煙草NtPLR1基因克隆與表達分析
2017-08-16 06:01:09李冰冰劉國峰黃龍全張劍韻
浙江農林大學學報 2017年4期
李冰冰,劉國峰,魏 書,黃龍全,張劍韻
(1.安徽農業大學 茶與食品科技學院,安徽 合肥 2 30036;2.安徽農業大學 外國語學院,安徽 合 肥230036)
煙草NtPLR1基因克隆與表達分析
李冰冰1,劉國峰1,魏 書1,黃龍全1,張劍韻2
(1.安徽農業大學 茶與食品科技學院,安徽 合肥 2 30036;2.安徽農業大學 外國語學院,安徽 合 肥230036)
維生素B6(VB6)在植物體內參與多種生化反應,對植物生長至關重要,吡哆醛還原酶(PLR)是VB6代謝轉換的作用酶,催化吡哆醛(PL)生成吡哆醇(PN),對維持細胞內VB6的動態平衡發揮重要作用,而PLR在植物中鮮有報道。以擬南芥Arabidopsis thaliana吡哆醛還原酶氨基酸序列AtPLR1為模板,在公用數據庫通過同源比對獲得數條煙草Nicotiana tabacum NtPLR1基因的片段,結合互補脫氧核糖核酸(cDNA)的末端快速擴增-聚合酶鏈式反應(RACE-PCR)技術獲得了煙草吡哆醛還原酶NtPLR1基因。該基因全長1 370 bp,編碼369個氨基酸殘基,預測其編碼蛋白的分子量為41 kDa,理論等電點為9.42。氨基酸多序列比對結果表明:NtPLR1與其他物種的PLR1相似性較高。實時熒光定量PCR(qRT-PCR)分析結果表明:外源吡哆醛PL處理時,NtPLR1表達先升高后降低,在4 d達到頂峰。相應地,高效液相色譜分析結果表明:煙草葉片中PL含量隨時間逐漸降低而吡哆醇PN含量逐漸升高,表明NtPLR1可像酵母PLR一樣,催化PL形成PN。此外,定量分析結果表明:NtPLR1在煙草根、莖和葉片中均有表達,其中在葉片中表達顯著高于其他部位(P<0.05)。在紫外線、氧化和鹽害脅迫下,NtPLR1的表達與對照相比均顯著上調(P<0.05),表明NtPLR1對這3種逆境有響應,可能參與煙草的抗逆過程。將NtPLR1連入原核表達載體pET32a,并進行誘導表達,成功表達出目的蛋白。報道的煙草NtPLR1基因功能為進一步探明植物PLR基因的功能和調控機制以及VB6的生物合成提供了重要參考。圖8表1參25
植物學;維生素B6;煙草;NtPLR1;基因克隆;表達分析
維生素B6(VB6)是一類吡啶化合物的總稱,包括吡哆醛(PL),吡哆醇(PN),吡哆胺(PM),磷酸吡哆醛(PLP),磷酸吡哆醇(PNP),磷酸吡哆胺(PMP)等,其中PLP是其主要活性形式,作為輔酶參與生物體內100多種生化反應,包括氨基酸代謝、抗生素合成、免疫調節等生理反應及氧化脅迫等抗逆反應[1]。細胞內VB6各組分的平衡是機體進行正常代謝的前提,因而VB6對植物的生長發育至關重要。研究[2]發現,自然界中VB6有從頭合成(de novo synthetic pathway)和補救合成(salvage pathway)2種方式,補救合成途徑使VB6異養型生物能夠利用外源攝入的PN,PM和PL來合成機體代謝所需要的活化型PLP并維持細胞各型VB6濃度相對穩定。從頭合成已被廣泛研究,而補救合成途徑的研究卻相對缺乏。其中,吡哆醛還原酶(PLR)是VB6補救合成途徑中的作用酶,最初在酵母中被發現,屬于醛酮還原酶(aldo-keto reductase),在還原型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)存在的條件下催化PL轉換成PN[3-4],從而可維持細胞內VB6動態平衡,對于生物體進行正常的生理活動具有重要意義。植物性食物中VB6主要以PN或其糖基化形態存在,并推測植物中可能有高效的PN生成機制[5]。迄今為止,植物中只有擬南芥Arabidopsis thaliana的AtPLR1基因得到分離鑒定,對酵母突變體進行互補實驗表明AtPLR1像酵母PLR一樣,可催化PL形成PN[6]。T-DNA插入的Atplr1突變體根系生長較野生型明顯緩慢,氯化鈉、甘露醇脅迫下Atplr1的生長受到抑制[6],推測PLR可能與植物抵抗鹽害和滲透壓脅迫有關。煙草Nicotiana tabacum是一種重要的模式植物及經濟作物,對其PLR進行克隆和功能分析,有助于進一步明確植物體內VB6的補救合成途徑,同時為煙草良種選育提供理論儲備。本研究以AtPLR1為模板,經過對美國生物技術信息中心(NCBI)公共數據庫(http://www.ncbi.nlm.nih.gov/)中的序列進行比對和拼接并結合cDNA末端快速擴增技術-聚合酶鏈式反應(RACE-PCR)得到了煙草NtPLR1基因的全長序列。以此為基礎,分析了其生物功能和表達特性,結果NtPLR1可催化PL形成PN。NtPLR1在葉片中表達最高,與紫外線、氧化及氯化鈉脅迫和外源PL處理有應答反應。
1 材料與方法
1.1 實驗材料
煙草 ‘云煙85’Nicotiana tabacum‘Yunyan 85’種子和pET32a原核表達載體為實驗室保存。pEASY-Blunt載體、菌株BL21(DE3)Rosetta和大腸埃希菌Escherichia coli DH5α購自TransGen公司。
1.2 處理方法
紫外線處理:在無菌操作臺上用紫外線照射生長至旺長期的煙草,照射時間分別為2 h,4 h和8 h,取莖尖以下的第3片葉,液氮冷凍,備用。
鹽處理:用100.0mmol·L-1的氯化鈉溶液澆灌旺長期煙草,處理1 d,4 d和7 d后取樣,取莖尖以下第2片葉。液氮冷凍,備用。
氧化處理:澆灌亞硫酸氫鈉-亞硫酸鈉(NaHSO3-Na2SO3)混合物(10.0mmol·L-1,以亞硫酸鈉濃度計),處理1 d,4 d和7 d后取樣,取莖尖以下第2片葉,液氮冷凍,備用。
PL處理:將旺長期煙草根部洗凈,置于添加100.0mg·L-1PL的水培液中,用錫紙將煙草根部及水培液遮住,莖葉接受正常光照。分別于培養的第2天、第4天和第8天采集莖尖以下第2片葉,液氮冷凍,備用。
以上均設置重復3個·處理-1。
1.3 總核糖核酸(RNA)提取及互補脫氧核糖核酸(cDNA)的合成
根據RNAiso Plus RNA提取試劑盒使用說明書(Takara),進行總RNA提取。提取的RNA用質量分數為1.0%瓊脂糖凝膠電泳進行純度與完整性檢測。參照PrimeScript RT reagent Kitwith gDNA Eraser反轉錄試劑盒(Takara)對質量合格的RNA進行反轉錄,置于-20℃備用。
1.4 基因克隆及生物信息學分析

表1 NtPLR1基因克隆與表達分析所用引物信息Table 1 Primers used in NtPLR1 gene cloning and expression analysis
以擬南芥吡哆醛PLR的氨基酸序列(NP_200170.2)為模板,在煙草表達序列標簽(EST)數據庫里同源檢索,根據得到的EST序 列 , 設 計 引 物 3RACE-1,3RACE-2,3RACE-3(表 1)進行巢式PCR擴增目的基因3′端序列。擴增產物經切膠回收,連接pEASY-Blunt載體后,轉化至大腸埃希菌DH5α感受態細胞,篩選陽性克隆子進行測序。3′端序列擴增產物測序結果驗證后,使用DNAMAN軟件將它與EST起始序列結合得到全長cDNA序列。隨后設計全長克隆引物NtPLR-F1和NtPLR-R1(表1),擴增NtPLR1序列。PCR反應程序為95℃預變性3.0 min;94℃變性30 s,60℃退火30 s,72℃延伸1.5 min,共28個循環;72℃延伸10.0 min。獲得的NtPLR1序列在NCBI(http://www. ncbi.nlm.nih.gov/)數據庫BLAST中進行序列比對。用ProtParam軟件(http://web.expasy.org/protparam/)在線分析該蛋白的分子量和等電點;采用DNAMAN 7.0軟件對基因編碼的氨基酸序列進行比對分析。
1.5 NtPLR1表達特性分析
根據NtPLR1基因的cDNA序列,設計特異性定量引物(表1),以18 S rRNA為內參基因,以根、莖、葉及紫外線、氧化、氯化鈉處理下不同時間點取樣葉片的cDNA為模板,進行熒光實時定量PCR(qRT-PCR)分析。qRT-PCR反應程序為95℃3.0 min,95℃10 s,55℃40 s,35個循環;溶解曲線:從65℃ 按0.5℃/循環增加到95℃ 。以2-ΔΔCt法計算相對表達量。
1.6 VB6標準試劑的配制及VB6的色譜分析條件
VB6檢測參照張劍韻等[7-9]的方法,加以改進。VB6色譜分析所用色譜柱為H&E公司的XPODS-A 5 μm 120 A(250.0mm×4.6mm)。高效液相色譜儀為Waters 600,配備2475熒光檢測儀。流動相A(分析用):體積分數為1%乙腈(CH3CN)-25.0 mmol·L-1磷酸二氫鉀(KH2PO4)-25.0 mmol·L-1高氯酸鈉(Na-ClO4),pH 2.5;流速為0.5 mL·min-1。進樣量均為5.0μL,熒光檢測波長為395 nm,調整激發波長為290 nm。
1.7 基因原核表達載體的構建及蛋白誘導表達
原核表達載體構建:根據載體pET32a多克隆位點信息,設計帶有酶切位點(Bam H1和Sal1)的原核表達引物(表1),以云煙85 cDNA為模板擴增NtPLR1基因的cDNA片段。經瓊脂糖凝膠電泳分離,回收目的條帶后,連接pEASY-Blunt載體,轉化至大腸埃希菌DH5α感受態細胞。挑取經PCR和測序驗證的陽性菌落,擴大培養,使用質粒小抽試劑盒(TransGen2)提取質粒,即得到pEASY-NtPLR1載體。用限制性內切酶BamH1和Sal1雙酶切pEASY-NtPLR1和pET32a質粒后進行T4連接,即得到pET32a-Nt-PLR1重組質粒。測序正確后,提取目的質粒并轉化至BL21(DE3)Rosetta菌株,即得到融合表達菌。
蛋白誘導表達:37℃培養融合表達菌至D(600)約為0.60,加入終濃度為1.0 mmol·L-1的異丙基硫代半乳糖苷(IPTG),并在18℃/37℃200 r·min-1誘導24 h。以含pET32a空質粒的BL21(DE3)Rosetta為對照。離心收獲菌體,8 000 r·min-1離心10.0min,棄上清,用PBS重懸。重復1次后進行超聲波破碎。分別取上清和沉淀,加入上樣緩沖液,沸水浴5.0 min,冷卻至室溫后,取20.0μL進行SDS-PAGE(5%濃縮膠,10%分離膠)電泳檢測。電泳后,經考馬斯亮藍染色、拍照,分析蛋白表達結果。
2 結果與分析
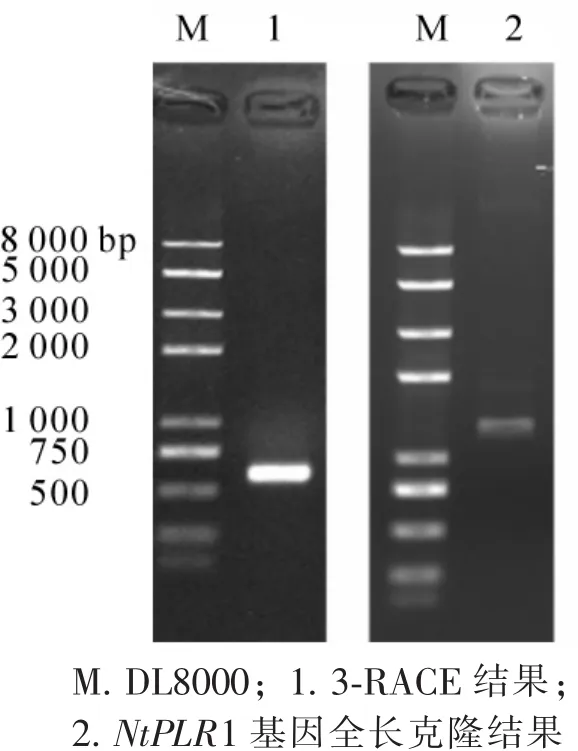
圖1 瓊脂糖凝膠電泳結果Figure 1 Agorase gel electrophoresis results
2.1 NtPLR1克隆及氨基酸序列分析
以擬南芥吡哆醛PLR的氨基酸序列(NP_200170.2)為模板,在煙草EST數據庫里同源檢索到一條同源性(79%)序列(GenBank: HS082453.1)。以此EST序列為起點,進行延伸檢索后得到4條候選EST序列,登錄號分別為GenBank:FS425789,GenBank:FS385536.1,GenBank:FS432618,GenBank:FS431044.1。使用DNAMAN比對5條EST序列,發現它們來源于同一基因,拼接后得到1條長800 bp的起始序列。根據該序列設計3輪3′-RACE引物(表1),進行巢式PCR擴增得到大小約為700 bp單一明亮條帶(圖1中泳道1)。將該條帶測序后和起始序列拼接得到全長cDNA序列。隨后設計全長克隆引物(表1),PCR擴增得到大小約1 500 bp的序列(圖1中泳道2)。測序正確后,將此全長序列命名為NtPLR1,其 cDNA長度為1 370 bp,開放閱讀框1 110 bp,5′非編碼區(UTR)長55 bp,3′UTR長205 bp。編碼369個氨基酸,起始密碼子為ATG,終止密碼子為TGA(圖2),具有Aldo-keto還原酶家族保守底物結合位點[6](圖2中下劃線強調部分)。在線預測其編碼蛋白的分子量為41 070.5 Da,理論等電點為9.42。氨基酸多序列比對結果顯示,NtPLR1與AtPLR相似性為75%(圖3),與栗酒裂殖酵母Schizosaccharomyces pombe[10],釀酒酵母Saccharomyces cerevisiae[11]PLR的氨基酸相似性分別為24%和26%。
2.2 NtPLR1表達特性分析
分別提取煙草根、莖、葉的總RNA,D(260)/D(280)為1.9~2.0,表明RNA純度較好,可進行后續實驗。如圖4A所示:NtPLR1在根、莖和葉均有表達,在葉中表達最高,根、莖表達水平較低。對不同逆境脅迫下NtPLR1的表達分析發現:紫外線脅迫下,隨時間延長,NtPLR1基因在煙草葉片中的表達量表現出先升高后下降的趨勢,并在紫外線處理4 h時達到最大值(圖4B)。氧化及氯化鈉(100.0mmol· L-1)澆灌處理時,隨脅迫時間的增加,NtPLR1基因在煙草葉片中的表達持續升高,7 d時表達最高(圖4C)。在以上逆境脅迫下,NtPLR1表達呈現不同程度的上調,這表明NtPLR1與紫外線、氧化、氯化鈉脅迫有應答反應。
2.3 PL對NtPLR1表達的誘導效果及對PN生成量的影響
煙草水培液添加外源PL后,分別于培養的第2天、第4天、第8天取樣,分析NtPLR1基因的表達水平和PL,PN含量。定量PCR分析結果表明:NtPLR1表達隨處理時間延長呈現先上升后下降的趨勢,在第4天表達量達到最高,第2天、第4天、第8天的NtPLR1表達量分別是對照的2.20,2.85和1.50倍(圖5)。
VB6標準品高效液相色譜法(HPLC)檢測結果如圖6A,峰型和區分度良好。對未處理的煙草葉片提取液分析發現,在PMP,PM,PLP,PL及PN的洗脫位置上均出現了相應的洗脫峰(圖6B),說明檢測方法可行。據此,對PL處理組煙草葉片進行HPLC分析,結果表明:隨時間延長,處理組煙草葉片中PL含量逐漸降低,PN含量增幅明顯(圖7),同時,PMP,PM含量有小幅增長,表明煙草吸收外源PL后,主要將PL轉化為PN。PL處理后NtPLR1的表達受到誘導,而在8 d時表達下降,結合PN,PMP和PM含量的逐漸增多可知,NtPLR1在煙草中催化PL形成PN。VB6各組分在煙草中動態轉化,且各組分間存在反饋調節。
2.4 NtPLR1原核載體構建及重組蛋白誘導表達
將重組質粒pET32a-NtPLR1轉入BL21(DE3)Rosetta,分別在28℃和37℃條件下經異丙基硫代半乳糖苷(IPTG)誘導后,超聲波破碎菌體,離心分離上清和沉淀。將上清和沉淀分別進行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)檢測,結果顯示:上清中無目的條帶,而沉淀中在約53 kDa處出現明顯蛋白條帶,因pET32a的組氨酸標簽(his-tag)約為12 kDa,故蛋白條帶的大小與預期相符。僅沉淀中出現目的條帶,表明NtPLR1在大腸埃希菌中以包涵體形式存在(圖8)。
3 討論
VB6在自然界中廣泛地存在,主要以輔酶的形式參與生物體內多種物質代謝反應,是生物機體內很多重要酶系的輔酶[12]。植物體內,VB6參與淀粉、亞油酸等物質的合成[13],對于生長素、葉綠素以及乙烯的合成是不可或缺的[14-15]。近年來的研究還發現,VB6具有抗氧化作用,可猝滅超氧陰離子自由基及單線態氧[16];除此之外,在低溫、滲透壓、鹽害、紫外及病菌等逆境中,VB6可以提高植株的抵抗力,發揮一定的抗逆作用[13,17-20]。
VB6從頭合成途徑和補救合成途徑普遍存在于植物和微生物中[21-24]。植物和微生物是VB6自養生物,動物自身無法從頭合成VB6,只能從食物中獲得VB6前體物質,通過補救合成途徑滿足機體對VB6的需求。VB6補救途徑由多種酶參與,PLR是其中一種VB6補救合成酶,對于細胞進行正常生理活動具有重要意義。在研究擬南芥Atplr1時發現,Atplr1的VB6總水平下降,其中PL,PLP,PM和PMP水平顯著下降,而PN和PNP無顯著變化,推測在擬南芥內可能存在PLR的同工酶[6]。而HUANG等[25]的研究認為:煙草葉際PL—PN的轉換可能受葉際微生物的影響較大。VB6對植物的生長發育、逆境適應及人和動物的營養具有重要意義,其從頭合成途徑已有較多的研究,而補救途徑還有許多不明之處,有待深入研究。本研究從煙草中克隆得到煙草NtPLR1,并設置了不同的逆境脅迫條件對NtPLR1進行探究,結果表明:NtPLR1在葉中表達最高且受紫外線,氧化和氯化鈉脅迫的誘導,推測NtPLR1參與煙草植株對紫外線、氧化和氯化鈉脅迫的抗逆反應。外源添加PL后,NtPLR1表達量與對照相比顯著上調,表明PL對NtPLR1有顯著的誘導作用。此外,外源PL處理的前4 d NtPLR1的表達呈上升趨勢,而在培養的第8天時NtPLR1的表達降低,相應的PL持續降低,而PN,PM和PMP等都有不同程度的升高,表明VB6各組分在煙草葉片中可相互轉化并存在反饋調節。目前,本實驗室正在進行NtPLR1重組蛋白的優化表達及體外酶活測定,以期為進一步探明煙草PLR基因功能及VB6補救合成過程奠定基礎。

圖2 NtPLR1基因開放閱讀框及預測氨基酸序列Figure 2 ORF of NtPLR1 gene and the corresponding amino acid sequence

圖3 幾個已知物種的PLR多序列比對Figure 3 Multiple protein sequence alignment of several known PLR enzymes
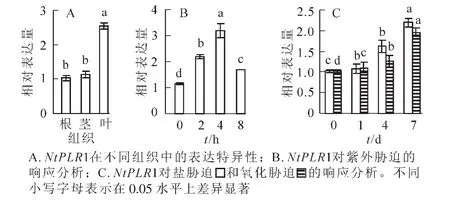
圖4 NtPLR1表達的組織特異性及對不同脅迫處理的響應Figure 4 QPCR analysis of spatial expression of NtPLR1 and its response to different stress treatments
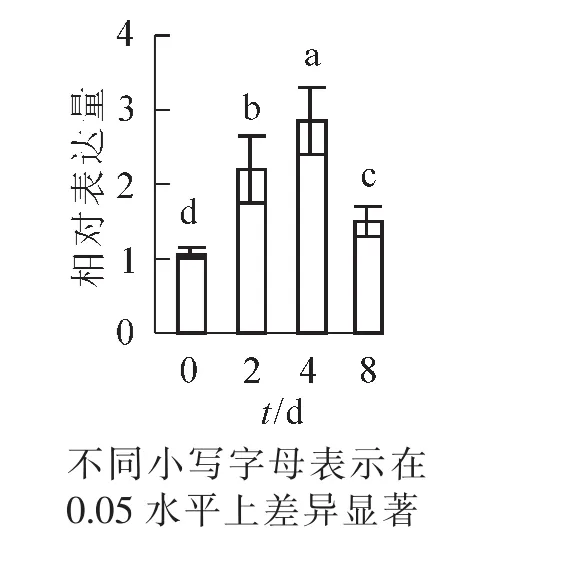
圖5 NtPLR1對外源PL處理的響應分析Figure 5 QPCR analysis of the response of NtPLR1 to exogenous PL

圖6 HPLC分析VB6標準品(A)及對照煙草葉片提取液(B)Figure 6 HPLC analysis for VB6authentic standards(A)and extracts from control tobacco leaves(B)
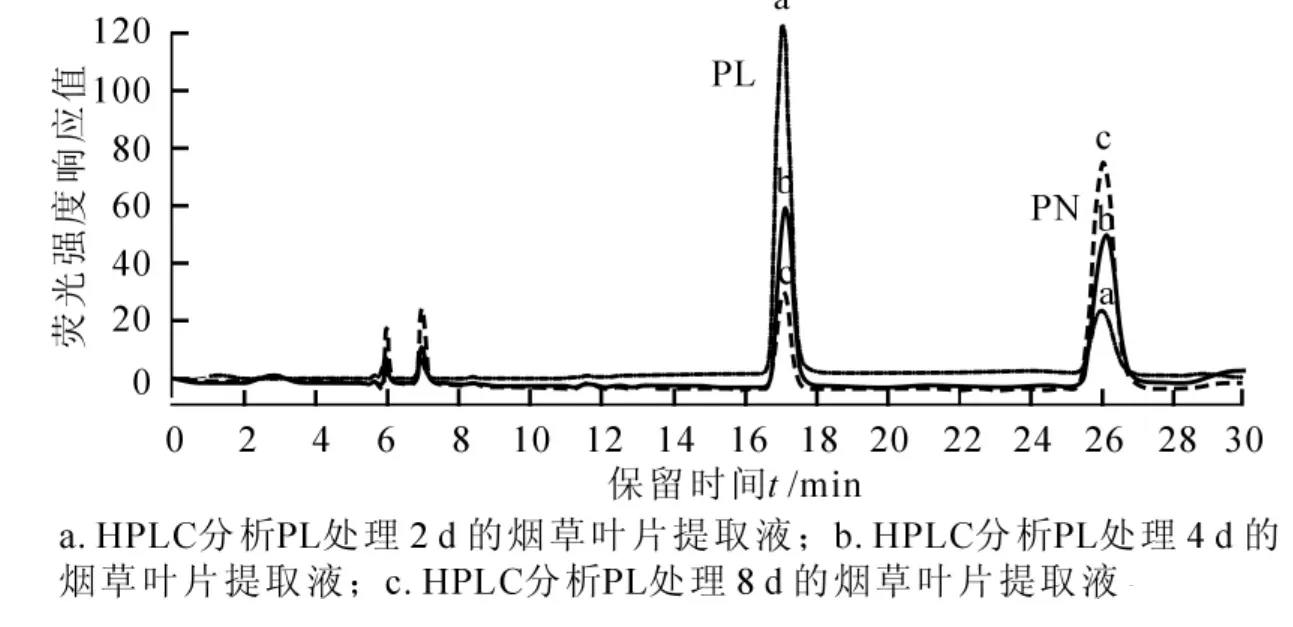
圖7 HPLC分析外源PL處理組煙草葉片提取液Figure 7 HPLC analysis of extracts from exogenous PL treated tobacco leaves
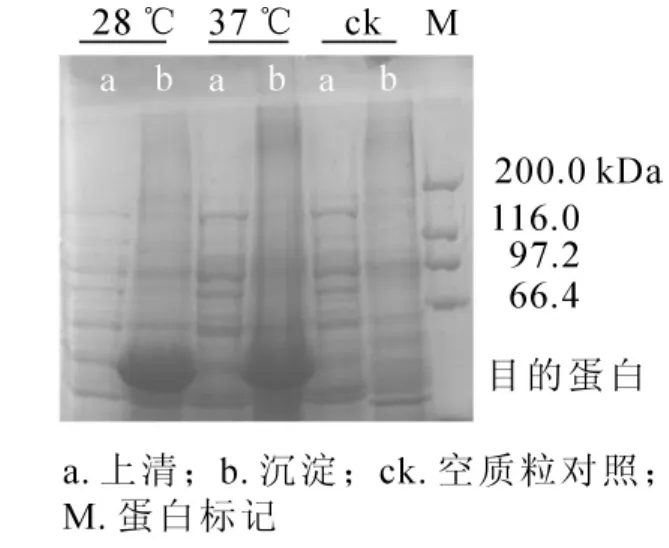
圖8 NtPLR1在表達菌株BL21(DE3)Rosetta中的表達Figure 8 Prokaryotic expression of NtPLR1 in BL21(DE3)Rosetta
[1] RAIL L C,NIKBINM S.Vitamin B6and immune competence[J].Nutr Rev,1993,51(8):217-225.
[2] HUANG Shuohao,ZENG Haibin,ZHANG Jianyun,et al.Interconversions of different forms of vitamin B6in tobacco plants[J].Phytochemistry,2011,72(17):2124-2129.
[3] MORINO Y,SAKAMOTO Y.Enzymatic studies on pyridoxinemetabolism(Ⅳ)a pyridoxine dehydrogenase from baker’s yeast[J].JBiochem,1960,48:733-744.
[4] HOLZER H,SCHNEIDER S.Purification and characterization of a TPN-dependent pyridoxol dehydrogenase from brewers yeast[J].Biochim Biophys Acta,1961,48(2):71-76.
[5] OLLILAINEN V.HPLC analysis of vitamin B6in foods[J].Agric Food Sci Finland,1999,8(6):515-619.
[6] HERRERO S,GONZALEZ E,GILLIKIN JW,et al.Identification and characterization of a pyridoxal reductase involved in the vitamin B6salvage pathway in Arabidopsis[J].PlantMol Biol,2011,76(1/2):157-169.
[7] 蔣守花,張劍韻,黃龍全.采用高效液相色譜技術分析茶樹體內維生素B6[J].茶葉科學,2010,30(2):79-82.
JIANG Shouhua,ZHANG Jianyun,HUANG Longquan.Analysis of VB6derivatives in tea plantwith high performance liquid chromatography[J].JTea Sci,2010,30(2):79-82.
[8] 張劍韻,黃龍全,早川享志,等.采用高效液相色譜技術分析生物體內維生素B6[J].高等學校化學學報,2004,25(4):638-640.
ZHANG Jianyun,HUANG Longquan,HAYAKAWA T,et al.Analysis of VB6derivatives in biological samples with high performance liquid chromatography[J].Chem JChin Univ,2004,25(4):638-640.
[9] 曾海彬,張劍韻,黃龍全.采用高效液相色譜技術分析煙草體內的維生素B6化合物[J].廣西植物,2011,31(5):695-698.
ZENG Haibin,ZHANG Jianyun,HUANG Longquan.Analysis of vitamin B6vitamers in tobacco plants by high performance liquid chromatography[J].Guihaia,2011,31(5):695-698.
[10] GUIRARD B M,SNELL E E.Physical and kinetic properties of a pyridoxal reductase purified from baker’s yeast[J].Biofactors,1988,1(2):187-192.
[11] NAKANOM,MORITA T,YAMAMOTO T,et al.Purification,molecular cloning,and catalytic activity of Schizosaccharomyces pombe pyridoxal reductase a possible additional family in the aldo-keto reductase superfamily[J].JBiol Chem,1999,274(33):23185-23190.
[12] LYON JB,BAIN JA,WILLIAMSH L.The distribution of vitamin B6in the tissues of two inbred strains ofmice fed complete and vitamin B6-deficient rations[J].JBiol Chem,1962,237(6):1989-1991.
[13] MOONEY S,HELLMANN H.Vitamin B6:killing two birdswith one stone?[J].Phytochemistry,2010,71(5/6):495-501.
[14] TAO Yi,FERRER J L,LJUNG K,et al.Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants[J].Cell,2008,133(1):164-176.
[15] VAVILIN D V,VERMAASW F J.Regulation of the tetrapyrrole biosynthetic pathway leading to heme and chlorophyll in plants and cyanobacteria[J].Physiol Plantarum,2002,115(1):9-24.
[16] BILSKIP,LIM Y,EHRENSHAFTM,et al.Vitamin B6(pyridoxine)and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants[J].Photochem Photobiol,2000,71(2):129-134.
[17] SHIHuazhong,XIONG Liming,STEVENSON B,et al.The Arobidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6in plant salt tolerance[J].Plant Cell,2002,14(3):575-588.
[18] CHEN Hao,XIONG Liming.Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses[J].Plant J,2005,44(3):396-408.
[19] ZHANG Yafen,JIN Xiaoyi,OUYANG Zhigang,et al.Vitamin B6contributes to disease resistance against Pseudomonas syringae pv.tomato DC3000 and Botrytis cinerea in Arabidopsis thaliana[J].JPlant Physiol,2015,175(1):21-25.
[20] 黃龍全,張劍韻.植物維生素B6從頭合成與代謝轉換研究進展[J].西北植物學報,2015,35(10):2124-2131.
HUANG Longquan,ZHANG Jianyun.Review on the de novo synthesis and metabolic conversions of vitamin B6in plants[J].Acta Bot Boreal-Occident Sin,2015,35(10):2124-2131.
[21] YU Shunwu,LUO Lijun.Expression analysis of a novel pyridoxal kinasemessenger RNA splice variant,PKL,in oil rape suffering abiotic stress and phytohormones[J].Acta Biochim Biophys Sin,2008,40(12):1005-1014.
[22] SANG Yuying,BARBOSA JM,WU Hongzhuan,et al.Identification of a pyridoxine (pyridoxamine)5′-phosphate oxidase from Arabidopsis thaliana[J].FEBSLetters,2007,581(3):344-348.
[23] EHRENSHAFTM,BILSKIP,LIM Y,et al.A highly conserved sequence is a novel gene involved in de novo vitamin B6biosynthesis[J].Proc Nat Acad Sci USA,1999,96(16):9374-9378.
[24] MITTENHUBER G.Phylogenetic analyses and comparative genomics of vitamin B6(pyridoxine)and pyridoxal phosphate biosynthesis pathways[J].JMol Microbiol Biotechnol,2001,3(1):1-20.
[25] HUANG Shuohao,ZHANG Jianyun,TAO Zhen,et al.Enzymatic conversion from pyridoxal to pyridoxine caused by microorganismswithin tobacco phyllosphere[J].Plant Physiol Biochem,2014,85:9-13.
Cloning and expression analysis of the tobacco NtPLR1 gene
LIBingbing1,LIU Guofeng1,WEIShu1,HUANG Longquan1,ZHANG Jianyun2
(1.School of Tea and Food Science,Anhui Agricultural University,Hefei230036,Anhui,China;2.School of Foreign Languages,Anhui Agricultural University,Hefei230036,Anhui,China)
Vitamin B6(VB6),essential for plant growth and developmentand involved inmore than 100 biological processes,utilizes pyridoxal reductase (PLR)as the key enzyme in the VB6salvage pathway,thereby catalyzing pyridoxal(PL)to generate pyridoxine(PN).Since studies on PLR of plant VB6are quite limited,PLR genes were cloned and characterized to improve understanding of VB6biosynthesis in plants.Several NtPLR1 gene fragments were found in Nicotiana tabacum through a homologous blast with Arabidopsis AtPLR1.Full length was obtained using rapid amplification of cDNA ends(RACE).Real-time quantitative polymerase chain reaction (PCR)and high performance liquid chromatography (HPLC)analysiswere conducted;NtPLR1 expression by ultraviolet,oxidation,exogenous PL,and NaCl treatments were compared to a control;and prokary otic expression of NtPLR1 was accomplished.Results of RACE showed that full length cDNA of NtPLR1 was 1 370 bp,which encoded 369 amino acid residueswith a proteinmolecular weight of about 41 kDa and a theoretical isoelectric point of 9.42.Real-time quantitative PCR analysis revealed that an exogenous PL treatmentinduced NtPLR1 expression with highest expression at 4 d.The HPLC analysis showed that PL content significantly decreased (P<0.05);whereas,PN content significantly increased (P<0.05)during an exogenous PL treatment.NtPLR1 was expressed in roots,stems,and leaveswith leaves having the highest(P<0.05)expression level.Also,ultraviolet,oxidation,and NaCl treatments,compared to a control,significantly induced (P<0.05)NtPLR1 expression.Furthermore,prokaryotic expression of NtPLR1 in vector pET32a successfully revealed the recombinant protein at the expected size.This study reported the NtPLR1 gene of N.tabacum for the first time,finding that it catalyzed PL to form PN in tobacco as found in yeast,and itmay be induced in response to ultraviolet,oxidation,and NaCl stress;thus,the NtPLR1 gene can be an important reference for further plant PLR gene functional characterization and regulation as well as VB6biosynthesis.[Ch,8 fig.1 tab.25 ref.]
botany;vitamin B6;Nicotiana tabacum;NtPLR1;gene cloning;expression analysis
S572;S718.3
A
2095-0756(2017)04-0581-08
10.11833/j.issn.2095-0756.2017.04.003
2016-08-30;
2016-10-27
安徽省教育廳自然科學基金重點資助項目(KJ2010A116);國家自然科學基金面上項目(31670297)
李冰冰,從事維生素B6代謝轉換研究。E-mail:lbb126@126.com。通信作者:張劍韻,教授,博士,從事維生素B6代謝和生物多樣性研究。E-mail:jianyun218@aliyun.com
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
當代化工研究(2016年9期)2016-03-20 16:22:08
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
聲屏世界(2014年6期)2014-02-28 15:18:09
終身教育研究(2014年5期)2014-02-28 01:23:06
中國煙草學報(2012年5期)2012-04-12 06:21:18
