亞麻快速生長期細胞壁形成相關(guān)基因的表達分析
2017-07-19 13:17:53江海霞郭棟良李玉環(huán)閆文亮楊亮杰謝麗瓊
中國農(nóng)業(yè)科學(xué) 2017年13期
關(guān)鍵詞:研究
江海霞,郭棟良,李玉環(huán),閆文亮,楊亮杰,謝麗瓊
?
亞麻快速生長期細胞壁形成相關(guān)基因的表達分析
江海霞,郭棟良,李玉環(huán),閆文亮,楊亮杰,謝麗瓊
(新疆大學(xué)生命科學(xué)與技術(shù)學(xué)院,烏魯木齊 830046)
【目的】亞麻在快速生長期其韌皮纖維細胞發(fā)育在SP(the snap point)點上下端分別經(jīng)歷細胞伸長和次生細胞壁加厚2個不重疊時期。研究亞麻快速生長期不同組織、不同時期、不同器官中與細胞壁形成相關(guān)的β-半乳糖苷酶(LuBGALs)、纖維素合酶(LuCESAs)等家族基因的表達譜,探討快速生長期亞麻韌皮纖維細胞細胞壁的發(fā)育模式,為改善亞麻纖維產(chǎn)量提供理論依據(jù)。【方法】以生長45 d的亞麻根、莖韌皮纖維、葉為材料,用透射電鏡觀察并測量莖韌皮纖維細胞細胞壁結(jié)構(gòu)和厚度,采用實時熒光定量(qRT-PCR)方法,研究亞麻快速生長期和等細胞壁形成相關(guān)的基因在亞麻韌皮纖維不同階段的表達特點。【結(jié)果】在SP點上部TOP端纖維細胞細胞壁薄,約110 nm;緊鄰SP點莖中部的MID區(qū)(約500 nm)和莖下部的BOT區(qū)(約650 nm),細胞壁厚度明顯增厚,細胞壁質(zhì)地均一,沒有明顯的分層現(xiàn)象,說明SP點中部和下部韌皮纖維細胞細胞壁已經(jīng)開始加厚但還未進入次生壁加厚階段,與TOP明顯不同。亞麻在TOP區(qū)的表達顯著低于MID區(qū)和BOT區(qū),表明其主要參與纖維細胞細胞壁加厚過程。而、、在TOP區(qū)表達最高,MID區(qū)次之,表明此類基因主要參與亞麻韌皮細胞伸長和細胞壁重建過程。在幼嫩的TOP區(qū)表達量高,在亞麻莖較為成熟的MID區(qū)較低,說明在細胞壁形成過程中起作用。其他基因的表達量均較低。在亞麻莖幼嫩的TOP區(qū)纖維細胞中,亞麻纖維素合酶基因、、、、和都檢測出較高的表達量,且明顯高于其在MID區(qū)和BOT區(qū)的表達。其中和在MID區(qū)的表達顯著低于BOT區(qū),其他幾個基因在MID和BOT的表達并無明顯差異。結(jié)合這些基因在亞麻快速生長期不同器官中的表達模式,結(jié)果說明,亞麻中6個(、、、、和)主要促進亞麻韌皮纖維細胞的伸長。在幼莖韌皮纖維細胞中表達量高,表明亞麻莖伸長和加粗需要大量能量。在亞麻細胞壁發(fā)育過程中發(fā)揮作用。【結(jié)論】快速生長期亞麻莖韌皮纖維細胞細胞壁沒有次生加厚過程;、、、、、、和在亞麻細胞壁細胞伸長過程中起作用;主要促進亞麻細胞壁加厚過程;和在亞麻細胞壁發(fā)育中發(fā)揮作用。
亞麻;韌皮纖維;細胞壁形成;基因相對表達量
0 引言
【研究意義】亞麻(L.)是亞麻科亞麻屬一年生草本植物,其纖維來源于成熟亞麻莖的韌皮纖維。亞麻韌皮纖維細胞起源于單個細胞,其發(fā)育在形態(tài)學(xué)上經(jīng)歷了細胞伸長和次生細胞壁加厚2個不重疊時期,這兩個階段被主莖上的分界點(the snap point,SP)分開。植株進入快速生長期后,SP分界點之上為韌皮細胞伸長階段,之下為細胞壁加厚階段[1]。這兩個階段分別決定了成熟亞麻韌皮纖維的長度和厚度,最終影響亞麻纖維的產(chǎn)量和品質(zhì)。因此,研究與細胞壁形成相關(guān)基因在亞麻SP點不同發(fā)育階段細胞中的表達模式對進一步理解亞麻纖維細胞形成、細胞壁加厚的發(fā)育規(guī)律具有重要意義。【前人研究進展】β-半乳糖苷酶家族(BGALs),纖維素合成酶家族(CESAs)等在植物細胞壁形成和重建過程中起重要作用。BGAL屬于糖苷水解酶家族成員(glycosyl hydrolyse family 35,GH35),參與細胞壁纖維素形成[2-3]和細胞壁細胞伸長[4]等過程。擬南芥中有17個β-半乳糖苷酶基因,其中,已證實、、、、和在細胞壁重建和擴展中起作用[5]。β-半乳糖苷酶還與棉花纖維細胞的起始和分化相關(guān)[6]。亞麻中預(yù)測有35個,芯片和qRT-PCR方法檢測發(fā)現(xiàn)部分在纖維中表達[7]。纖維素是植物細胞壁中最重要的組分,由位于質(zhì)膜上的纖維素合酶復(fù)合體(cellulose synthase complexs,CSCs)合成。擬南芥、楊樹、棉花、豌豆、水稻等多種植物中研究發(fā)現(xiàn)纖維素合酶是多基因家族,不同組分形成不同CSC復(fù)合體,在初生細胞壁和次生細胞壁形成過程中分別起作用,作用模式現(xiàn)已基本研究清楚。擬南芥CESA1、CESA3、CESA6蛋白形成CSC參與初生壁纖維素合成[8],缺失其中任一組分,植株致死;而、、決定次生壁纖維素合成[9],其他基因在不同生理時期和組織中參與細胞壁形成。棉花中至少有15個基因是纖維素合成所必需的[10],其中、、、主要參與次生細胞壁的合成,、、、在棉花初生細胞壁的合成中起作用[11]。已有研究表明楊樹中有18個基因,其中5個基因(、-A、-B、-A、-B)主要在次生細胞壁合成中發(fā)揮作用[12-16],其他在楊樹不同生長發(fā)育過程中參與細胞壁構(gòu)建過程[17-19]。通過病毒介導(dǎo)的基因沉默(VIGS)法,研究者發(fā)現(xiàn)、、、、在亞麻纖維細胞壁構(gòu)建中扮演重要角色[20]。和在亞麻快速生長期、花期和綠熟期的表達高,推測這兩個基因在亞麻次生細胞壁加厚過程中起重要作用[21]。【本研究切入點】盡管已有研究分析了亞麻不同組織器官的基因表達譜,但各個研究者關(guān)于韌皮纖維發(fā)育的芯片數(shù)據(jù)結(jié)果并不一致,亞麻快速生長期與纖維合成密切相關(guān)的、等基因的表達模式還不清楚[20,22-24]。因此,和等基因家族在亞麻韌皮纖維細胞中的表達模式需進一步分析明確。【擬解決的關(guān)鍵問題】本研究通過qRT-PCR方法,擬探討與細胞壁形成相關(guān)的和等基因在亞麻快速生長期韌皮纖維細胞伸長和細胞壁加厚過程中的表達模式,為研究纖用亞麻韌皮纖維發(fā)育機制奠定理論基礎(chǔ)。
1 材料與方法
試驗于2016年在新疆大學(xué)生命科學(xué)與技術(shù)學(xué)院植物培養(yǎng)室完成。
1.1 材料
亞麻(L.)品種范妮(FANY)。范妮亞麻種子種植在花土﹕蛭石=3﹕1的土壤中,置于光照強度0—10 000 lx,16 h光照/8 h黑暗,(25±2)℃溫室中培養(yǎng)45 d。
1.2 亞麻總RNA的提取和cDNA的合成
種植后21—50 d為亞麻快速生長期,45 d的亞麻處于快速生長期后期,莖的表皮和木質(zhì)部較易分離。取生長45 d亞麻莖3段,每段約2 cm。最上端(TOP)距莖尖1—3 cm且位于分界點(SP)上部。第二段(MID)距莖尖4—6 cm。基部(BOT)距莖尖17—19 cm。TOP直接取莖,MID和BOT取莖的表皮(圖1-A)。每9棵亞麻莖段為一個池,每個部位設(shè)置6個重復(fù),共18個樣品。所取亞麻器官還包括生長了45 d的亞麻幼葉、成熟葉、根和基部,每個器官設(shè)置4個生物學(xué)重復(fù),共16個樣品。
參照TIANGEN RNAprep Pure 植物總RNA提取試劑盒說明書提取總RNA,使用Thermo超微量分光光度計進行RNA定量。參照TaKaRa第一鏈合成試劑盒說明書合成cDNA。
1.3 引物設(shè)計
根據(jù)已知擬南芥相關(guān)基因序列,通過Phytozome v9.1和Quant Prime qPCR在線工具設(shè)計qRT-PCR引物。引物如電子版附表1所示。
1.4 亞麻莖透射電鏡TEM觀察
亞麻莖TOP、MID、BOT樣品經(jīng)過固定液(2.5%戊二醛-磷酸緩沖液)固定、梯度乙醇脫水、包埋、固化后切片,切片經(jīng)3%醋酸鈾-枸櫞酸鉛雙染色后在透射電鏡下觀察拍照。每個部位設(shè)置3個生物學(xué)重復(fù)。
1.5 實時熒光定量PCR(qRT-PCR)檢測基因的相對表達量
以合成的cDNA為模板,(eukaryotic translation initiation factor 1)為內(nèi)參基因[7],使用ABI PRISM 7500實時熒光定量PCR儀進行qRT-PCR 試驗,每個樣品設(shè)置3個重復(fù)。反應(yīng)體系為20 μL;包括1 μL cDNA、10 μL iTaqTMUniversal SYBR?Green Supermix、上下游引物各0.5 μL和8 μL RNase free H2O。PCR反應(yīng)條件為95℃2 min;95℃15 s,60℃20 s,72℃15 s,40個循環(huán)。
相對表達水平數(shù)據(jù)采用2-ΔΔct法進行分析。用GraphPad Prism 5和SPSS 19軟件對目的基因的相對表達量進行統(tǒng)計學(xué)分析。用△CT法對相對轉(zhuǎn)錄水平進行分析,用MeV 4.9.0(Multi Experiment Viewer,http://www.tm4.org/)作圖。
2 結(jié)果
2.1 亞麻莖不同部位韌皮纖維細胞細胞壁的發(fā)育
亞麻進入快速生長期后,株高呈現(xiàn)明顯的日變化。為了探討亞麻莖韌皮纖維細胞細胞壁在快速生長期的發(fā)育變化,研究檢測了亞麻莖SP點上下不同部位細胞壁發(fā)育的情況。結(jié)果表明,SP點上部,靠近莖尖生長點,莖部組織分化不明顯,莖的表皮與髓部無法分離。SP點下部靠近莖中部區(qū)域,細胞壁開始加厚,莖表皮能夠與木質(zhì)部分離,越往莖的基部,亞麻莖表皮越易剝離,韌皮纖維也越厚(圖1-A)。透射電鏡觀察亞麻韌皮纖維在快速生長期細胞壁的厚度變化(圖1-B),結(jié)果表明,在SP點上部TOP端纖維細胞可見結(jié)構(gòu)疏松的細胞壁物質(zhì),壁薄約110 nm;緊鄰SP點下部的MID區(qū),細胞壁厚度明顯增厚,約500 nm,壁物質(zhì)密度增大;靠近莖中下部的BOT區(qū),細胞壁厚度增大到約650 nm(表1)。盡管中下部細胞壁厚度增加,從圖中可看出,細胞壁質(zhì)地均一,沒有明顯的分層現(xiàn)象。
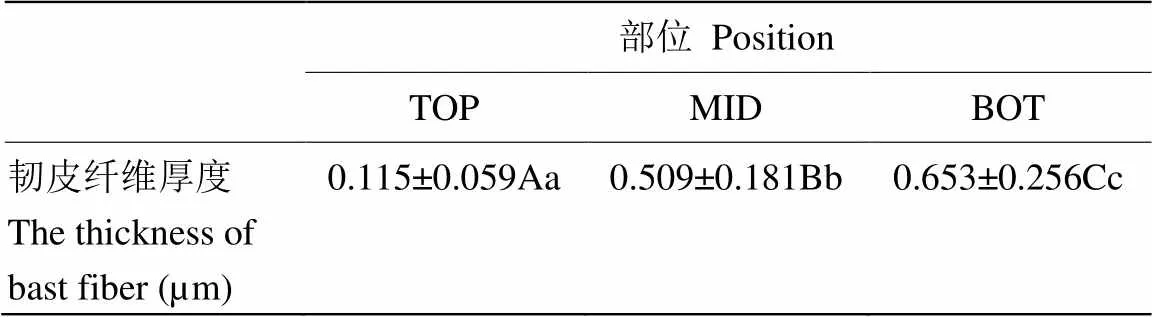
表1 亞麻莖在SP點不同區(qū)域韌皮纖維厚度
平均值±SD,N≥30。大寫字母和小寫字母分別表示在<0.01和<0.05水平差異顯著
Average ± SD, N≥30. Uppercase and lowercase letters indicate significance at<0.01 and<0.05, respectively
2.2在亞麻快速生長期不同器官和莖韌皮纖維的表達分析
為了獲得β-半乳糖苷酶家族基因在亞麻快速生長期細胞壁形成中的作用,研究分析了這些基因在亞麻莖SP點不同部位韌皮纖維細胞和根、莖、葉不同器官中的表達水平(圖2)。
2.2.1在亞麻韌皮纖維中的表達分析 圖2-A和2-C顯示了SP點上下不同部位表達有差異的。結(jié)果表明,在亞麻莖的3個不同部位的表達量存在極顯著差異(<0.01),TOP區(qū)的表達顯著低于MID區(qū)和BOT區(qū)(<0.01),表明它主要促進韌皮纖維細胞壁加厚過程。而、、在TOP區(qū)表達最高,MID區(qū)次之,表明此類基因主要參與亞麻韌皮細胞伸長和細胞壁形成過程。在幼嫩的TOP區(qū)表達量高,麻莖較為成熟的MID區(qū)較低(<0.05)說明主要參與細胞壁細胞伸長過程。、在亞麻莖的表達都較低,但在TOP區(qū)的表達極顯著高于MID和BOT區(qū)(<0.01),說明這兩個基因可能在亞麻韌皮纖維細胞伸長過程中起作用。、、、在莖韌皮纖維中都具有較高的表達水平,說明在細胞壁加厚過程中多個共同作用。
2.2.2在亞麻不同器官中的表達分析 為了進一步分析基因家族的表達模式,研究檢測了基因在亞麻根、莖、葉3個不同器官中的表達情況(圖2-B和2-D)。結(jié)果顯示和在幼葉中、成熟葉和根中的表達極顯著低于BOT區(qū)(<0.001)。在幼葉和成熟葉中的表達極顯著高于BOT區(qū)(<0.001),但在根中的表達極顯著低于BOT區(qū)(<0.001)。和具有相似的表達模式,在幼葉中的表達顯著高于BOT(<0.001),根和成熟葉的表達量顯著低于BOT(<0.01)。和在幼葉、成熟葉、根中的表達顯著高于對照(<0.05)。
盡管BGALs家族基因在亞麻中預(yù)測有30多個成員,但多數(shù)在麻莖韌皮纖維的生長發(fā)育過程中處于不表達狀態(tài)(數(shù)據(jù)未顯示)。在BOT和MID的表達較高,而在根中表達較低;在幼莖、幼葉和成熟葉中的表達量高,而在根中的表達低;、在幼嫩部位(TOP和YL)的表達高;在所有組織的表達都較高;和在所有研究組織中的表達都較低。說明BGALs家族基因的表達具有明顯的組織器官差異。
2.3在亞麻快速生長期不同器官和莖韌皮纖維中的表達分析
擬南芥、楊樹、棉花等植物中基因有多個組合,分別在不同的細胞壁發(fā)育階段起作用[8, 11-19]。研究探討了該家族基因在亞麻快速生長期細胞壁形成過程中的表達模式(圖3)。
2.3.1在亞麻韌皮纖維中的表達分析 圖3-A和圖3-C結(jié)果顯示在麻莖幼嫩的TOP區(qū)纖維細胞中,、、、、和都檢測出較高的表達量,且明顯高于其在MID區(qū)和BOT區(qū)的表達(<0.001)。其中和在MID區(qū)的表達顯著低于BOT區(qū)(<0.05),其他幾個基因在MID和BOT的表達并無明顯差異。上述結(jié)果說明亞麻中6個基因(、、、、和)主要促進亞麻韌皮纖維細胞的伸長。在TOP和MID的表達極顯著高于BOT區(qū)(<0.01),而在TOP和MID的表達無顯著性差異。在莖不同部位的表達無顯著性差異。和不僅在亞麻TOP區(qū)的表達較高,在BOT和MID區(qū)也具有較高的表達,表明這兩個基因也參與細胞壁加厚過程。
2.3.2在亞麻不同器官中的表達分析 圖3-B和3-D表明在纖維素合酶基因家族中,除了和,其余的成員都在幼葉中有顯著高表達(<0.01)。、、和等基因在根中的表達較低,達到極顯著水平(<0.001),而在根中的表達與莖BOT區(qū)中的表達沒有顯著差異,而在幼葉和成熟葉中的表達極顯著低于根和莖BOT區(qū)(<0.001)。
LuCESA1、LuCESA3、LuCESA9和LuCESA10在莖和葉中都表達,在TOP和YL的表達更高。結(jié)合纖維素合成酶復(fù)合體的模式,LuCESA1、LuCESA3、LuCESA9和LuCESA10可能參與初生壁的形成。
2.4和在快速生長期不同組織和莖韌皮纖維中的表達分析
蔗糖合酶(SuSy)和木聚糖水解酶(XTH)在細胞壁形成與加厚過程中起作用。本研究檢測了和的表達情況(圖4)。結(jié)果顯示在亞麻幼嫩組織中相對表達量較高(圖4-A),在莖的TOP區(qū)和幼葉中的表達顯著高于其他部位。在莖、葉和根中的表達都較低(圖4-B),盡管在亞麻麻莖MID區(qū)的表達顯著高于TOP區(qū)(<0.05,圖4-C),但在幼葉和根中明顯比在莖中活躍(圖4-D)。表明和在亞麻細胞壁細胞形成和增大過程中起作用。
盡管細胞壁形成相關(guān)基因、、、、和在不同部位和器官中的表達量較低,但其表達具有顯著性差異,研究認為其在亞麻纖維發(fā)育表達水平上仍有一定的生物學(xué)意義。
3 討論
亞麻是研究韌皮纖維細胞發(fā)育理想的模式植物[25]。本研究通過觀察亞麻莖纖維細胞細胞壁的發(fā)育,發(fā)現(xiàn)亞麻莖SP點基部和中部在細胞壁形態(tài)上沒有明顯區(qū)別,只有細胞壁的加厚,沒有出現(xiàn)明顯的分層,說明此時細胞還未進入次生細胞壁加厚階段。國內(nèi)研究認為亞麻纖維細胞壁加厚階段在現(xiàn)蕾期到綠熟期[26-27],而Gorshkova等[28]認為快速生長期伴隨次生細胞壁加厚過程,本文的結(jié)果表明至少在快速生長期后期,仍未出現(xiàn)次生壁加厚情況。
β-半乳糖苷酶是糖苷水解酶家族成員,廣泛分布于植物組織中,在植物果實生長成熟[29-30]過程中研究較多[31]。BGALs家族基因在亞麻中預(yù)測有30多個成員,多數(shù)在亞麻細胞壁發(fā)育過程中的功能并不清楚[7]。在亞麻莖韌皮纖維細胞中,發(fā)現(xiàn)多個在幼嫩的莖中高表達。結(jié)合這些基因在幼葉、成熟葉、根中的表達(圖2-D)情況,認為在亞麻快速生長期,主要在SP點下部表達,在全株的表達較高,主要在幼葉和幼莖(SP點上部)起作用,與前人研究結(jié)果一致[7]。不僅在莖中表達,在葉中表達更高,在幼莖、幼葉和根中有較高的表達量,和在開花前各部位表達都較低,與NEIL HOBSON[7]的結(jié)果有差異。前人根據(jù)BGAL蛋白序列將LuBGAL1、LuBGAL3、LuBGAL5、LuBGAL6和LuBGAL9劃分在A1族[7],認為這些半乳糖苷酶在亞麻韌皮纖維發(fā)育中有重要作用,本研究證實了這個推論。
纖維素合酶基因在亞麻中的研究主要從2012年亞麻全基因組測序完成后開始,通過擬南芥、楊樹等植物中的同源序列分析,推測亞麻中有10—16個基因可能參與不同階段細胞壁合成過程。本研究結(jié)果顯示,、、、主要促進韌皮纖維細胞初生細胞壁的形成,與前人報道相同[32]。已有研究報道、和在木質(zhì)部中參與亞麻次生細胞壁的合成[23,32],亞麻外莖組織中,、和在亞麻韌皮纖維細胞的表達模式與前人結(jié)果一致[23]。結(jié)合三者的表達情況和莖的橫切,認為亞麻快速生長期的后期,盡管細胞壁沒有明顯分層,但與次生壁相關(guān)的基因已經(jīng)開始表達。、在CESA家族中處于同一亞族,親緣關(guān)系非常近[21, 23, 32],由于在檢測的組織中它們的表達量都較低,因此認為、可能在亞麻韌皮纖維細胞發(fā)育過程中出現(xiàn)功能冗余。
在植物細胞形成和增大過程中,蔗糖合酶(SuSy)為細胞壁纖維素合成提供底物UDPG,糖苷水解酶家族GH16中的木聚糖水解酶(XTH)[33]催化木葡聚糖分子的斷裂和重新連接,參與細胞壁的合成或松弛過程,二者在初生壁和次生壁的合成中起重要作用。在亞麻幼嫩組織中相對表達量較高說明在新生細胞中提供底物;同時,在麻莖中的表達量高于成熟葉和根,說明在亞麻快速生長期,莖是重要的代謝庫,需要更多的UDPG參與到韌皮纖維細胞壁加厚的過程。盡管在亞麻麻莖MID區(qū)的表達顯著高于TOP區(qū)(<0.05),但在幼葉中表達量更高,說明可能主要在幼葉中參與細胞壁的形成。和的表達與莖發(fā)育變化呈現(xiàn)一致性。
前人對亞麻韌皮纖維發(fā)育的研究,在研究材料、取材時期和取材部位上多有不同[20,22-23],不同研究者的結(jié)果存在差異。研究同一時期,同一植株的不同部位細胞壁形成相關(guān)基因的表達,為亞麻快速生長期韌皮纖維細胞的發(fā)育提供基本的理論支持。
4 結(jié)論
4.1 快速生長期亞麻莖韌皮纖維細胞厚度TOP<MID<BOT區(qū),SP點下部細胞壁沒有次生加厚過程。
4.2 多個共同參與亞麻細胞壁的形成,其中、、和主要促進亞麻細胞壁細胞伸長,主要參與亞麻細胞壁加厚過程。
4.3 LuCESA1、LuCESA3、LuCESA9、LuCESA10參與初生壁的形成。
4.4和的表達與莖發(fā)育變化呈現(xiàn)一致性。
References
[1] GORSHKOVA T A, AGEEVA M, CHEMIKOSOVA S, SALNIKOV V. Tissue-specific processes during cell wall formation in flax fiber., 2005, 139(1): 88-92.
[2] ROACH M J, MOKSHINA N Y, BADHAN A, SNEGIREVA A V, HOBSON N, DEYHOLOS M K, GORSHKOVA T A. Development of cellulosic secondary walls in flax fibers requires β-galactosidase., 2011, 156(3): 1351-1363.
[3] MOKSHINA N E, IBRAGIMOVA N N, SALNIKOV V V, AMENITSKII S I, GORSHKOVA T A. Galactosidase of plant fibers with gelatinous cell wall: Identification and localization., 2012, 59(2): 246-254.
[4] SAMPEDRO J, GIANZO C, IGLESIAS N, GUITI N E, REVILLA G, ZARRA I. AtBGAL10 is the main xyloglucan β-galactosidase in, and its absence results in unusual xyloglucan subunits and growth defects., 2012, 158(3): 1146-1157.
[5] CHANDRASEKAR B, RA V D H. Beta galactosidases inand tomato-a mini review., 2016, 44(1): 150-158.
[6] ZHAO G R, LIU J Y, XIONGMING D U. Molecular cloning and characterization of cotton cDNAs expressed in developing fiber cells., 2001, 65(12): 2789-2793.
[7] HOBSON N, DEYHOLOS M K. Genomic and expression analysis of the flax () family of glycosyl hydrolase 35 genes., 2013, 14(1): 207-210.
[8] ROBERT S, MOUILLE G, H FTE H. The mechanism and regulation of cellulose synthesis in primary walls: Lessons from cellulose-deficientmutants., 2004, 11(3): 351-364.
[9] TAYLOR N G. Cellulose synthesis in thesecondary cell wall., 2004, 11(3): 329-338.
[10] FANG L, TIAN R, CHEN J, WANG S, LI X, WANG P, ZHANG T. Transcriptomic analysis of fiber strength in upland cotton chromosome introgression lines carrying differentchromosomal segments., 2014, 9(4): 220-226.
[11] LI A, XIA T, XU W, CHEN T, LI X, FAN J, WANG R, FENG S, WANG Y, WANG B, PENG L. An integrative analysis of four CESA isoforms specific for fiber cellulose production betweenand., 2013, 237(6): 1585-1597.
[12] HAI G, JIA Z, XU W, WANG C, CAO S, LIU J, CHENG Y. Characterization of the Populus PtrCesA4 promoter in transgenic×., 2016, 124(3): 495-505.
[13] DJERBI S, LINDSKOG M, ARVESTAD L, STERKY F, TEERI T T. The genome sequence of black cottonwood () reveals 18 conserved cellulose synthase (CesA) genes., 2005, 221(5): 739-746.
[14] KUMAR M, THAMMANNAGOWDA S, BULONE V, CHIANG V, HAN K H, JOSHI C P, MANSFIELD S D, MELLEROWICZ E, SUNDBERG B, TEERI T. An update on the nomenclature for the cellulose synthase genes in Populus., 2009, 14(5): 248-254.
[15] SONG D, SHEN J, LI L. Characterization of cellulose synthase complexes in Populus xylem differentiation., 2010, 187(3): 777-790.
[16] SUZUKI S, LI L G, SUN Y H, CHIANG V L. The cellulose synthase gene superfamily and biochemical functions of xylem-specific cellulose synthase-like genes in., 2006, 142(3): 1233-1245.
[17] WU L, JOSHI C P, CHIANG V L. A xylem-specific cellulose synthase gene from aspen () is responsive to mechanical stress., 2000, 22(6): 495-502.
[18] SAMUGA A, JOSHI C P. A new cellulose synthase gene (PtrCesA2) from aspen xylem is orthologous toAtCesA7 (irx3) gene associated with secondary cell wall synthesis., 2002, 296(1/2): 37-44.
[19] KALLURI U C, JOSHI C P. Isolation and characterization of a new, full-length cellulose synthase cDNA, PtrCesA5 from developing xylem of aspen trees., 2003, 54(390): 2187-2188.
[20] CHANTREAU M, CHABBERT B, BILLIARD S, HAWKINS S, NEUTELINGS G. Functional analyses of cellulose synthase genes in flax () by virus-induced gene silencing., 2015, 13(9): 1312-1324.
[21] 袁紅梅, 郭文棟, 趙麗娟, 于瑩, 吳建忠, 張利國, 程莉莉, 趙東升, 吳廣文, 關(guān)鳳芝. 亞麻纖維素合酶超基因家族的生物信息學(xué)及表達分析. 中國農(nóng)業(yè)科學(xué), 2016, 49(23): 4656-4668.
Yuan HM, Guo WD, Zhao LJ, Yu Y, Wu JZ, Zhang LG, Cheng LL, Zhao DS, Wu GW, Guan FZ. Bioinformatics and expression analysis of the cellulose synthase supergene family in flax., 2016, 49(23): 4656-4668. (in Chinese)
[22] KHOTYLEVA L, GALINOUSKY D, ANISIMOVA N, RAISKI A, LEONTIEV V, TITOK V. Cellulose synthase genes that control the fiber formation of flax (L.)., 2014, 50(1): 26-34.
[23] MOKSHINA N, GORSHKOVA T, DEYHOLOS M K. Chitinase-like (CTL) and cellulose synthase (CESA) gene expression in gelatinous-type cellulosic walls of flax (L.) bast fibers., 2014, 9(2): e97949.
[24] HOBSON N, DEYHOLOS M K. Genomic and expression analysis of the flax () family of glycosyl hydrolase 35 genes., 2013, 14(1): 207-210.
[25] 李翔, 陳信波, 鄒杰, 龍松華, 鄧欣. 亞麻韌皮纖維細胞發(fā)育分子生物學(xué)研究進展. 湖南農(nóng)業(yè)科學(xué), 2011(9): 4-7.
Li X, Chen X B, Zou J, Long S H, Deng X. Advance in molecular biology of cell development in flax bast., 2011(9): 4-7. (in Chinese)
[26] 關(guān)鳳芝, 張福修, 趙德寶, 倪錄. 亞麻纖維發(fā)育規(guī)律的研究. 中國麻業(yè)科學(xué), 1993(1): 14-17.
Guan F Z, Zhang F X, Zhao D B, Ni L. Study on development of flax fiber., 1993(1): 14-17. (in Chinese)
[27] 李明, 王克榮. 關(guān)于亞麻韌皮纖維生長發(fā)育階段劃分的探討. 中國麻業(yè)科學(xué), 1996(1): 29.
LIM, Wang K R. Discussion on the growth stage of flax bast., 1996(1): 29. (in Chinese)
[28] GORSHKOVA T, MORVAN C. Secondary cell-wall assembly in flax phloem fibres: Role of galactans., 2006, 223(2): 149-158.
[29] SMITH D L, GROSS K C. A family of at least seven beta-galactosidase genes is expressed during tomato fruit development., 2000, 123(3): 1173-1183.
[30] TATEISHI A, INOUE H, YAMAKI S. Fluctuations in activities of three beta-galactosidase isoforms from ripening avocado () fruit and their different activities against its cell wall polysaccharides as substrates., 2001, 70(5): 586-592.
[31] 田愛梅, 劉金龍, 曹家樹. 植物β-半乳糖苷酶. 中國細胞生物學(xué)學(xué)報, 2014(5): 703-707.
Tian A M, Liu J L, Cao J S. Beta galactosidase in plants., 2014(5): 703-707. (in Chinese)
[32] PYDIURA N A, BAYER G Y, GALINOUSKY D V, YEMETS A I, PIRKO Y V, PADVITSKI T A, ANISIMOVA N V, KHOTYLEVA L V, KILCHEVSKY A V, BLUME Y B. Bioinformatic search for cellulose synthase genes in flax () and their phylogenetic analysis., 2015, 49(5): 279-287.
[33] 宣云, 趙竑緋, 郭肖穎, 任杰, 王艷, 盧碧蕓. 植物細胞壁重構(gòu)酶木葡聚糖內(nèi)轉(zhuǎn)糖苷酶/水解酶(XTH)的研究進展. 中國農(nóng)學(xué)通報, 2016(18): 83-88.
Xuan Y, Zhao H F, Guo X Y, Ren J, Wang Y, Lu B Y. Plant cell wall remodeling enzyme xyloglucan endotransglucosylase/hydrolase (XTH)., 2016(18): 83-88. (in Chinese)
(責(zé)任編輯 李莉)
附錄1 引物序列
Tabel 1 Primersequences

基因名稱 Genes 基因代碼 The genetic code引物序列Primersequences LuCESA1Lus10018902FGACTAATGGCCAGCAGATTTCGG Lus10018902RTCGAGATAAGGAGACGACTGCAC LuCESA3Lus10007538FATCTCCTGGACGGTACTCGATG Lus10007538RTCTCCCAGTCTAATGGTTGCTCTG LuCESA4Lus10008225FTGCAGAATGGACAGAACGAGTGG Lus10008225RGTTGTCTGGCATCAGCGGTTAAG LuCESA5Lus10003525FGGCTATGAATGCACTGCGATGG Lus10003525RTTAATGAGGAGGTCGTGGCTCTGG LuCESA6Lus10002940FCTGTTCTGCGGGTCTAGGAAGAAC Lus10002940RTCCAGCCAACCTGTTCAACTTC LuCESA7Lus10007296FGGTTTGCTTTCTCTTGGGTGCTG Lus10007296RACCTGGCCTTTCAAACCTTGC LuCESA8Lus10029245FGGTTTGCTTTCTCTTGGGTGTTGG Lus10029245RACCTGGCCTTTCAAACCTTGC LuCESA9Lus10002939FCAGAGAGGCCATCCAAGTTATCAG Lus10002939RCGAACCATATATCCAGCCAACCTC LuCESA10Lus10028597FTGAAGCAGGAGAAGGGAATGATGC Lus10028597RGAGCATCGTCAGCCATTTGAAGC LuSuSyLus10030176FTAGATTTCGGGATGGCCGTTTG Lus10030176RACGGATGACAACATTGCATTGCC LuXTH4Lus10011052FCCACCTCGATAACTACACTGGAAC Lus10011052RTCCACCAACCATCTTGATGTGC LuBGAL1 Lus10008974FACTCATCGGTAGCGCTTATGGC Lus10008974RGAAGTGTAAGCCAACATTCGCAAG LuBGAL3 Lus10006009FTGCATTCCTGGCGAACTACGAC Lus10006009RGCACTCTCGGAGCAGAAACCTTTG LuBGAL5 Lus10000701FAGCTGTCATGCCTTGCACTCATAC Lus10000701RACCAATTCTGTCCAACACAAAGCC LuBGAL6 Lus10015625FGGTGAACATCTGAGCCTTCATTCC Lus10015625RGCAGGAGCATTGAATGTGGTCTTG LuBGAL9 Lus10024292FTAGCGGTTGGATTGCCGAATG Lus10024292RACCATCTGTGGCCTGTCAAGTC LuBGAL20 Lus10003343FAGCTGTTTGCTTCTCAGGGAGAC Lus10003343RTTGAGCAACGGCCATTTGAGC LuBGAL22 Lus10025108FACCGAGAATTGGTCTGGATGGTG Lus10025108RAGTTGACCGGTGAGGATTAGTGG LuETIF1 Lus10015124 FGATCCAGCTTCAAGGTGACCAG Lus10015124 RTCACAATTCCAGCCTGAACGAG
The Expression of Genes Related to Cell Wall Formation at Fast growth Stage in Flax(L.)
JIANG HaiXia, GUO DongLiang, LI YuHuan, YAN WenLiang, YANG LiangJie, XIE LiQiong
(College of Life Science and Technology, Xinjiang University, Urumqi 830046)
【Objective】The development of flax bast cells is separated into two non-overlapping phases, cell elongation and secondary cell wall thickening phases which are splitted by the snap point at rapid-growing phase. Genes of β-glycosyl hydrolyse (LuBGALs) family and Cellulose synthase (LuCESAs) family focused on the role of cell wall formation in flax () fibres were analyzed in order to gain a deeper insight into the development of cell walls in rapid growth of flax.【Method】The microstructure of the cell walls at snap point (SP) of stem was studied by transmission electron microscope.andwere analyzed by quantitative real-time PCR (qRT-PCR) in the bast fiber of stem, leaves and root of flax at 45 days after germination. 【Result】The width of cell walls at top of SP (TOP) was 110 nm. Though they at MID were about 500 nm and BOT about 650 nm under the SP were thicker than theirs in TOP, it was difficult to find different layers in the thicken cell wall. It showed that the cell walls above the SP were undergone cell elongation and below were thickening but not secondary cell walls thickening. The expression ofat TOP was significantly lower than it’s in MID and BOT, indicating that it was mainly involved in cell wall thickening of fiber cell in the stem. While the expressions of,,andat TOP were the highest and the followed was MID zone, which showed that those genes mainly worked in cell wall reconstructure and bast fiber elongation.was higher at TOP and lower at MID than other genes, which demonstrated thatplayed a role in cell wall formation. And the expression of othergenes were low.,,,,, andhighly expressed in TOP zone and were significantly higher than their expression in MID and BOT. The expression ofandMID was lower than theirs in BOT and there was no significant difference between MID and BOT of othergenes. The results suggested that thesegenes played an important role in cell elongation of flax bast fiber combining their expression in different organs. The character ofexpression indicated that a lot of UDPG was required in cell elongation and thickening of bast fiber.played a role in cell wall development of flax.【Conclusion】The cell walls of bast fiber had not processed secondary thickening in the stem at fast growth stage.,,,,,,, andmainly promoted cell elongation of cell wall in flax.played an important role in cell wall thickening.andalso took part in the development of cell wall.
flax(L.); bast fiber; cell wall formation; gene expression
2016-11-30;接受日期:2017-03-17
國家自然科學(xué)基金(31160056/C020408)
江海霞,E-mail:738230018@qq.com。通信作者謝麗瓊,E-mail:picea@sina.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
