兩種天然產物對B16F10細胞增殖及黑色素合成抑制機理研究
2017-08-07 23:22:05程杏安張淑明周曉武吳波林賢偉秦湘靜黃素青劉展眉蔣旭紅
生物技術通報 2017年8期
程杏安張淑明周曉武吳波林賢偉秦湘靜黃素青劉展眉蔣旭紅
(1. 仲愷農業工程學院天然產物化學研究所,廣州 510225;2. 中國科學院南海海洋研究所,廣州 510301)
兩種天然產物對B16F10細胞增殖及黑色素合成抑制機理研究
程杏安1張淑明1周曉武1吳波1林賢偉1秦湘靜2黃素青1劉展眉1蔣旭紅1
(1. 仲愷農業工程學院天然產物化學研究所,廣州 510225;2. 中國科學院南海海洋研究所,廣州 510301)
旨在研究10-羥基喜樹堿(10-hydroxycamptothecin,HCPT)和白藜蘆醇(Resveratrol,Res)對體外培養的小鼠惡性黑色素瘤B16F10細胞的增殖及黑色素合成抑制機理。利用MTT法、顯微觀察、L-Dopa氧化法、NaOH裂解法分析不同濃度HCPT和Res對細胞增殖、細胞形態、酪氨酸酶活性及黑色素合成含量的影響。熒光半定量PCR方法(Semi-RT-PCR)分析該化合物對黑色素合成關鍵因子酪氨酸酶(TYR)和小眼相關轉錄因子(MITF)基因表達的影響。結果表明HCPT(40、80、120、160和200 μmol/L)和Res(80、120、160和200 μmol/L)能夠通過誘導細胞凋亡抑制B16F10細胞的增殖,同時對酪氨酸酶活性和細胞黑色素生成具有明顯抑制作用(P<0.05),且呈現濃度依賴性。另外,不同濃度的HCPT以及高濃度Res(120和160 μmol/L)能夠顯著下調B16F10細胞TYR和MITF基因的mRNA水平。HCPT和Res可能通過誘導細胞凋亡抑制B16F10細胞的增殖,同時通過下調MITF基因轉錄,抑制TYR mRNA的表達及TYR酶活性,進而抑制細胞黑色素的生成。
10-羥基喜樹堿;白藜蘆醇;酪氨酸酶;黑色素;小眼相關轉錄因子
10-羥 基 喜 樹 堿(10-hydroxycamptothecin,HCPT)是從我國特有珙桐科植物喜樹中分離出的天然生物堿,也是一種新型有效的抗腫瘤藥物[1],通過抑制拓樸異構酶I(Topoosomerase I,Topo-I)干擾腫瘤細胞的DNA復制,抑制腫瘤細胞生長[2,3]。白藜蘆醇(Resveratrol,Res)是廣泛分布在葡萄、花生、桑葚、虎杖、決明等植物中的非黃酮類多酚化合物。近年來有報道Res對肝癌、胃癌、自血病、宮頸癌、食管癌等均有較好的抗腫瘤效果,而且對人體正常機體沒有明顯的毒副作用[4-6]。黑色素瘤是目前位居人類皮膚癌第三位的腫瘤疾病,因其惡性程度高,易轉移,導致患者生存率低,引起了研究者的廣泛重視[7-9]。因此,不斷發掘治療黑色素瘤的新藥物,提高患者的生存率成為當前重要任務。如今醫學界已逐漸注重發揮中醫學的優勢,從天然植物中篩選出具有抗瘤活性的成分作為防治黑色素瘤的重要候選藥物[10]。合成黑色素是黑素瘤細胞增殖生長過程中的重要特征,其過程主要由酪氨酸酶基因家族調控。其中,酪氨酸酶(TYR)是黑色素合成的關鍵限度酶,其基因家族受小眼相關轉錄因子(MITF)的調節。MITF是黑素細胞的特異性轉錄因子,可以直接結合并激活酪氨酸酶啟動子,上調酪氨酸酶基因的表達,促進黑色素的生成[11-13]。本研究以小鼠黑色素瘤B16F10細胞為模型,研究HCPT和Res對該細胞株增殖及黑色素合成的影響,通過分析藥物作用下TYR活性變化、TYR和MITF基因的表達水平,初步闡明兩種天然產物抑制B16F10細胞黑色素合成的作用機理,旨為深入研究HCPT和Res抗黑色素瘤機理及其相關藥物開發提供理論依據。
1 材料與方法
1.1 材料
小鼠黑色素瘤B16F10細胞購自中國科學院菌種保藏中心;10-羥基喜樹堿(純度98%以上)、白藜蘆醇(純度98%以上)、左旋多巴(L-DOPA)購自阿拉丁試劑有限公司;MTT(四甲基偶氮唑藍)、DMSO(二甲基亞砜)購自SIGMA;DMEM高糖培養基、胎牛血清、抗生素購自上海立菲生物技術有限公司;PCR試劑盒和RT-PCR試劑盒購自東盛生物科技有限公司,PCR引物由華大基因公司合成。
1.2 方法
1.2.1 細胞培養 用含10%胎牛血清的DMEM培養基在37℃,5% CO2條件下常規培養B16F10細胞,待細胞生長至近融合狀態,以500 μL胰蛋白酶消化傳代,每3天傳代1次。
1.2.2 細胞活力檢測 取處于最佳生長狀態的B16F10細胞,常規處理,細胞懸浮液密度調整到7×104-9×104個/mL,以每孔100 μL細胞懸液接種于96孔板中,置于37℃、5% CO2培養箱培養。
1.2.2.1 藥物溶劑對細胞活力影響實驗 24 h后,實驗分為3組:實驗組為正常培養和終濃度0.5% DMSO的處理組,調零組只加培養液不接種細胞。設1、2、3、4、5和6 d共6個時間點,每個時間點4個重復。到時間點時,每孔加10 μL MTT(5 mg/mL,PBS配制),常規孵育4 h。棄掉培養液,每孔加150 μL DMSO,室溫震蕩15 min,使紫藍色結晶物充分溶解。使用酶標儀測定各孔492 nm處吸光度(OD)值。記錄結果,OD = OD實驗組- OD調零組。
1.2.2.2 藥試實驗 24 h后,實驗分為兩組:對照組和實驗組(濃度為20、40、80、120、160和200 μmol/L HCPT處理組或濃度40、80、120、160和200 μmol/L Res的處理組)。設24、48、72 h共3個時間點,每個時間點4個重復。到時間點時,每孔加入10 μL MTT,常規孵育4 h。棄掉培養液,每孔加150 μL DMSO,室溫震蕩15 min,使紫藍色結晶物充分溶解。使用酶標儀測定各孔492 nm處OD值。記錄結果。
細胞抑制率(%)=(1-OD實驗組/OD對照組)×100%
1.2.3 倒置生物顯微鏡細胞形態特征觀察 取狀態最佳的B16F10細胞以(7-9)×104個/mL的密度鋪板于6孔細胞培養板中,24 h后,加HCPT和Res至供試濃度進行處理,48 h后用倒置顯微鏡觀察細胞形態變化并拍片記錄。
1.2.4 黑素含量測定 選取處于最佳生長狀態的B16F10細胞,常規處理,細胞懸浮液密度調整到7×104-9×104個/mL,以每孔2 mL細胞懸液接種于6孔板中,置于37℃、5% CO2培養箱培養。24 h后換液,實驗組分別加入不同濃度HCPT(終濃度為20、40、80、120、160、200 μmol/L)和Res(終濃度為40、80、120、160和200 μmol/L)。藥物作用72 h后,收獲細胞,將細胞懸浮液離心后用pH=7.2 PBS洗滌兩次,加入1 mol/L NaOH,置80℃水浴中30 min,使細胞團塊完全溶解,再加入超純水將NaOH的終濃度稀釋至0.2 mol/L。混合均勻,取各組溶液放入96孔板中,每孔100 μL,設立8個重孔。用酶標儀在492 nm測定OD值。黑素含量(%)=(OD實驗組/OD對照組)×100%。
1.2.5 酪氨酸酶活性測定 選取處于最佳生長狀態的B16F10細胞,常規處理,細胞懸液調整細胞濃度為7×104-9×104個/mL,以每孔100 μL接種于96孔板,置于37℃、5% CO2培養箱中培養24 h。實驗分兩組:實驗組調整HCPT終濃度為20、40、80、120、160和200 μmol/L,Res終濃度為40、80、120、160和200 μmol/L,并設對照組,每一濃度設立4個重復。藥物作用72 d后,棄去上清液,用pH 7.2的PBS洗滌,每孔加入1% Triton X-100溶液100 μL,手動平穩震蕩5 min,接著在-20℃冰箱凍存1 h。隨后在室溫中融化,使細胞裂解,37℃預溫后每孔加入0.1% L-DOPA溶液(PBS配制)100 μL,37℃反應2 h,用酶標儀在492 nm測定OD值。酪氨酸酶活性激活率(%)=(OD實驗組/OD對照組)× 100%。
1.2.6 半定量逆轉錄- 聚合酶鏈式反應(Semi-RTPCR)分析法 采用通用RNA提取試劑盒(參照試劑說明書)提取以上述濃度HCPT和Res處理48 h的B16F10細胞總RNA,并反轉錄成cDNA用于PCR擴增。目的基因PCR引物如下。TYR:上游5′-GGCCAGCTTTCAGGCAGAGGT-3′,下游5′-TGGTGCTTCATGGGCAAAATC-3′。MITF:上游5′-AAGTGGTCTGCGGTGTCTCC-3′,下游5′-GTTGTTGGTAAAGGTGATGG-3′。β-actin:上游5′-CGAGCGGGAAATCGTGCGTGACATTAAGGAGA-3′,下游5′-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3′。其中先以對照組的cDNA為模板,PCR的反應條件為:94℃60 s,64℃ 60 s,72℃ 60 s,設定系列循環數(25、26、27、28、29、31、33和35),通過瓊脂糖凝膠中電泳分析,篩選并確定每個基因進行Semi-RTPCR分析的合適循環數,最后再按照該循環數,利用Semi-RT-PCR技術分析各基因mRNA相對表達量。
1.2.7 數據處理 實驗結果用ORIGIN7.5統計軟件處理,數據用均數±標準差(x-±s)表示,組間比較采用單因素方差分析,P<0.05認為兩組間的差異具有統計學意義。
2 結果
2.1 HCPT和Res抑制B16F10細胞增殖
B16F10細胞在接種后第2天完成第1次倍增,進入對數生長階段。藥試試驗中,選擇鋪板培養24 h作為加藥起點時間。以0.5% DMSO處理的B16F10細胞,其生長活力與正常生長細胞相比,總體上沒有明顯差異(圖1),表明DMSO在終濃度低于0.5%的情況下對B16F10細胞活力沒有顯著影響,可作為供試藥物的溶劑。
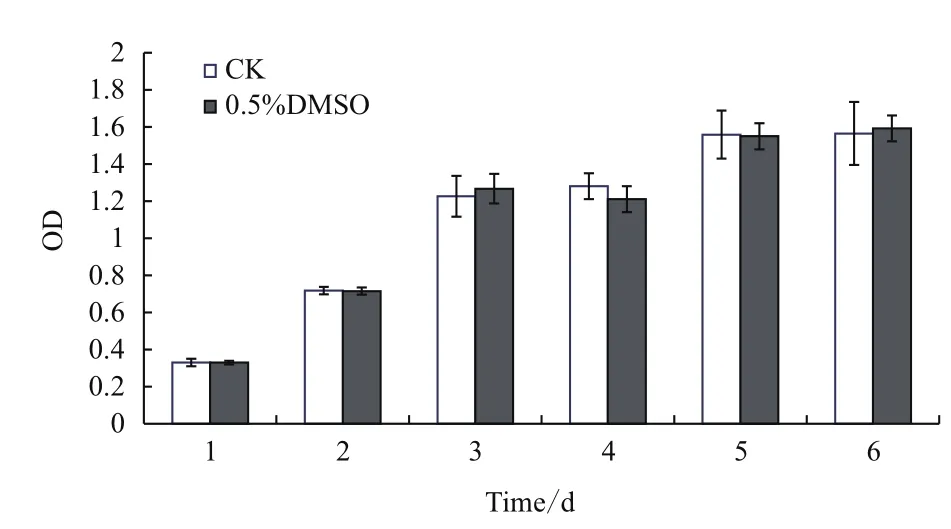
圖1 DMSO對B16F10細胞活力的影響(n=4,)
利用MTT法測定不同濃度HCPT和Res對B16F10細胞增值生長的影響。HCPT誘導B16F10細胞24、48和72 h,細胞增殖均受到明顯抑制,抑制率50%以上,其中24和48 h處理濃度間抑制率差異不明顯;48和72 h處理抑制率比24 h明顯增加,表明HCPT對B16F10細胞增殖抑制存在一定時效性;72 h處理,隨著HCPT濃度增加,細胞抑制率逐漸增加,處理濃度200 μmol/L時,抑制率達到84.20%,表現出一定濃度依賴性(表1)。Res低濃度(20-40 μmol/L)誘導處理,藥物作用24 h后其對B16F10細胞增值有促進作用;處理48和72 h對細胞增值生長沒有明顯影響。然而,較高濃度Res(80-200 μmol/L)作用下,處理24、48和72 h均對B16F10細胞生長有明顯的增值抑制作用,且隨著濃度的加大,誘導作用延長,增值抑制作用增加,抑制率最高達到68%(表2)。
表1 HCPT對B16F10細胞增殖抑制率(n=4,)

表1 HCPT對B16F10細胞增殖抑制率(n=4,)
注:*與對照組比較,P<0.05,下同
Concentration/(μmol·L-1)Inhibition rate/% 24 h 48 h 72 h 0 0 0 0 20 50.47 ± 3* 59.20 ± 5* 37.20 ± 4* 40 52.28 ± 2* 71.10 ± 5* 58.00 ± 3* 80 52.10 ± 2* 73.40 ± 3* 74.50 ± 3* 120 48.80 ± 2* 79.70 ± 2* 77.40 ± 4* 160 53.20 ± 3* 79.90 ± 2* 83.40 ± 4* 200 48.40 ± 6* 77.60 ± 2* 84.20 ± 1*
表2 Res對B16F10細胞增殖抑制率(n=4,)
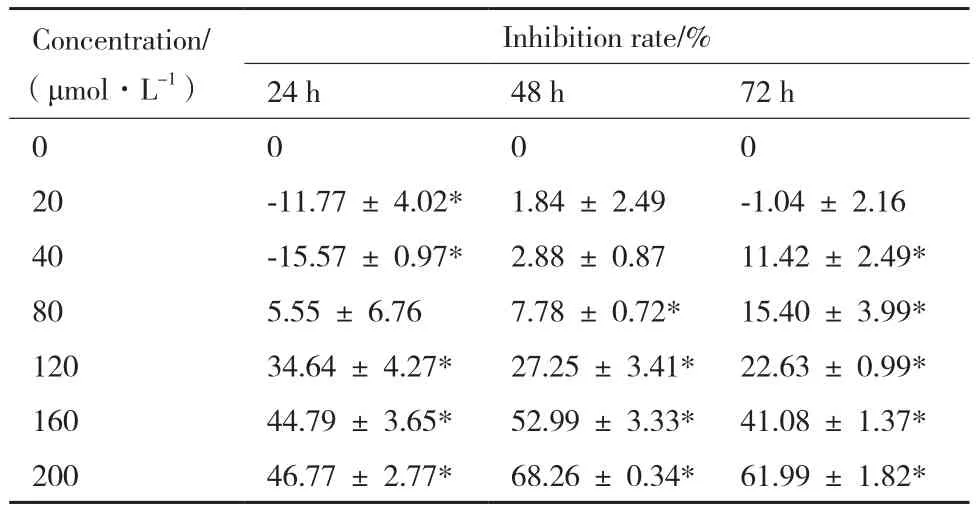
表2 Res對B16F10細胞增殖抑制率(n=4,)
Concentration/(μmol·L-1)Inhibition rate/% 24 h 48 h 72 h 0 0 0 0 20 -11.77 ± 4.02* 1.84 ± 2.49 -1.04 ± 2.16 40 -15.57 ± 0.97* 2.88 ± 0.87 11.42 ± 2.49* 80 5.55 ± 6.76 7.78 ± 0.72* 15.40 ± 3.99* 120 34.64 ± 4.27* 27.25 ± 3.41* 22.63 ± 0.99* 160 44.79 ± 3.65* 52.99 ± 3.33* 41.08 ± 1.37* 200 46.77 ± 2.77* 68.26 ± 0.34* 61.99 ± 1.82*
2.2 HCPT和Res誘導B16F10細胞凋亡
用倒置相差顯微鏡觀察藥物誘導B16F10細胞48 h的形態變化。對照組B16F10細胞形態飽滿,呈爪形或梭形,細胞膜完整,核區清晰,折光性好,增殖速度快(圖2-A,B,C)。40 μmol/L HCPT處理后,部分細胞出現降解,核區皺縮或膨大,細胞貼壁能力差,呈團塊狀生長,同時出現大量凋亡小體(圖2-D)。80和160 μmol/L HCPT處理組,細胞數量大幅度減少,大部分細胞細胞核皺縮,細胞解體成碎片或形成凋亡小體(圖2-E,F)。40 μmol/L Res處理組,多細胞未出現明顯的形態異常,仍保持良好的增殖狀(圖2-J),80 μmol/L Res處理組貼壁細胞未見明顯減少,部分細胞核區空泡化并逐漸解體,在細胞周圍出現顆粒狀的凋亡小體(圖2-H),160 μmol/L Res處理組只有少數細胞貼壁生長,大部分細胞完全解體為顆粒狀凋亡小體或形狀不規則碎片,部分細胞核區皺縮,且異常細胞聚集成團(圖2-I)。綜合觀察表明表現HCPT和Res對B16F10細胞具有明顯的凋亡誘導作用,其中80 μmol/L以上處理濃度的誘導作用尤為明顯,而且HCPT凋亡誘導作用明顯優于Res,40 μmol/L的濃度處理細胞已表現出一定的凋亡特征。
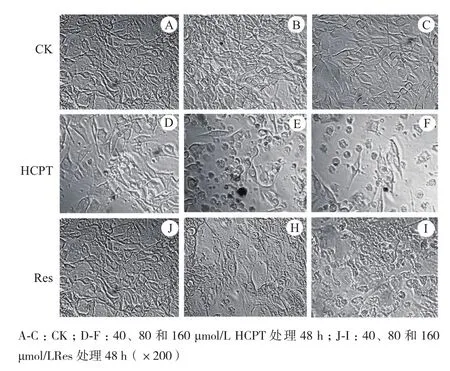
圖2 HCPT和Res對B16F10細胞顯微形態的影響
2.3 HCPT和Res抑制B16F10細胞黑色素的生成
與對照組相比,HCPT濃度在40和80 μmol/L時對B16F10細胞黑色素合成量具有顯著性抑制作用,黑色素生成量分別下降到87.6%和78.4%,呈現一定濃度依賴性;當HCPT濃度大于80 μmol/L后,黑色素合成量有持續下降趨勢,但濃度間差異不明顯,其中200 μmol/L處理組黑色素合成量為73.1%(圖3-A)。Res誘導作用下,B16F10細胞黑色素合成也受到顯著抑制,在20-200 μmol/L Res處理下,與對照組比較,細胞黑色素生成量下降到82%-88%,期間增加Res處理濃度,黑色素生成量雖有下降趨勢,但濃度間差異不明顯(圖3-B)。
2.4 HCPT和Res抑制B16F10細胞酪氨酸酶的活性
不同濃度HCPT處理對B16F10細胞中酪氨酸酶活性具有顯著性抑制作用,20-200 μmol/L間濃度處理下,隨著濃度的增大,抑制效果有逐漸增強趨勢,呈現一定濃度依賴性,其中200 μmol/L HCPT處理組,酪氨酸酶活性降低至71.8%(圖4-A)。不同濃度Res處理對B16F10細胞中酪氨酸酶活性也具有顯著性抑制作用,在20-200 μmol/L Res的誘導下,與對照組相比,酪氨酸酶活性下降到80%-90%,不同濃度處理間,沒有明顯濃度依賴性(圖4-B)。
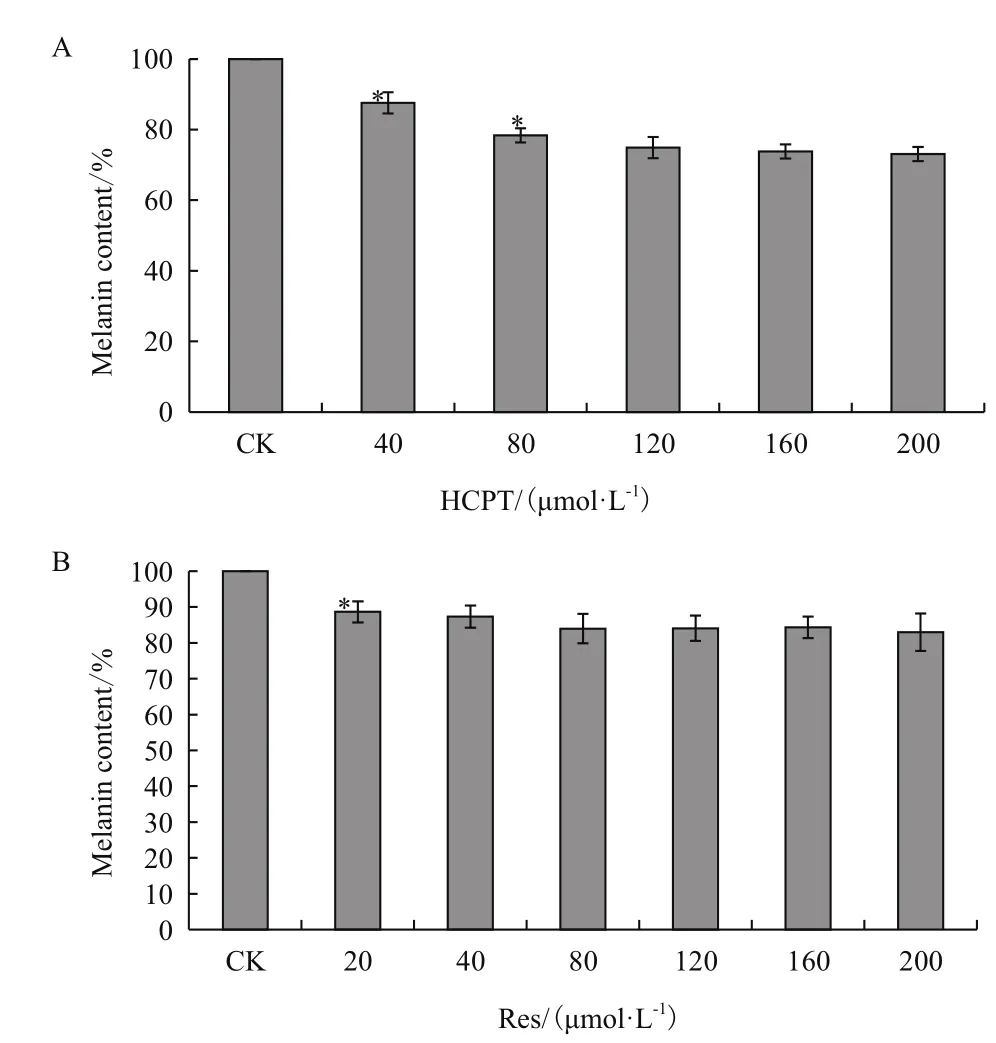
圖3 HCPT和Res對B16F10細胞黑色素合成的影響(n=4,)

圖4 HCPT和 Res對B16F10細胞酪氨酸酶活性的影響(n=4,)
2.5 HCPT和Res抑制B16F10細胞MITF和TYR基因的轉錄
在PCR反應中,目標產物的數量隨著循環數呈指數級增長。在反應過程中,隨著酶活力的下降及體系中各底物濃度的下降,反應速率逐漸變慢,最后反應進入一個平臺期,反應產物的濃度不再呈指數級增長。在達到平臺期之前,產物的相對濃度一定程度上反映了初始模版的相對濃度。因此,要首先確定內參基因β-actin以及目的基因TYR和MITF擴增過程中的反應平臺期。β-actin和TYR基因在31個循環時,擴增達到平臺期(圖5),因此本實驗選擇29個循環作為擴增循環數。MITF基因在35個循環時,擴增達到平臺期,我們選擇33個循環作為擴增循環數。

圖5 三基因片段不同循環數的擴增結果
利用Semi-RT-PCR實驗檢測HCPT誘導B16F10細胞凋亡TYR mRNA的表達量變化。β-actin mRNA的表達量在不同濃度HCPT誘導下保持穩定不變,說明該實驗過程中沒有其他因素干擾TYR mRNA的表達(圖6)。與對照組比較,HCPT(20、40、80、120和160 μmol/L)誘導細胞48 h,MITF和TYR mRNA表達受到顯著抑制,其中MITF mRNA表達抑制尤為顯著,TYR mRNA隨著HCPT濃度增加,表達量總體呈現降低趨勢。MITF蛋白是TYR基因的調控因子,結果表明HCPT可能通過抑制MITF mRNA表達下調TYR mRNA的水平,從而降低TYR活性,進而抑制細胞黑色素的生成。
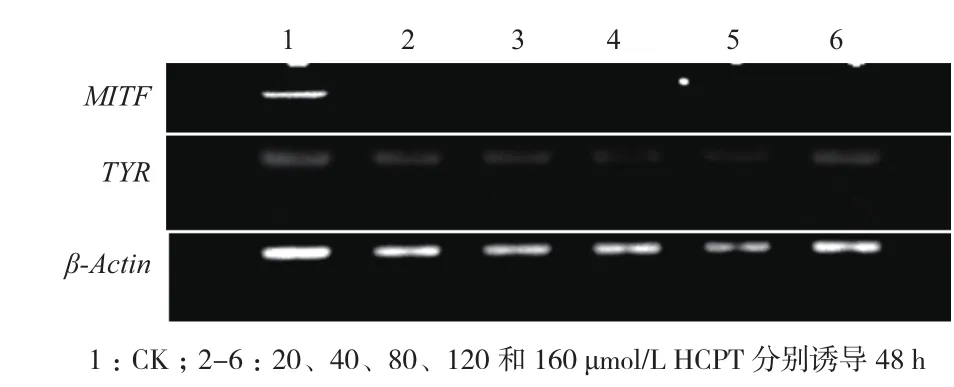
圖6 HCPT對B16F10細胞MITF and TYR mRNA相對表達影響
120和160 μmol/L Res分別誘導細胞48 h,MITF mRNA表達量明顯下降,低濃度(20、40和80 μmol/L)Res對細胞TYR mRNA表達沒有顯著影響(圖7),表明Res也可能通過抑制MITF mRNA表達下調TYR mRNA的水平,從而降低TYR活性,進而抑制細胞黑色素的生成。

圖7 Res對B16F10細胞TYR和MITF mRNA相對表達影響
3 討論
研究表明HCPT能夠通過干擾腫瘤細胞周期,促進腫瘤細胞凋亡,從而抑制其增殖生長[14,15]。HCPT具有抑制拓撲異構酶I的活性而使DNA斷裂的獨特機制。它能選擇性作用于Topo-I,形成穩定的“藥物-Topo-I-DNA復合物”。該復合物抑制最初由Topo-I介導的DNA裂解和重新連接反應,使處于細胞周期中S期DNA復制時形成的復制叉與已斷裂的DNA鏈沖突,會造成不可逆的復制叉阻滯、雙鏈DNA斷裂和可逆的可解離復合物轉變成不可逆的復合物,最終導致腫瘤細胞周期阻滯和細胞凋亡,表現出顯著的腫瘤活性[2]。Res對腫瘤也有確切的抑制作用,可作用于腫瘤的起始、促進、發展這3個階段[16],但其作用機制很復雜,尚未明確。Res可引起細胞生長抑制,細胞周期阻滯及細胞凋亡的生物學變化,而且變化程度與其濃度有關[17]。Res對腫瘤細胞增殖有雙相性,低濃度促進細胞增殖,高濃度抑制細胞增殖,其抑制作用呈量效關系[18]。同時,Res還具有抗腫瘤遷移的作用[19]。有研究報道,Res(25-100 μmol/L)可抑制黑色素腫瘤細胞增殖,引起其細胞周期改變[20]。本研究也發現HCPT和Res對小鼠黑色素瘤B16F10細胞均具有顯著的增殖抑制作用,同時藥物誘導下的細胞形態觀察發現兩種藥物對目標細胞具有顯著的凋亡誘導作用,因此這兩種藥物也很可能通過促進小鼠黑色素瘤B16F10細胞凋亡,抑制其增殖生長,表現出抗腫瘤活性。兩者的抗腫瘤具體分子機制仍不清楚,需深入研究。
惡性黑色素瘤是一種惡性程度較高的惡性腫瘤,其具有逃避凋亡、易轉移等重要特點。合成黑色素是黑素瘤細胞增殖生長過程的重要特征,其過程主要由黑素合成關鍵限速酶TYR基因家族調控,而TYR基因家族調節因子MITF可以上調酪氨酸酶基因表達,促進黑色素的生成[11-13]。本研究發現HCPT和Res可能通過抑制MITF mRNA表達下調TYR mRNA的水平,從而降低TYR活性,進而抑制細胞黑色素的生成。也有研究者發現乳酸菌發酵的豆奶提取物可通過抑制B16F0黑素細胞的TYR和MITF活性和表達,從而減少黑色素的合成[21]。該提取物抑制黑色素的作用機制與本研究的結果基本一致。然而,黑色素的合成過程也有酪氨酸酶基因家族中的酪氨酸相關蛋白1(TRP-1)和酪氨酸相關蛋白2(TRP-2)的參與,Res對這兩種相關蛋白是否有影響有待研究。
4 結論
本研究發現HCPT和Res對B16F10細胞增殖都有抑制作用,且隨著濃度增加,抑制作用增強。HCPT和Res可能通過誘導細胞凋亡抑制B16F10細胞的增殖,同時通過下調MITF基因轉錄,抑制TYR mRNA的表達及TYR酶活性,進而抑制細胞黑色素的生成。
[1]袁丹, 容如濱, 喻宗沅等. 抗腫瘤藥物10-羥基喜樹堿的研究與應用進展[J]. 化學與生物工程, 2007, 12:9-11.
[2]Meng GR, Li JJ, Wang GL, et al. Synthesis and cytotoxic activities of the amino acid-conjugates of 10-hydroxycamptothecin[J]. Chinese Journal of Organic Chemistry, 2014, 1:155-160.
[3]代魯平, 宋春霞, 邵先祥, 等. 抗癌藥物喜樹堿類衍生物的研究進展[J]. 中國藥學雜志, 2010, 23:1813-1815.
[4]王征, 羅澤民, 鄧林偉. 白藜蘆醇的藥理作用機理和合成途徑[J]. 天然產物研究與開發, 2003, 15(2):178-181.
[5]劉宏勝, 王金環, 徐新女, 等. 白藜蘆醇對U251人腦膠質瘤細胞生長的抑制及相關機制的研究[J]. 中國中藥雜志, 2009, 34(8):1027-1031.
[6]盧晨欣, 孫警輝, 伍春蓮. 白藜蘆醇與紫杉醇聯合用藥對人喉癌Hep-2細胞凋亡機制的研究[J]. 中國中藥雜志, 2016, 41(3):476-483.
[7]呂莎, 李哲, 王顯江, 等. 靶向NRP1單克隆抗體對黑色素瘤細胞B16F10的增殖、遷移和凋亡的影響[J]. 中國藥學雜志, 2014, 49(20):1806-1812.
[8]曾紅梅, 張思維, 鄭榮壽, 等. 2003~2007年中國皮膚黑色素瘤發病與死亡分析[J]. 中國腫瘤, 2012, 3:183-189.
[9]唐智柳, 石建偉, 蔡美玉, 等. 中國皮膚黑色素瘤發病及其影響因素分析[J]. 中國腫瘤, 2014, 10:829-833.
[10]楊嫻, 劉汝清, 杜德榮, 等. 白花蛇舌草總黃酮對黑色素瘤a375和b16細胞增殖抑制作用的研究[J]. 世界中西醫結合雜志, 2011, 10:844-846, 856.
[11]Vance KW, Goding CR. The transcription network regulating melanocyte development and melanoma[J]. Pigment Cell Research, 2004, 17(4):318- 325.
[12]Steingrimsson E, Copeland NG, Jenkins NA. Melanocytes and the microphthalmia-transcription factor network[J]. Annual Review of Genetics, 2004, 38:365-411.
[13]Levy C, Khaled M, Fisher DE. M ITF:master regulator of melanocyte development and melanoma oncogene[J]. Trends in Molecular Medicine, 2006, 12:406- 414.
[14]胡文晉, 胡巍, 方蕓. 喜樹堿類藥物抗腫瘤機制研究進展[J].中國藥房, 2012, 27:2581-2584.
[15]張超, 胡巍, 方蕓. 霧化吸入羥基喜樹堿對小鼠B16黑色素瘤實驗性肺轉移的影響[J]. 中國中藥雜志, 2011, 36(5):618-623.
[16]Shankar S, Singh G, Srivastava RK. Chemoprevention by resveratrol:molecular mechanisms and therapeutic potential[J]. Frontiers in Bioscience, 2007, 12:4839- 4854.
[17]Joe AK, Liu H, Masumi S, et al. Resveratrol induces growth inhibition, S-phase arrest, apoptosis, and changes in biomarker expression in several human cancer cell lines[J]. Clinical Cancer Research, 2002, 8:893-903.
[18]陳家順, 陶方方, 侯敏, 等. 白藜蘆醇對鼠纖維肉瘤S180細胞周期及其機制研究[J]. 南京醫科大學學報, 2008, 28(5):570-574.
[19]Bishayee A. Cancer prevention and treatment with resveratrol:from rodent studies to clinical trials[J]. Cancer prevention research(Philadelphia, Pa. ), 2009, 2(5):409-418.
[20]孟德萍, 鄒志孟, 許曉群, 等. 白藜蘆醇對黑色素瘤細胞增殖的抑制作用[J]. 中華腫瘤防治雜志, 2012, 19(5):328-331.
[21]Chen YM, Shih TW, Chiu CP, et a1. Effects of lactic acid bacteriafermented soy milk on mela nogenesis in B16F0 melanocytes[J]. Journal of Functional Foods, 2012, 5:395-405.
(責任編輯 李楠)
Study on Inhibition Mechanism of Melanogenesis and Melanin Synthesis of B16F10 Cell Induced by Two Natural Products
CHENG Xing-an1ZHANG Shu-ming1ZHOU Xiao-wu1WU Bo1LIN Xian-wei1QIN Xiang-jing2HUANG Su-qing1LIU Zhan-mei1JIANG Xu-hong1
(1. Institute of Natural Product Chemistry,Zhongkai University of Agriculture and Engineering,Guangzhou 510225;2. South China Sea Institute of Oceanology,Chinese Academy of Sciences,Guangzhou 510301)
This study aims to clarify the inhibition mechanism of proliferation and melanin synthesis of B16F10 cell induced by 10-hydroxycamptothecin(HCPT)and resveratrol(Res). MTT cytotoxicity assay,microscope observation-assay,L-Dopa oxidation-assay and NaOH cracking-assay were employed to investigate the proliferation,cellular morphology,tyrosinase(TYR)activity and melanin synthesis of B16F10 cells treated by HCPT and Res at different concentrations,respectively;and semi-RT-PCR to analyze the relative mRNA expression of TYR and microphthalmia associated transcription factor(MITF)in B16F10 cells. It was found that HCPT(40,80,120,160,and 200 μmol/L)and Res(80,120,160,and 200 μmol/L)inhibited B16F10 cell proliferation through inducing apoptosis,and exhibited significantly dose-dependent inhibiting effects on tyrosinase activity and melanin synthesis on B16F10 cell(P < 0.05). In addition,relative mRNA expressions of the TYR and MITF genes in B16F10 cells were depressed remarkably by HCPT at different concentrations and Res at high concentrations(120 and 160 μmol/L). Conclusively,HCPT and Res may inhibit the B16F10 cell proliferation via inducing cell apoptosis,and depress the melanin synthesis in B16F10 cells from down-regulating the transcription of MITF gene and subsequently inhibiting the gene expression of TYR mRNA and activity of TYR.
10-hydroxycamptothecin;resveratrol;tyrosinase;melanin;microphthalmia associated transcription factor
10.13560/j.cnki.biotech.bull.1985.2017-0122
2017-02-22
國家自然科學基金項目(21406274,21376281),廣東省大學生創新訓練項目(201411347034,201511347056)
程杏安,男,博士,副教授,研究方向:天然產物化學;E-mail:anzai_28@163.com
劉展眉,教授,碩士生導師,E-mail:liuzhanm@21cn.com;蔣旭紅,教授,E-mail:jiangxh69@163.com
