油菜NO3–的吸收、分配及氮利用效率對低氮脅迫的響應
2017-09-03 09:34:49榮楠韓永亮榮湘民宋海星彭建偉謝桂先張玉平張振華
植物營養與肥料學報 2017年4期
榮楠,韓永亮,榮湘民,宋海星,彭建偉,謝桂先,張玉平,張振華
(湖南農業大學南方糧油作物協同創新中心/資源環境學院,長沙410128)
油菜NO3–的吸收、分配及氮利用效率對低氮脅迫的響應
榮楠,韓永亮*,榮湘民,宋海星,彭建偉,謝桂先,張玉平,張振華*
(湖南農業大學南方糧油作物協同創新中心/資源環境學院,長沙410128)
【目的】探究油菜NO3–的吸收、分配和對低氮脅迫的響應及其氮利用效率,為理解油菜在不同低氮脅迫下相關生理變化及其氮素利用效率提供科學依據。【方法】以常規油菜品種814為研究材料,采用砂培試驗,在正常供氮水平(10mmol/L)和低氮脅迫水平(3mmol/L、1mmol/L)下,研究油菜的根系特性、蒸騰作用對低氮脅迫的響應及其氮素吸收效率,并研究油菜NO3–的運輸分配與同化對低氮脅迫的響應及其氮素利用效率。【結果】與正常供氮處理(10mmol/L)相比,低氮脅迫處理(3mmol/L、1mmol/L)的油菜NO3–含量、全氮含量均顯著下降,但(NO3–)葉/根、(全氮(%))葉/根顯著升高,植株根系干物質重、根系吸收面積均顯著下降,但根冠比顯著升高。油菜植株在低氮脅迫下氣孔導度和蒸騰速率顯著增加,一方面促進植株對NO3–的捕獲,另一方面也促使更大比例的NO3–分配在植物的地上部分,但植株的水分散失加劇,水分利用效率顯著下降。低氮脅迫處理油菜根和葉中NR、GS活性與正常供氮處理之間的差異不顯著或有增加,其葉綠素含量、光合速率均顯著下降,但光合氮素利用率顯著升高。【結論】在低氮脅迫條件下,油菜植株的氮素和干物質累積均顯著下降,但NO3–在植株的地上部分分配比例的增加以及光合氮素利用率的升高促使植株的氮素利用效率顯著提高。
油菜;NO3–;氮素吸收效率;氮素利用效率
在低氮環境中,植物發生一系列形態和生理上的變化來適應外界環境,如根長變長、側根密度降低等[1–2],同時有較多的光合產物被根系利用,促進根系的生長,從而提高根冠比。曹蘭芹等[3]研究表明,與高氮處理相比,油菜在低氮處理下其氮素吸收效率升高,但其機理尚不夠明確。根系是油菜吸收養分的主要器官,其活力水平和根表面積對氮的吸收極為重要,氮吸收能力強的植物根系活力和吸收面積往往較大。NO3–是油菜的主要氮源,土壤中的NO3–主要通過質流遷移至根系表面,而質流的動力來源于植物的蒸騰作用,因此,油菜的蒸騰速率對NO–的吸收具有重要促進作用。油菜吸收NO–33后,一部分在根系中同化利用,另一部分通過木質部運輸到植物的地上部參與同化反應,因此,木質部傷流液中NO3–含量可反映其體內分配對不同氮水平的適應性。硝酸還原酶(NR)和谷氨酰胺合成酶(GS)是NO3–同化的兩個關鍵酶。NR是NO3–誘導酶,是其參與同化的第一個步驟,即在NR作用下,NO3–在細胞中還原成NO2–。生成的NO2–在亞硝酸還原酶的作用下還原成氨,參與下一步的同化反應。由于亞硝酸還原酶的活性遠遠高于硝酸還原酶,所以植物體內幾乎沒有NO2–的積累,故NR是NO3–還原的限速酶[4]。NO3–被還原生成的NH4+,在GS酶的作用下,合成谷氨酰胺,參與體內的各種新陳代謝反應。NR和GS對植株的氮效率有顯著促進作用,氮效率較高的油菜植株其NR和GS活性往往也較高[5–6]。植物體內氮代謝與碳代謝相互偶聯,NO3–的同化過程利用由光合作用提供的還原劑、能量和碳骨架,因此,植物的光合作用對NO3–的同化利用具有重要促進作用[7–8]。
油菜是我國主要油料作物,其氮肥利用率較低[9],在農業生產上氮肥施用過量和不足并存[10–12]。為了精確控制油菜生長的營養環境,本研究以水和稀鹽酸清洗干凈的珍珠巖砂粒作為油菜的生長基質,開展砂培試驗,以正常供氮水平為對照,設置中度和重度低氮脅迫處理,在油菜移栽后第40d采樣(該時期油菜對低氮脅迫的適應性生理變化趨于穩定),測定油菜NO3–吸收、分配和同化的相關生理指標,對研究油菜在不同低氮脅迫下相關生理變化及其氮素利用效率,對生產實踐具有重要指導意義。
1 材料與方法
1.1 試驗設計
供試油菜品種為814(常規油菜品種),由國家油料作物改良研究中心湖南分中心提供。
根據氮水平的不同,該試驗設3個處理:1)正常供氮處理(NO3–10mmol/L);2)中度低氮脅迫處理(NO3
–3mmol/L);3)重度低氮脅迫處理(NO3–1 mmol/L)。每處理重復3次。
油菜播種出苗后,長至第4片葉展開后進行移栽,油菜移栽至塑料缽中(30cm×30cm),每缽一株。塑料缽中盛放用水和稀鹽酸清洗干凈的珍珠巖砂粒(無營養液成分)作為油菜的生長基質。移栽油菜后的塑料缽放置在玻璃大棚中,油菜在自然光照和自然溫度下生長,在油菜移栽后的生長期內,每3d澆一次營養液,一次240mL。正常供氮處理(即10 mmol/L處理)的營養液成分:KNO33.3mmol/L、Ca(NO3)2·4H2O3.3mmol/L、Fe-EDTA1mmol/L、KH2PO41mmol/L、MgSO45mmol/L、B0.5mg/L、Mn0.5mg/L、Zn0.05mg/L、Cu0.02mg/L、Mo0.01 mg/L,使用HCl和NaOH調節pH至5.8;處理2)和處理3)的營養液成分中除KNO3和Ca(NO3)2·4H2O的用量減少為NO3–濃度為3mmol/L和1mmol/L外,其他同正常供氮處理,減量的鉀和鈣以KCl和CaCl2的形式補充。油菜生長的水分全部來源于所澆營養液。
1.2 測定方法
在油菜的苗期(移栽后40d)分植株的地上部(從下往上第4片葉)和地下部取樣。分別測定全氮和NO3
–含量,酶活及干物質量等。并測定植株地下部分的根系活力、根系吸收面積,地上部分的氣孔導度、蒸騰速率、光合速率。
樣品經研磨浸提后,采用連續流動分析儀(AA3)測定NO3
–含量;NR活性采用Fan等[14]方法測定;GS活性采用楊蘭[15]所用測定方法測定;植株全氮采用凱氏定氮法測定[16];根系活力采用TTC法測定[13];根系吸收面積采用甲烯藍法測定[13]。
氣孔導度、蒸騰速率、光合速率采用LI-6400便攜式光合儀測定。測定參數設置如下:光照為1000 μmol/(m2·s),相對濕度為65%,CO2濃度為400 mg/kg,溫度為環境溫度。
木質部汁液的收集及其NO3–含量的測定:用剪刀將油菜植株從根莖上1cm處剪斷,其根立即轉移至營養液中,用預稱重的脫脂棉球附在油菜的切割面上并用塑料薄膜包裹,1h后用注射器擠壓棉花,收集棉花的滲出液,即木質部汁液。根據棉花的增重來計算木質部汁液的體積。在連續流動分析儀(AA3)上測定滲出液NO3–濃度。
1.3 數據計算及處理
氮素光合效率=光合速率/葉片單位面積氮素量;
水分利用效率(WUE)=光合速率/蒸騰速率[17];
氮素吸收效率=植株總氮量/營養液提供的總氮量;
氮素利用效率=生物產量/植株總氮量。
采用SPSS13.0和Microsoft Excel軟件進行數據處理。處理間差異顯著性分析采用LSD檢驗法。
2 結果與分析
2.1 油菜NO3–、全氮含量及體內分布對低氮脅迫的響應
與正常供氮處理(10mmol/L)相比,在低氮脅迫下(3mmol/L、1mmol/L)油菜根和葉中NO3–含量均顯著下降,3個氮處理之間的差異均達到了極顯著水平(圖1)。但(NO3–含量)葉/根在低氮脅迫下升高,3mmol/L比10mmol/L處理高25.99%,兩者之間的差異達到了顯著水平,1mmol/L比3mmol/L處理高40.06%,兩者之間的差異達到了極顯著水平。
油菜植株根和葉的全氮含量均隨供氮水平的降低而降低。[植株全氮(%)]葉/根在低氮脅迫下有升高趨勢。3mmol/L與10mmol/L處理之間的差異不顯著,1mmol/L比10mmol/L處理高16.88%,兩者之間的差異達極顯著水平,1mmol/和3mmol/L處理之間的差異不顯著。
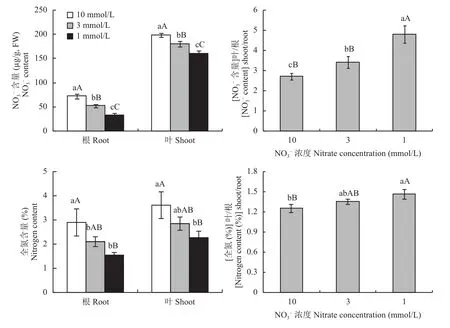
圖1 不同氮濃度水平下油菜NO3–、全氮含量及其體內分布Fig. 1 Nitrate nitrogen content, total nitrogen and their distribution of oilseed rape in different nitrate concentration levels
2.2 油菜根系特性對低氮脅迫的響應
油菜植株的根干物質重隨供氮水平的降低而降低(圖2),3mmol/L比10mmol/L處理顯著降低36.91%,1mmol/L與3mmol/L之間的差異不顯著。油菜植株的根系吸收面積隨供氮水平的降低而減少,3個氮處理之間差異均達到了極顯著水平。10 mmol/L和3mmol/L處理根系活力差異不顯著,但均顯著高于1mmol/L處理,分別高出27.69%和41.25%。油菜植株的根冠比隨著供氮水平的降低而升高,3mmol/L的根冠比比10mmol/L處理高26.31%,兩者之間的差異達到了顯著水平,1mmol/L的根冠比比3mmol/L高28.23%,兩者之間的差異達到了極顯著水平,這可能是油菜植株的氮素吸收效率隨供氮水平降低而升高的原因之一。
2.3 油菜氣孔導度、蒸騰速率對低氮脅迫的響應
由圖3可知,油菜植株的氣孔導度、蒸騰速率在低氮脅迫下均顯著上升。3個氮處理之間的差異均達到了極顯著水平,3mmol/L和1mmol/L處理的蒸騰速率比10mmol/L處理分別高出12.30%和22.25%。
2.4 油菜木質部汁液對低氮脅迫的響應
由表1可以看出,3mmol/L處理的木質部汁液體積極顯著高于10mmol/L和1mmol/L處理,10 mmol/L和1mmol/L處理之間的差異不顯著。3 mmol/L處理的木質部汁液中NO3–濃度最低,與10 mmol/L處理之間的差異達到了極顯著水平,與1 mmol/L處理之間的差異達到了顯著水平。10mmol/L處理收集到的木質部汁液中NO3–的總量顯著高于3 mmol/L、1mmol/L處理,10mmol/L處理分別比3 mmol/L、1mmol/L處理高34.35%、64.98%,而3 mmol/L和1mmol/L處理間差異不顯著。
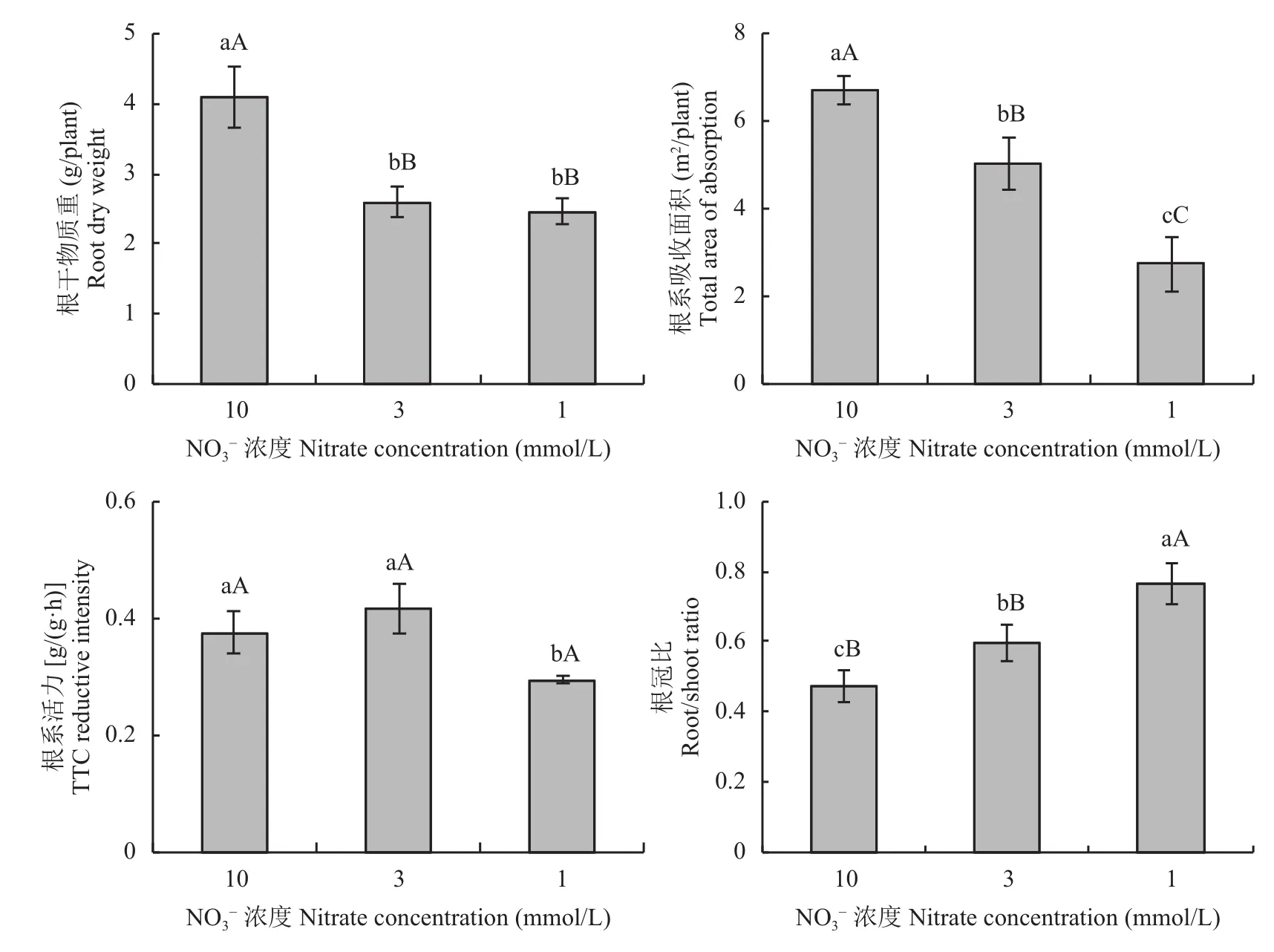
圖2 不同氮濃度水平下油菜的根系特性Fig. 2 Root characteristics of oilseed rape in different nitrate concentration levels
2.5 油菜NO3–代謝關鍵酶活性對低氮脅迫的響應
由圖4可知,10mmol/L和3mmol/L處理的油菜根系中NR活性差異不顯著,但均極顯著低于1mmol/L處理。在葉中,10mmol/L處理的NR活性與3mmol/L、1mmol/L處理之間的差異均不顯著。3 mmol/L比1mmol/L處理低38.39%,兩者之間的差異達到了顯著水平。
在根中,3個氮處理之間的GS活性差異均不顯著;在葉中,10mmol/L和3mmol/L處理之間的差異不顯著,但均極顯著低于1mmol/L處理。
2.6 油菜光合速率、光合氮素利用率及水分利用率對低氮脅迫的響應
由表2可以看出,植株的葉綠素含量和光合速率在低氮脅迫下均顯著下降,但3mmol/L與1 mmol/L處理之間的差異不顯著。在低氮脅迫下,植株的光合氮素利用率顯著升高,3mmol/L比10 mmol/L處理高19.03%,兩者之間的差異達顯著水平,1mmol/L比10mmol/L處理高38.11%,兩者之間的差異達極顯著水平,1mmol/L與3mmol/L處理之間的差異達顯著水平。
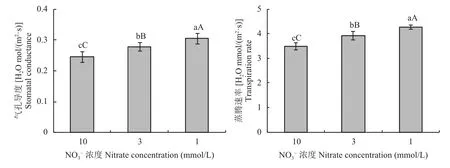
圖3 不同氮濃度水平下油菜氣孔導度、蒸騰速率Fig. 3 Stomatal conductance, transpiration of oilseed rape in different nitrate concentration levels
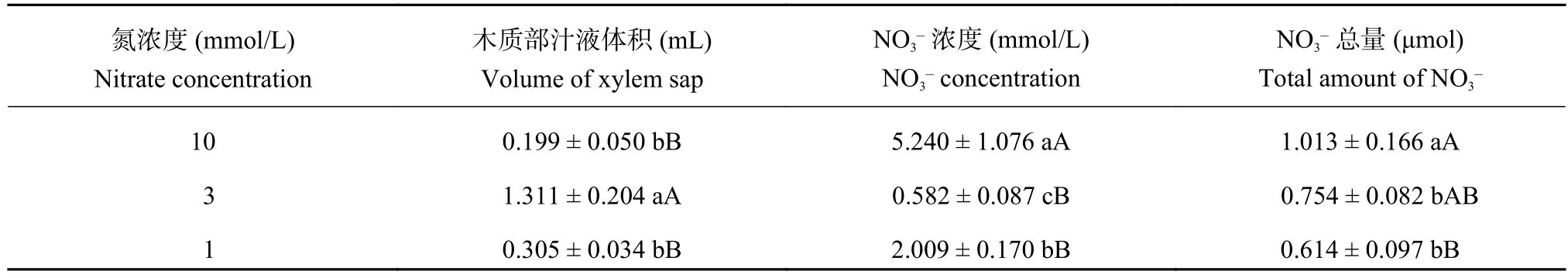
表1 不同氮濃度水平下油菜木質部汁液體積及NO3–濃度Table 1 Volume of xylemsap, NO3–concentration of oilseed rape in different nitrate concentration levels

圖4 不同氮濃度水平下油菜NR、GS活性Fig. 4 NR, GS activities of oilseed rape in different concentration nitrate levels
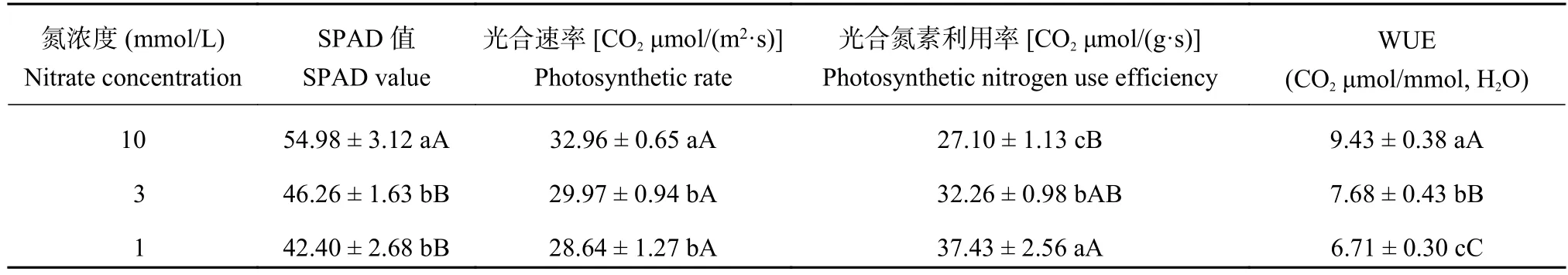
表2 油菜光合速率、光合氮素利用率及水分利用率對低氮脅迫的響應Table 2 Photosynthetic nitrogen use efficiency and water use efficiency (WUE) of oilseed rape in different nitrate concentration levels
油菜植株的水分利用效率在低氮脅迫下顯著下降,3mmol/L比10mmol/L處理降低18.58%,1 mmol/L比10mmol/L處理降低28.86%,3個氮處理之間的差異均達到了極顯著水平。
2.7 油菜氮素與干物質累積以及氮素吸收利用效率對低氮脅迫的響應
由表3可知,油菜植株的總氮、地上部生物量及總生物量均在低氮脅迫下顯著下降。3mmol/L、1 mmol/L處理的總氮和總生物量與10mmol/L之間的差異均達到了極顯著水平,3mmol/L與1mmol/L處理之間的差異達到了顯著水平。3個氮處理地上部生物量之間的差異均達到了極顯著水平。3mmol/L處理的總氮比10mmol/L的降低了60.42%,其地上部生物量比10mmol/L降低了49.61%,其植株總生物量比10mmol/L降低了47.73%。1mmol/L處理的總氮比10mmol/L的降低了75.86%,其地上部生物量降低了62.79%,其植株總生物量降低了57.89%。
植株的氮素吸收效率和氮素利用效率在低氮脅迫下顯著升高,3個氮處理之間的差異均達到了極顯著水平。3mmol/L處理的氮素吸收效率比10mmol/L處理的提高了31.66%,其氮素利用效率提高了31.71%。1mmol/L的氮素吸收效率比10mmol/L處理提高了141.20%,其氮素利用效率提高了73.73%。
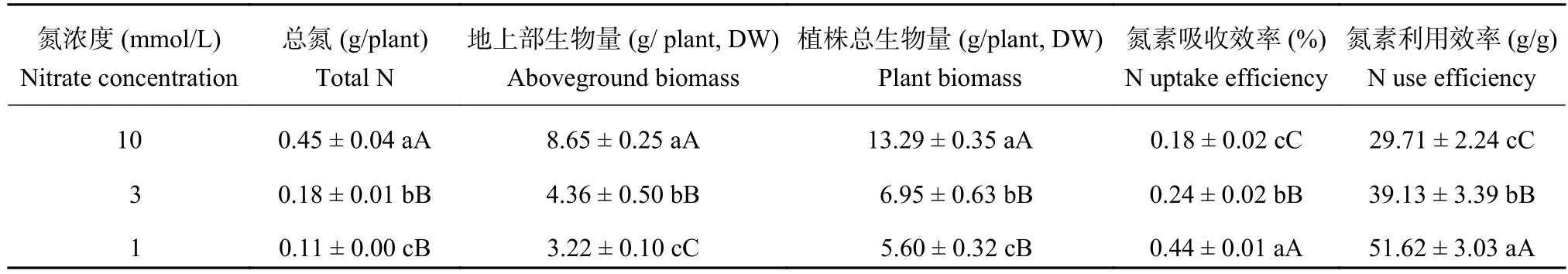
表3 油菜氮素與干物質累積以及氮素吸收利用效率對低氮脅迫的響應Table 3 Nitrogen and dry matter accumulation, nitrogen uptake and utilize efficiency of oilseed rape in different nitrate concentration levels
3 討論
3.1 根系特性與蒸騰作用對低氮脅迫的響應及其對氮素吸收效率的影響
根是植物吸收NO3–的主要器官,氮素吸收效率高的植物一般都具有根系發達、生物量大、分布密度大等特點。為了更好地吸取養分,根系在不同的生長環境中,進化出一系列形態或生理上的塑性反應。在低氮環境中,植物體通過減少側根密度,增加根長來減少根系間的競爭和增加根系在土壤中的生長范圍[1–2],從而提高NO3–吸收效率。在本試驗中,中度低氮脅迫處理(3mmol/L)和重度低氮脅迫處理(1mmol/L)的油菜植株由于總氮吸收量減少(表3),蛋白質合成量降低,造成植株生長矮小,生物量下降,但根冠比顯著增加(圖2)。1mmol/L與3 mmol/L的根干物質重差異不顯著,但根冠比差異達極顯著水平,這是由于1mmol/L的地上部生物量極顯著低于3mmol/L所致(表3)。作物通過增加生物量在根系中的分配來促進植物體對養分離子的吸收,提高其氮素吸收效率(表3),這可能是植物對低氮環境的適應機制之一。在低氮脅迫下,植物地上部合成的碳水化合物向根系運輸加強,根系中出現淀粉累積現象[18–19],根系的生長優于地上部的生長,這可能是導致植物體根冠比變大的主要原因。
本試驗表明,油菜在低氮脅迫下,不僅通過改變植物的根系特性來適應低氮環境,而且能夠改變植物地上部分的生理狀況來加強對氮素的吸收。研究表明,土壤中的氮素能夠調控植物的蒸騰作用[17],在缺氮環境中,植物體的氣孔導度、蒸騰速率提高,土壤中的質流加強[17,20–21]。這與本試驗的研究結果一致,低氮脅迫下油菜植株的氣孔導度、蒸騰速率顯著升高(圖3)。質流雖然不能直接促進植物體對NO3
–的吸收,但能夠提高根際NO3–濃度,因此,蒸騰作用對于植物體對NO3–的捕獲具有重要促進作用。在低氮脅迫下,油菜植株通過提高蒸騰作用來加強對NO3
–的吸收,是其適應低氮環境的機制。但在低氮脅迫下,升高的蒸騰作用加劇了植物體水分的散失,導致其水分利用效率(WUE)下降(表2)。因此,油菜植株在低氮脅迫下通過犧牲其水分利用效率來促使其氮素吸收效率的提高。
3.2 NO3
–的運輸分配與同化對低氮脅迫的響應及氮素利用效率NO3
–被植物體吸收后,既可以在根系中同化利用,也可以轉移到根系液泡中儲存起來,被吸收的硝態氮還可以通過木質部運往植物的地上部分,在植物的地上部分同化利用。硝態氮在植物的地上部分與地下部分分配的比例,取決于植物品種、外部NO3
–濃度、溫度、光照強度等[22]。NO3–在植物體內的分配對氮素利用效率有重要影響,研究表明,油菜體內NO3
–被更多的分配在地上部分,能夠促進其氮效率的提高[23]。NO3–在葉片中同化能更有效的利用由光合作用提供的還原劑、能量和碳骨架,因此,NO3
–的還原發生在植物的地上部分比在地下部分更為節能高效[7–8]。在本試驗中,低氮脅迫處理油菜體內NO3
–含量顯著下降(圖1),但有更高比例的NO3
–被分配在植物的地上部分。且低氮脅迫得越重,向地上部分分配NO3–的比例越高。NR和GS是NO3
–代謝的兩個關鍵酶,對植株NO3–的同化有顯著促進作用。低氮脅迫處理油菜體內NR、GS活性無明顯下降(圖4)。低氮脅迫下的油菜植株葉綠素含量、光合速率降低,但光合氮素利用率顯著增加,且低氮脅迫得越重,光合氮素利用率增加越多(表2)。油菜植株在低氮脅迫下,NO3–在地上部的分配比例增加(圖1),同時伴隨著光合氮素利用率的顯著提高,這是植株氮素利用效率顯著提高的重要原因(表3)。
低氮脅迫處理油菜植株蒸騰速率提高(圖3),不僅能夠促進植物體對NO–的吸收,而且促進了NO–33從根系向地上部分的運輸[24–25]。當供氮水平從10 mmol/L減至3mmol/L時,木質部汁液中的NO3–總量僅減少25.57%,供氮水平從3mmol/L減至1mmol/L時,木質部汁液中的NO3–總量僅減少22.80%(表1)。
氣孔的開合控制著植物的蒸騰速率和植物體對CO2的吸收,氣孔孔徑增加,氣體交換加強,一方面可以促進植物的光合作用,另一方面,也加劇了水分的散失。隨著供氮水平的降低,油菜植株氮含量下降(圖1),葉綠素合成受阻,光合作用受到限制,但蒸騰作用加劇,導致其水分利用效率降低(表2)。氣孔的閉合受保衛細胞中NO3–的調控,當其濃度降低時,氣孔的開合度也隨之降低[26]。當外界氮水平降低時,氣孔的分布和密度不變[27],但油菜植株的氣孔導度增加(圖3)。該研究表明植物的氮代謝對氣孔的閉合具有調控作用,但這一調控機理還需要進一步研究。
4 結論
1)在低氮脅迫下,油菜氮素吸收總量降低,但氮素吸收效率和氮素利用效率均顯著升高。
2)油菜植株的根冠比、蒸騰速率均在低氮脅迫下增加,促進了植株對NO3–的吸收。
3)低氮脅迫下,油菜蒸騰速率的升高促進了NO3
–向植株地上部分的運輸,同時光合氮素利用率顯著升高,促進了植株氮素利用效率的提高,但植株水分散失加劇,水分利用效率顯著下降。
[1]Postma JA,Dathe A,Lynch JP.The optimal lateral root branching density for maize depends on nitrogen and phosphorus availability[J]. Plant Physiology,2014,166(2):590–602.
[2]謝孟林,李強,查麗,等.低氮脅迫對不同耐低氮性玉米品種幼苗根系形態和生理特征的影響[J].中國生態農業學報,2015,23(8): 946–953. Xie ML,Li Q,Zha L,et al.Effects of low nitrogen stress on the physiological and morphological traits of roots of different low nitrogen tolerance maize varieties at seedling stage[J].Chinese Journal of Eco-Agriculture,2015,23(8):946–953.
[3]曹蘭芹,伍曉明,楊睿,等.不同氮吸收效率品種油菜氮素營養特性的差異[J].作物學報,2012,38(5):887–895. Cao LQ,Wu XM,Yang R,et al.Differences of nitrogen status between different N-uptake-efficiency rape-seed(Brassica napus L.) cultivars[J].Acta Agronomica Sinica,2012,38(5):887–895.
[4]吳禮樹.土壤肥料學[M].北京:中國農業出版社,2011. Wu SL.Soli fertilizer science[M].Beijing:China Agriculture Press, 2011.
[5]Wang GL,Ding GD,Li L,et al.Identification and characterizationof improved nitrogen efficiency in interspecific hybridized new-type Brassica napus[J].Annals of Botany,2014,114(3):549–559.
[6]Ye X,Hong J,Shi L,et al.Adaptability mechanism of nitrogenefficient germplasm of natural variation to low nitrogen stress in Brassica napus[J].Journal of Plant Nutrition,2010,33(13): 2028–2040.
[7]Tang YF,Sun XC,Hu CX,et al.Genotypic differences in nitrate uptake,translocation and assimilation of two Chinese cabbage cultivars[Brassica campestris L.ssp.Chinensis(L.)][J].Plant Physiology and Biochemistry,2013,70:14–20.
[8]Luo JK,Sun SB,Jia LJ,et al.The mechanism of nitrate accumulation in pakchoi[Brassica campestris L.)][J].Plant and Soil, 2006,282:291–300.
[9]劉強,宋海星,榮湘民,等.不同品種油菜子粒產量及氮效率差異研究[J].植物營養與肥料學報,2009,15(4):898–903. Liu Q,Song HX,Rong XM,et al.Study on oilseed yield and nitrogen efficiency in different cultivars of oilseed rape(Brassica napus)[J].Plant Nutrition and Fertilizer Science,2009,15(4): 898–903.
[10]李銀水,余常兵,廖星,等.湖北省不同油菜輪作模式下作物施肥現狀調查[J].中國農學通報,2012,28(36):205–211. Li YS,Yu CB,Liao X,et al.Investigation of present fertilization on crops by different rapeseed rotation systems in Hubei province[J]. Chinese Agricultural Science Bulletin,2012,28(36):205–211.
[11]徐華麗,魯劍巍,李小坤,等.四川省油菜施肥狀況調查[J].西南農業學報,2011,24(4):1372–1376. Xu HL,Lu JW,Li XK,et al.Investigation of fertilizer condition of rapeseed in Sichuan province[J].Southwest China Journal of Agricultural Sciences,2011,24(4):1372–1376.
[12]王小英,劉芬,同延安,等.陜南秦巴山區油菜施肥現狀評價[J].中國油料作物學報,2013,35(2):190–195. Wang XY,Liu F,Tong YA,et al.Fertilization evaluation of rapeseed in Qin-Ba mountain area of southern Shanxi province[J]. Chinese Journal of Oil Crop Sciences,2013,35(2):190–195.
[13]鄒琦.植物生理生化實驗指導[M].北京:中國農業出版社,1995. Zou Q.Guide of plant physiology and biochemistry experiment[M]. Beijing:China Agriculture Press,1995.
[14]Fan XR,Jia LJ,Li YL,et al.Comparing nitrate storage and remobilization in two rice cultivars that differ in their nitrogen use efficiency[J].Journal of Experiment Botany,2007,58(7): 1729–1740.
[15]楊蘭.生長調節劑對油菜氮素再利用的影響及其與氮素利用效率的關系研究[D].長沙:湖南農業大學碩士學位論文,2012. Yang L.Effect of growth regulators on nitrogen reutilization of oilseed rape and relation to nitrogen utilization efficiency[D]. Changsha:MS Thesis,Hunan Agriculture University,2012.
[16]鮑士旦.土壤農化分析(第三版)[M].北京:中國農業出版社,1999. Bao SD.Agricultural chemistry analysis of soil(3rd Ed.)[M]. Beijing:China Agriculture Press,1999.
[17]Matimati I,Verboom GA,Cramer MD.Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients[J].Journal of Experimental Botany,2013,65(1):159–168.
[18]Cai J,Chen L,Qu HY,et al.Alteration of nutrient allocation and transporter genes expression in rice under N,P,K,and Mg deficiencies[J].Acta Physiologiae Plantarum,2012,34(3):939–946.
[19]孫虎威,王文亮,劉尚俊,等.低氮脅迫下水稻根系的發生及生長素的響應[J].土壤學報,2014,51(5):1096–1102. Sun HW,Wang WL,Liu SJ,et al.Formation of rice root regulated by nitrogen deficiency[J].Acta Pedologica Sinica,2014,51(5): 1096–1102.
[20]Cramer MD,Hoffmann V,Verboom GA.Nutrient availability moderates transpiration in Erharta calycina[J].New Phytologist, 2008,179(4):1048–1057.
[21]Kupper P,Rohula G,Saksing L,et al.Does soil nutrient availability influence night-time water flux of aspen saplings?[J].Environmental and Experimental Botany,2012,82:37–42.
[22]Smirnoff N,Stewart GR.Nitrate assimilation and translocation by higher plants:comparative physiology and ecological consequences[J].Physiologia Plantarum,1985,64(2):133–140.
[23]Han YL,Song HX,Liao Q,et al.Nitrogen use efficiency is mediated by vacuolar nitrate sequestration capacity in roots of Brassica napus[J].Plant Physiology,2016,170:1684–1698.
[24]Tanner W,Beevers H.Does transpiration have an essential function in long-distance ion transport in plants?[J].Plant,Cell and Environment,1990,13(8):745–750.
[25]Tanner W,Beevers H.Transpiration,a prerequisite for long distance transport of minerals in plants?[J].Proceedings of the National Academy of Sciences,2001,98(16):9443–9447.
[26]Guo FQ,Young J,Crawford NM.The nitrate transporter AtNRT1.1 (CHL1)functions in stomatal opening and contributes to drought susceptibility in Arabidopsis[J].The Plant Cell,2003,15(1): 107–117.
[27]Broadley MR,Escobar-Gutierrez AJ,Burns A,et al.Nitrogenlimited growth of lettuce is associated with lower stomatal conductance[J].New Phytologist,2001,152(1):97–106.
RONG Nan,HAN Yong-liang*,RONG Xiang-min,SONG Hai-xing,PENG Jian-wei,XIE Gui-xian,ZHANG Yu-ping,ZHANG Zhen-hua*
(Southern Regional Collaborative Innovation Center for Grain and Oil Crops in China/College of Resources and Environment, Hunan Agricultural University, Changsha 410128)
【Objectives】This research was conducted in order to investigate response of NO3–uptake and distribution to limited nitrogen(N)stress and its influence on nitrogen use efficiency(NUE)in oilseed rape,and supply scientific basis for understanding the physiological and NUE changes under limited-N stress condition.【Methods】Oilseed rape genotype814was used as plant materials,and sand culture experiment was conducted under normal-N(10mmol/L)and limited-N(3mmol/L,1mmol/L)levels.The response of root characteristics, transpiration,NO3–distribution and assimilation to limited-N stress and its influence on NO3–uptake efficiency and NUE were studied.【Results】Compared with the normal-N level,NO3–content,total N,root biomass and root uptake area were decreased significantly under limited-N level(3mmol/L,1mmol/L),but ratio of(NO3–) shoot/root and(total N)shoot/root,ratio of root biomass/shoot biomass were increased significantly.Stomatal conductance and transpiration rate were increased significantly in oilseed rape under limited-N stress,which not only stimulated NO3–uptake,but also higher proportion of NO3–distributed from root to shoot;meanwhile,wateruse efficiency was decreased significantly.There were no significant differences of NR and GS in root and shoot between normal-N and limited-N treatments,chlorophyll content and photosynthesis were decreased significantly, but photosynthetic NUE was increased significantly.【Conclusions】Total Nand biomass of oilseed rape plants were decreased significantly under limited-N stress,but the NUE was increased significantly,which is result from the higher proportion of NO3–distributed from root to shoot and higher photosynthetic NUE under limited-N stress.
oilseed rape;nitrate;N uptake efficiency;N use efficiency
2016–08–09接受日期:2016–11–09
土壤與農業可持續發展國家重點實驗室開放課題(Y20160015);湖南農業大學大學生科技創新基金(2016ZK13);湖南農業大學青年科學基金(16QN35)資助。
榮楠(1996—),女,湖南東安縣人,本科,主要從事植物營養生理方面的研究。E-mail:423318689@qq.com
*通信作者E-mail:xiaohliang@163.com;E-mail:zhzh1468@163.com
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
