; 3. 中國科學院煙臺海岸帶研究所, 山東煙臺 264003)
為明確煙臺牟平海洋牧場大型底棲動物群落特征、影響因子及低氧對該區域底棲動物群落特征的影響, 于2015年6月和8月分兩個航次在該區域分別設置相同的18個采樣點進行大型底棲動物群落特征調查和分析。對采集的生物樣品進行群落特征分析, 使用生物統計軟件PRIMER計算優勢度指數()、Shannon-Wiener多樣性指數()、Margalef豐富度指數()和Pielou均勻度指數(), 并進行等級聚類(CLUSTER)和非度量多維標度法(MDS)分析以及azti’s marine biotic index (AMBI)和 multivariate-AMBI (m-AMBI)分析。其中6月航次共采集和鑒定大型底棲動物97種, 優勢類群為多毛類。總平均生物量達到10.64 g/m2, 甲殼類貢獻率最高; 8月航次共采集和鑒定大型底棲動物78種, 多毛類是優勢類群。總平均生物量達到9.28 g/m2, 多毛類貢獻率最高。6月份的,,分別為3.071±0.773、2.782± 0.778和0.622±0.143; 8月份分別為3.192±0.369、2.730±0.510和0.751±0.076。通過CLUSTER聚類和MDS標序發現, 6月份和8月份底棲動物物種組成之間存在顯著差異。環境因子與大型底棲動物豐度和生物量關系較大, 其中最能解釋該群落豐度和生物量的環境因子為水深、水溫、濁度, 而溶解氧是造成6月份與8月份豐度和生物量較大差異的主要因素。AMBI和m-AMBI分析表明該區域部分采樣點大型底棲動物物種組成以污染耐受種為主, 底棲生態健康不佳。煙臺牟平海洋牧場8月份季節性的缺氧環境, 已在短期內影響了底棲動物的物種組成。
煙臺牟平海洋牧場; 大型底棲動物; 低氧; 群落結構; 環境因子
海洋牧場是一種先進的基于生態學原理和現代海洋工程技術的養殖模式, 該模式充分利用原有海區的自然生產力, 在滿足特定環境和生態條件的海域進行科學培育和管理漁業資源而形成的人工漁場。與傳統意義上的海洋捕撈、海水養殖及人工放流不同, 海洋牧場是以環境和生態和諧為目的導向, 集環境保護、資源養護、高效生產以及休閑漁業為一體的海洋經濟新業態, 是解決我國近海環境保護和漁業資源可持續利用的重要途徑[1]。1965年, 曾呈奎先生原創性地提出建設“牧場”的理念, 在海洋中人工控制種植或養殖海洋生物[2]。1977年日本曾建成世界上第一個海洋牧場[3], 之后海洋牧場的建設迅速在世界各國開展, 極大地促進了海洋漁業可持續發展和海洋環境保護。然而, 海洋牧場因對周圍環境極其敏感, 被污染的水體及其他不良的生態環境對其發展存在致命影響[4]。
煙臺牟平海洋牧場地處煙臺市牟平區姜格莊鎮云溪村。云溪海洋牧場面積12 km2, 人工魚礁投放0.58 km2, 總投資1.6億元, 主要增養殖刺參、梭子蟹、魚類等海產品。該海洋牧場所在海域平均年產海參苗種2.5萬kg, 商品參400 t。作為重要的海水養殖基地, 該海域由于養殖密度超過近海生態系統的環境容量, 加之由養殖帶來的污染和多條河流帶來的陸源污染較為嚴重, 致使近岸水域環境污染逐漸加劇。我們開展的現狀調查觀測發現, 2013—2014年夏季, 海洋牧場所在海域曾連續出現底層缺氧現象。
海洋中的溶解氧是海水重要的生源參數, 也是海洋生態系統得以維持發展的關鍵因子[5]。通常情況下, 當溶解氧含量低于4 mg/L時, 養殖魚類就會受到有害影響; 而當溶解氧含量低于2 mg/L, 即出現低氧事件[6]。當低氧事件長時間持續時, 海洋生態系統中的各種動物, 特別是運動能力較弱的底棲動物群落就會受到嚴重損害, 對海洋牧場的生產力和底棲動物的生態環境產生危害[6]。
本文的目的是通過現狀調查并結合歷史資料, 分析煙臺牟平海洋牧場大型底棲動物群落特征及其差異性, 計算該區域底棲生態健康狀況, 掌握低氧對底棲動物群落結構的影響。以期明確牟平海洋牧場大型底棲動物群落特征以及低氧后群落特征的變化, 并開展基于底棲動物群落生物指數的海洋牧場區域底棲生態健康評價。
1 材料與方法
1.1 站位布設和樣品采集
采樣地點位于煙臺牟平海洋牧場, 在該區域海洋牧場共設置18個站位(圖1), 分別于2015年6月與8月進行兩個航次的調查。采用0.05 m2箱式采泥器定量采泥, 每個站位成功采樣3次, 使用0.5 mm孔徑的網篩分選后用75%的乙醇現場固定, 帶回實驗室進行種類鑒定、個體計數和稱量質量。樣品的處理、保存、稱量質量均按照《海洋調查規范》(GB/T 12763.6-2007)進行。研究站位底層水的溫度()、鹽度()和散射濁度(Turb)為CTD現場測定, 溶解氧(DO)為現場采用電化學探頭法測定[7]。
1.2 群落多樣性分析
采用以下指數計算并分析大型底棲動物群落優勢度和多樣性: 優勢度指數(), Shannon-Wiener多樣性指數(), Margalef物種豐富度指數(), Pielou物種均勻度指數()。
根據陳亞瞿等[8]的優勢度標準, 當>0.02時, 即該種為優勢種。
1.3 數據分析方法
使用生物統計軟件PRIMER 7.0進行群落結構的分析。為了平衡優勢種和稀有種在群落中的影響, 數據需經平方根轉換后計算Bray-Curtis相似性系數, 構建相似性矩陣[9]。采用等級聚類(CLUSTER)和非度量多維標度法(MDS)分析群落結構的異質性。通過相似性百分比(SIMPER)方法計算分析不同物種對組內相似性以及組間差異性的平均貢獻率。利用生物環境(BIOENV)相關性分析環境因子與底棲動物群落結構的相關系數; 利用SPSS 18.0軟件方差 (ANOVA)分析6月份與8月份環境因子的差異; 使用azti’s marine biotic index (AMBI)和 multivariate-AMBI (m-AMBI)方法分析大型底棲動物群落受干擾狀況。
AMBI和M-AMBI值均用AMBI v5.0軟件進行計算, 去除非大型底棲無脊椎動物和個體數低于3個的種類, 其余物種的生態分組按照AZTI 中心網站(http: // www. azti. es)公布的最新種類名錄數據(2014年11月)進行, 對于未包含的物種均歸為“未分組類(not assigned)” (Angel Borja等, 2005)。各生態分組中, EG I表示干擾敏感種(disturbance-sensitive species), EG Ⅱ表示干擾不敏感種(disturbance-indifferent species), EG Ⅲ表示干擾耐受種(disturbance-tolerant species), EG Ⅳ表示二階機會種(the second-order opportunistic species), EG Ⅴ表示一階機會種(the first-order opportunistic species)。
2 結果
2.1 海洋牧場環境因子現狀
煙臺牟平海洋牧場6月份與8月份兩次調查的水體環境因子如表1所示。兩個月份各站位鹽度相差不大, 水深、水溫和散射濁度各站位相差較大; 8月份水深和水溫較6月份明顯上升。8月份所有調查站位水體中溶解氧含量明顯低于6月份。
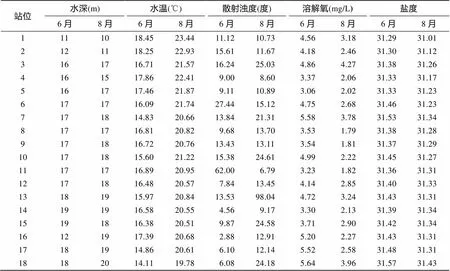
表1 煙臺牟平海洋牧場環境因子數據
2.2 物種組成與優勢種
2015年6月航次共發現和鑒定大型底棲動物97種, 其中優勢類群為多毛類44種, 占總物種數的45.36%; 其次是甲殼類29種, 占總物種數的29.90%; 軟體動物共19種, 占總物種數的19.59%; 棘皮類3種, 占總物種數的3.09%; 其他動物2種, 占總物種數的2.06%。優勢種組成包括短葉索沙蠶()不倒翁蟲微小海螂()尖葉長手沙蠶()、深鉤毛蟲(), 絲異須蟲()。其分別為0.24、0.12、0.09、0.03、0.02、0.02。
2015年8月航次共發現和鑒定大型底棲動物78種, 其中優勢類群為多毛類40種, 占總物種數的51.28%; 其次是甲殼類24種, 占總物種數的30.77%; 軟體動物10種, 占總物種數的12.82%; 棘皮類2種, 占總物種數的2.56%; 其他動物2種, 占總物種數的2.56%。優勢種組成與6月份航次差別不大, 其中有5種重復, 即: 不倒翁蟲、短葉索沙蠶、絲異須蟲、深鉤毛蟲、尖葉長手沙蠶; 兩種為新出現的優勢種, 即: 寡鰓齒吻沙蠶()、內肋蛤()。其分別為0.24、0.13、0.12、0.06、0.06、0.04、0.03。
2.3 生物量和豐度
2015年6月航次底棲動物總平均生物量為10.64 g/m2。其中甲殼類最高, 為4.66 g/m2(占總平均生物量的43.78%); 其后依次為多毛類(3.61 g/m2, 占33.98%)、軟體動物(1.53 g/m2, 占14.37%)、棘皮類(0.68 g/m2, 占6.39%)和其他動物(0.15 g/m2, 占1.46%); 2015年8月航次所調查的底棲動物總平均生物量低于6月份, 為9.28 g/m2。各主要類群所占比例也發生變化, 其中多毛類最高, 為3.00 g/m2(占總平均生物量的32.35%); 其后依次為甲殼類(2.69 g/m2, 占29.06%)、軟體動物(2.19 g/m2, 占23.59%)、其他動物(0.97 g/m2, 占10.48%)和棘皮類(0.42 g/m2, 占4.50%)。其中6月和8月生物量在各采樣點的空間分布不均, 6月高值區位于12號站, 達到89.56 g/m2, 且大大高于6月所有站位生物量平均值; 8月高值區位于6號站, 達到22.37 g/m2(圖2)。
2015年6月航次底棲動物總平均豐度1 084.44 個/m2。其中多毛類最高, 為809.25 個/m2, 占總平均豐度的74.62%; 其后依次是軟體動物(183.33 個/m2, 占16.90%)、甲殼類(82.59 個/m2, 占7.61%)、其他動物 (5.55 個/m2, 占0.51%)、棘皮類(3.70 個/m2, 占0.34%); 2015年8月調查的底棲動物總平均豐度數值明顯較低至154.81 個/m2。其中軟體動物最高, 為90.37 個/m2, 占總平均豐度的58.37%; 其后依次是甲殼類(60.37 個/m2, 占38.99%)、多毛類(1.48 個/m2, 占0.96%)、棘皮類 (1.48 個/m2, 占0.96%)、其他動物(1.11 個/m2, 占0.72%)。其中, 豐度在各采樣點的分布不均, 6月高值區位于5號站, 達到1 806.67個/m2; 8月高值區位于17號站, 達到2 786.67 個/m2, 大大高于8月所有站位豐度平均值(圖3)。
2.4 群落多樣性
2015年6月份航次調查各站位的平均為2.78 ± 0.78, 12號站最高(3.99), 5號站最低(1.60);平均為0.62 ± 0.14, 18號站最高(0.86), 1號站最低(0.27);平均為3.07 ± 0.77, 12號站最高(4.0), 1號站最低(1.01)。
2015年8月份航次調查各站位的平均為2.73 ± 0.51, 16號站最高(3.89), 2號站最低(1.80);平均為0.75 ± 0.08, 12號站最高(0.85), 12號站最低(0.57);平均為3.19 ± 0.37, 13號站最高(3.80), 8號站最低(2.62)。
2.5 群落結構分析
將6月份和8月份的大型底棲動物豐度合在一起, 對其進行聚類分析和非參數性多維標度排序, CLUSTER聚類(圖4a)和MDS排序(圖4b)的分析結果一致。CLUSTER顯示6月份和8月份底棲動物群落物種組成可明顯區分開, MDS也可看到兩個月份的站位也有明顯不同的標序。按照群落物種組成的相似性程度, 6月與8月均可歸為兩組。6月份兩組的平均相似性為38.79%, 貢獻率大于10%的有3種, 以貢獻率大小依次為短葉索沙蠶19.29%, 微小海螂14.60%(表2)。8月份兩組的平均相似性52.36%, 貢獻率大于10%的有4種, 其中貢獻率最大的是不倒翁蟲, 貢獻率為15.46%, 其次是短葉索沙蠶, 貢獻率為14.06%(表2)。6月與8月兩組的平均差異性61.89%, 貢獻率最大的是微小海螂, 貢獻率為5.71%, 是絲異須蟲, 貢獻率為5.66%。
“6-1”表示6月份的1號站位, 以此類推
“6-1” represents for the first sampling station in June, the others are likewise
2.6 群落與環境因子的相關性
通過BIOENV分析得出, 6月份的底棲動物物種豐度與水深、水溫、濁度3個環境因子有很大關系, 相關系數為0.619; 8月份的豐度受環境因子影響最大的因子是濁度, 相關系數為0.374。

表 2 煙臺牟平海洋牧場6月份與8月份物種貢獻率
水深、水溫和濁度3個環境因子的組合與豐度、生物量的相關系數最高, 最能解釋群落豐度和生物量的空間分布特征。
2.7 AMBI與m-AMBI分析
AMBI與m-AMBI分析表明(表3), 6月份煙臺牟平海洋牧場兩個站點6, 11號站底棲動物狀態受到中度干擾, 1、2、5、6、11和17號站底棲群落健康狀況表現為中等; 8月份11號站受到了中度干擾, 11號站底棲群落健康狀況呈中等。

表3 煙臺牟平海洋牧場6月份與8月份各站點AMBI和m-AMBI結果

續表
注: Ⅰ, Ⅱ, Ⅲ, Ⅳ和Ⅴ表示生態類群, 6-1表示6月份的1號站位, 以此類推; 8-1表示8月份的1號站位, 以此類推
3 討論
3.1 煙臺牟平海洋牧場群落結構特征
調查表明, 海域兩個月份底棲動物物種存在較大差異, 8月份物種數明顯少于6月份, 物種組成也主要以多毛類和小型甲殼類為主, 棘皮類和其他類群生物較少, 優勢種均以耐污種類(如短葉索沙蠶和不倒翁蟲等)為主, 說明該海域底棲生物群落結構簡單且具有明顯小型化特征, 同時人類活動引起的沉積層有機質污染和低氧等現象影響到群落組成和分布特征。此外, 調查表明, 海域中未發現刺參的捕食者類群, 如魚類和梭子蟹等, 但其食物來源(如小型甲殼類等)的豐度及生物量水平較低, 也說明該牧場內食物網簡單, 食物鏈長度縮短。調查結果與煙臺近海近幾年的調查結果相差不大, 近些年來煙臺近海大型底棲動物群落物種組成相對穩定, 并沒有發生太劇烈變化[9]。
煙臺牟平海洋牧場大型底棲動物群落結構在6月份和8月份存在顯著差異, 且這些差異的主要貢獻物種為微小海螂和絲異須蟲, 可見這兩個優勢物種對煙臺牟平海洋牧場的環境變化反應較敏感, 可作為煙臺牟平海洋牧場環境評價的指示物種。群落特征和環境因子的關系表明, 水深、水溫和濁度3個環境因子與豐度和生物量的相關系數最高[10]。
物種多樣性指數以及AMBI和m-AMBI分析結果表明, 調查區域各站位的大型底棲動物受到的擾動程度不一, 局部區域受污染擾動較大, 特別是近岸的11號站, 物種組成單一, 說明受污染的環境給多毛類等小型耐污種提供了更大的生存機會[9]。耐污指示種小頭蟲()出現在此站, 說明該站沉積物已受到較嚴重的有機質污染。
3.2 影響底棲生物群落特征的因素
許多研究表明, 不同季節影響大型底棲動物群落特征的環境因素存在著較明顯的差異[11]。本次調查研究的結果也發現這一現象。其中影響6月份底棲生物群落特征主要環境因子為水深、水溫和濁度, 8月份則為濁度。煙臺牟平海洋牧場大型底棲動物的物種特征和群落分布格局與水深有著密切的關系。隨著水體深度的不斷增加, 該海洋牧場大型底棲動物的密度和種類數通常會隨之減少。一般而言, 單位面積內深海大型底棲動物多樣性低于淺水區, 而淺海區相對深海具有更高的豐度和生物量。且大型底棲動物種類數、豐度、生物量、多樣性指數和種類均勻度與水深呈負相關關系[12]。而海水的水溫與大型底棲動物種類數, 豐度, 生物量存在著正相關關系。隨著水溫的不斷升高, 大型底棲動物開始進行繁殖, 其個體數和生物量也隨之增加[13]。濁度是通過影響光照強度從而影響該海洋牧場浮游植物和著生藻類的生長, 進而影響以取食浮游植物或著生藻類為食的底棲動物[14]。
6月份與8月份煙臺牟平海洋牧場的含氧量差異顯著, 且8月份含氧量明顯低于6月份含氧量。8月份有3個站出現低氧情況, 分別為8號站1.79 mg/L, 9號站1.81 mg/L, 11號站1.82 mg/L, 且8月份平均含氧量也較低(表1)。6月份3站的生物量分別為15.57、8.19和3.35 g/m2, 而8月份3站的生物量分別為生物量7.45、3.49和2.14 g/m2。8月份處于低氧環境下的3站與6月份相同站位的生物量明顯減少。由此可推論, 含氧量是造成6月份與8月份生物量和豐度差異的主要環境因子。
3.3 低氧的影響
底層水中溶解氧含量是影響底棲生物的主要因素[15]。水體缺氧嚴重危害河口和近海生態系統, 海洋中大多數生物都需要充足的溶解氧來維持生長代謝[16]。因底棲動物大多移動能力弱或移動范圍有限, 區域性的缺氧對底棲動物的影響最為顯著, 當溶解氧質量濃度小于2 mg/L, 且持續時間較長時, 海洋中的大多數魚類、浮游動物, 特別是運動能力較弱的物種將面臨大規模死亡[17]。缺氧事件發生還會降低海洋物種多樣性, 改變海洋生物的群落結構, 減少魚類和底棲動物的豐富度, 從而影響漁業生產, 帶來直接或間接的經濟損失[18]。
低氧環境成因復雜, 低氧發生時會對大型底棲動物產生擾動, 進而使低氧區內外大型底棲動物的棲息密度、生物量和多樣性產生較大差異[19]。許多多毛類物種對環境溶解氧含量的適應性較強, 無論在低氧區還是非低氧區常作為調查海域沉積物中優勢類群[20]。同時, 我們也發現煙臺牟平海洋牧場8月份低氧區的存在并沒有完全破壞底棲生物群落, 只可能在短時間影響底棲動物的種類組成[21]。
[1] 楊紅生, 霍達, 許強, 等. 現代海洋牧場之我見[J]. 海洋與湖沼, 2016, 47(6): 1069-1074. Yang Hongsheng, Huo Da, Xu Qiang, et al. Views on modern marine ranching[J]. Oceanologia et Limnologia Sinica, 2016, 47(6): 1069-1074.
[2] 楊紅生. 我國海洋牧場建設回顧與展望[J]. 水產學報, 2016, 40(7): 1133-1140. Yang Hongsheng. Construction of marine ranching in China: reviews and prospects[J]. Journal of Fisheries of China, 2016, 40(7): 1133-1140.
[3] 楊金龍, 吳曉郁. 海洋牧場技術的研究現狀和發展趨勢[J]. 中國漁業經濟, 2004, 5: 48-50.Yang Jinlong, Wu Xiaoyu. Overview of marine ranching Technology[J]. Chinese Fisheries Economics, 2004, 5: 48-50.
[4] 范純增, 劉玉寶, 姜虹, 等. 山東海洋牧場可持續發展與環境保護[J]. 水產學雜志, 2011, 14(2): 7-11. Fan Chunzeng, Liu Yubao, Jiang Hong, et al. Marine ranch sustainable development and environment protection in Shandong province, China[J]. Chinese Joural of Fisheries, 2011, 14(2): 7-11.
[5] 鄭靜靜, 劉桂梅. 海洋缺氧現象的研究進展[J]. 海洋預報, 2016, 33(4): 88-97. Zheng Jingjing, Liu Guimei. Review on the marine hypoxia[J]. Marine Forecasts, 2016, 33(4): 88-97.
[6] 劉海霞. 長江口夏季低氧區形成及加劇的成因分析[D]. 上海: 華東師范大學河口岸學國家重點實驗室, 2011. Liu Haixia. Study on main influencing factors of formation and deterioration of summer hypoxia off the Yangtze River Estuary[D]. Shanghai: State Key Laboratory of Estuarine and Coastal Research of East China Normal University, 2011.
[7] 王智, 曲方圓. 夏季遼東灣西部海域大型底棲動物群落結構與多樣性[J]. 海洋科學, 2016, 40(1): 40-47. Wang Zhi, Qu Fangyuan. Commructure and diversity of macrobenthos in the western waters of Liaodong Bay during summer[J]. Marine Sciences, 2016, 40(1): 40-47.
[8] 鄧玉娟, 董樹剛, 劉曉收. 福建興化灣大型底棲動物種類組成和生物多樣性[J]. 海洋科學, 2016, 40(1): 54-63. Deng Yujuan, Dong Shugang, Liu Xiaoshou. Species composition and biodiversity of macrofauna in Xinghua Bay, Fujian[J]. Marine Sciences, 2016, 40(1): 54-63.
[9] 李曉靜, 周政權, 陳琳琳, 等. 山東煙臺大沽夾河河口及鄰近海域大型底棲動物群落特征[J]. 生物多樣性, 2016, 24(2): 157-165. Li Xiaojing, Zhou Zhengquan, Chen Linlin, et al. Characteristics of microbenthic communities in the estuary of Dagujia River and its adjacent water areas in Yantai, Shandong[J]. Biodiversity Science, 2016, 24(2): 157-165.
[10] 彭松耀. 黃東海大型底棲動物群落結構特征[D]. 青島: 中國科學院海洋研究所, 2013. Peng Songyao. Characteristics of macrobenthic community structure in the Yellow Sea and East China Sea[D]. Qingdao: Institute of Oceanology, the Chinese Academy of Sciences, 2013.
[11] 李莉, 張建樂. 秦皇島入海口臨近海域夏季大型底棲動物群落結構變化及影響因子研究[J]. 海洋學研究, 2014, 32(4): 83-89. Li Li, Zhang Jianle. Study on the community structural variation of macrobenthos and impact factors in the adjacent sea area of Qinghuandao Estuary in summer [J]. Journal of Marine Sciences, 2014, 32(4): 83-89.
[12] 張敬懷. 珠江口及鄰近海域大型底棲動物多樣性隨鹽度、水深的變化趨勢[J]. 生物多樣性, 2014, 22(3): 302-310. Zhang Jinghuai. The variation of biodiversity of macrobenthic fauna with salinity and water depth near the Pearl Estuary of the northern South China Sea [J]. Biodiversity Science, 2014, 22(3): 302-310.
[13] 陸強, 陳慧麗. 杭州西溪濕地大型底棲動物群落特征及與環境因子的關系[J]. 生態學報, 2013, 33(9): 2803-2815. Lu Qiang, Chen Huili. Ecological characteristics of macrobenthic communities and its relationships with enviromental factors in Hangzhou Xixi Wetland [J]. Acta Ecologica Sinica, 2013, 33(9): 2803-2815.
[14] 邵美玲, 韓新芹. 香溪河流域梯級水庫底棲動物群落比較[J]. 生態學報, 2017, 27(12): 4963-4971. Shao Meiling, Han Xinqin. Comparative on macroinvertebrate communities along a cascade of reservoirs in Xiangxi River Basin[J]. Acta Ecologica Sinica, 2017, 27(12): 4963-4971.
[15] 張曉舉, 丁龍, 馮春暉. 遼東灣中部海域春季大型底棲生物的群落結構及影響因子[J]. 海洋科學, 2016, 40(10): 43-48. Zhang Xiaoju, Ding Long, Feng Chunhui. Macrobenthos community and environmental factors in the middle waters of Liaodong Bay in Spring[J]. Marine Sciences, 2016, 40(10): 43-48.
[16] Gray J S, Wu R S S, Or Y Y. Effects of hypoxia and organic enrichment on the coastal marine environment[J]. Marine Ecology Progress Series, 2002, 238: 249-279.
[17] Karlson K, Rosenberg R, Bonsdorff E, et al. Temporal and spatial large-scale effects of eutrophication and oxygen deficiency on benthic fauna in Scandinavian and Baltic waters-a review[J]. Oceanography and Marine Biology, 2002, 40: 427-489.
[18] Yin K D, Lin Z F, Ke Z Y. Temporal and spatial distribution of dissolved oxygen in the Pearl River Estuary and adjacent coastal waters[J]. Continental Shelf Research, 2004, 24(16): 1935-1948.
[19] 袁偉, 金顯仕. 低氧環境對大型底棲動物的影響[J]. 海洋環境科學, 2010, 29(3): 293-296.Yuan Wei, Jin Xianshi. Influence of hypoxia environment upon macrobenthos[J]. Marine Environmental Science, 2010, 29(3): 293-296.
[20] 劉志國, 許韌. 長江口外低氧區特征及其影響研究[J].海洋通報, 2012, 31(5): 588-593. Liu Zhiguo, Xu Ren. Characters of hypoxia area off the Yangtze River Estuary and its influence[J]. Marine Science Bulletin, 2012, 31(5): 588-593.
[21] 王延明, 李道季. 長江口及臨近海域底棲生物分布及與低氧區的關系研究[J]. 海洋環境科學, 2008, 27(2): 139-144.Wang Yanming, Li Daoji. Study on relation of distribution of benthos and hypoxia in Yangtze River Estuary and adjacent sea [J]. Marine Environment Science, 2008, 27(2): 139-144.
Characteristics of macrobenthic communities at the Muping marine ranch of Yantai in summer
YANG Dong1, ZHOU Zheng-quan2, 3, ZHANG Jian-she1, LIU Tian-tian1, LI Xiao-jing2, 3, AI Bing-hua3, LI Bao-quan3, CHEN Lin-lin3
(1. Zhejiang Ocean University, Zhoushan 316000, China; 2. University of the Chinese Academy of Sciences, Beijing 100049, China; 3. Yantai Institute of Coastal Zone Research, the Chinese Academy of Sciences, Yantai 264003, China)
A survey on the characteristics of macrobenthic communities was carried out at the Muping marine ranch of Yantai during June and August 2015to clarify the characteristics and factors affecting the macrobenthic communities and to analyze the effects of hypoxia on macrobenthic community characteristics. The PRIMER software package was used to calculate four biodiversity indices and to carry out CLUSTER, MDS ordination analyses, AMBI AZTI’s Marine Biotic Index and m-AMBI (multivariate-AMBI) to analyze community structure. A total of 97 macrobenthic species were identified in June, of which the dominant group was polychaetes. The average biomass was 18.02 g/m2, of which crustacean species contributed most. A total of 78 macrobenthic species were identified in August, of which the dominant group was polychaetes. The average biomass was 9.28 g/m2, of which polychaete species contributed most. Three biodiversity indices, the richness index, the evenness index, and the Shannon–Wiener index were 3.071 ± 0.773, 2.782 ± 0.778, and 0.622 ± 0.143 in June and 3.192 ± 0.369, 2.730 ± 0.510, and 0.751 ± 0.076in August. The CLUSTER and MDS analyses showed a significant difference between June and August. Environmental factors, including water depth, water temperature, and turbidity were strongly correlated with the abundance and biomass of the macrobenthic communities. Dissolved oxygen was the main cause of the difference between June and August. The AMBI and M-AMBI analysis showed that the macrobenthic species were mostly tolerant and that the area had suffered a moderate disturbance. The seasonal hypoxic environment at the Muping marine ranch of Yantai in August affected species composition of the macrobenthos.
Muping marine ranch of Yantai; macrobenthos; hypoxia; community structure; environmental factors
(本文編輯: 劉珊珊)
[Key Program of the Chinese Academy of Sciences, No.KZZD- EW-14; Strategic Priority Research Program of the Chinese Academy of Sciences, No.XDA11020403, No.XDA11020702]
Mar. 15, 2017
Q933
A
1000-3096(2017)05-0134-10
10.11759//hykx 20170315002
2017-03-15;
2017-04-07
中國科學院院重點部署項目(KZZD-EW-14); 中國科學院戰略性先導科技專項(XDA11020403, XDA11020702)
楊東(1995-), 男, 安徽蕪湖人, 碩士, 主要從事底棲動物生態學研究, 電話: 18857087817, E-mail: 1581100508@qq.com; 李寶泉, 通信作者, 副研究員, 碩士生導師, 電話: 13695358760, E-mail: bqli@yic.ac.cn
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
