基于UPLC—Q—TOF/MS的RAW264.7細胞炎癥模型的代謝指紋分析
2017-07-13 21:16:52高山山郭慧清張澤坤白光燦高曉燕
中國中藥雜志 2017年12期
高山山+郭慧清+張澤坤+白光燦+高曉燕+馬長華

[摘要] 為揭示炎癥細胞模型的極性代謝組特征,該文選擇RAW264.7細胞為研究載體,以LPS刺激誘導炎癥細胞模型,進行細胞代謝組學的指紋分析。具體方法為:篩選、優化了細胞內極性代謝物的提取方法,建立了利用超高效液相色譜-四極桿飛行時間質譜(UPLC-Q-TOF/MS)進行細胞內代謝組的數據采集方法,采用正交偏最小二乘判別分析(OPLS-DA)進行數據處理,篩選并初步鑒定了17種差異代謝物。結果顯示,采用MeOH-CHCl3-H2O(8∶1∶1)作為提取溶劑,色譜峰數目較多,細胞內極性代謝物提取效率較高,分析方法的穩定性較好;與正常細胞相比,炎癥細胞的蛋白質代謝、糖代謝、核苷酸代謝等代謝通路受到擾動。該研究建立了基于UPLC-Q-TOF/MS的RAW264.7細胞代謝組學指紋分析方法,為揭示細胞炎癥機制及抗炎藥物機制研究奠定基礎。
[關鍵詞] 超高效液相色譜-四極桿飛行時間質譜; 代謝指紋分析; 代謝組學; 炎癥細胞; 正交偏最小二乘判別分析
[Abstract] In order to reveal the properties of polar metabolome in inflammatory cells, we selected LPS-induced RAW264.7 inflammatory cell models as the carrier for the research of metabolic fingerprint analysis. In this study, an ultra performance liquid chromatography coupled with quadrupole-time-of-flight mass spectrometry (UPLC-Q-TOF/MS)-based metabolomics protocol was optimized for the extraction of polar metabolites from RAW264.7 cell line. Then orthogonal partial least squares discriminant analysis (OPLS-DA) was used to process the metabolic data, and finally, a total of 17 metabolites were selected and identified. The results showed that MeOH-CHCl3-H2O (8∶1∶1) was chosen as the optimal extraction solvent to achieve higher number of chromatographic peaks, with the best relative extraction efficiency and stability. Comparing with the normal cells, the inflammatory cells presented an abnormal metabolism in protein, carbohydrate, nucleotide and phospholipids. In this study, a UPLC-Q-TOF/MS-based metabolomics protocol for the polar metabolites from RAW264.7 cell line was developed, which may provide important information for the study of mechanism of inflammation and the anti-inflammatory drugs.
[Key words] ultra performance liquid chromatography coupled with quadrupole-time-of-flight mass spectrometry(UPLC-Q-TOF/MS); metabolic fingerprint analysis; metabolomics; inflammatory cells; orthogonal partial least squares discriminant analysis (OPLS-DA)
細胞代謝組學是系統生物學的重要組成部分[1],分為代謝指紋分析和代謝足跡分析,其研究對象分別為胞內代謝組與胞外代謝組,兩者具有高度的信息互補性[2]。胞內代謝組是細胞內復雜生化活動狀態的綜合表現,細胞內代謝物濃度的改變能夠反映細胞基因組的改變和細胞對于外部刺激的響應,因而,獲取細胞代謝物信息對于系統地了解細胞的性質和功能、篩選藥物以及尋找用于分型的生物標志物至關重要。
RAW264.7小鼠單核巨噬細胞,具有很強的黏附和吞噬抗原的能力,免疫學研究中常采用細菌脂多糖(lipopolysaccharides,LPS)刺激RAW264.7細胞造炎癥反應模型。巨噬細胞的狀態及其功能發揮與細胞代謝密切相關,其自身代謝水平的差異甚至能夠影響其極化狀態,從而決定了不同的功能[3]。因此,利用RAW264.7細胞進行炎癥機制的研究具有重要意義。目前,文獻報道的RAW264.7細胞炎癥模型的研究多是以細胞內弱極性小分子代謝物組為主的脂質組學,揭示了以花生四烯酸為代表的脂質代謝物在炎癥細胞中被擾動的情況[4-5]。
然而,炎癥細胞中的極性代謝組亦發生改變。Newsholme P等[6]采用Gilford自動記錄式分光光度計測定活化的巨噬細胞內的己糖激酶和葡萄糖-6-磷酸脫氫酶活性,發現2種酶活性均增強,表明糖酵解途徑和磷酸戊糖途徑代謝加強。Western blot實驗結果證明活化的巨噬細胞內誘導型一氧化氮合酶(iNOS)表達增強,表明精氨酸的iNOS代謝通路增強。RAW264.7細胞炎癥模型的胞內極性代謝物組發生了改變,但是針對其細胞內極性代謝物組的研究尚未見報道。
因此,本文針對RAW264.7細胞內極性小分子代謝物組,對細胞代謝物提取溶劑及檢測條件進行優化,建立基于UPLC-Q-TOF/MS的RAW264.7細胞極性代謝物檢測方法,進行細胞代謝指紋分析,明確RAW264.7細胞炎癥模型的差異代謝物,發現炎癥擾動的細胞代謝通路,推測RAW264.7細胞極化過程,為揭示炎癥細胞的生物化學變化基礎提供數據支撐,同時為細胞極性代謝物組的代謝指紋分析提供了方法學參考。
1 材料
1.1 儀器與試劑
CO2細胞培養箱(Thermo Scientific 公司);ACQUITY I-Class超高效液相色譜儀;Waters Xevo-G2-SQ-TOF-MS質譜系統(美國Waters公司);D-3750 Osterode 高速冷凍離心機(德國Thermo Fisher公司);Millipore Synergy UV型超純水機(美國Millipore公司);水浴氮吹儀(北京成盟偉業科技有限公司);xw-80A旋渦混合器(上海醫科大學儀器廠);多功能酶標儀(Thermo Scientific 公司);100 mm細胞培養皿(美國Corning公司)。
胎牛血清(美國Corning公司)、DMEM細胞培養基(Hyclone 公司);胰蛋白酶、PBS(中國Solarbio公司);一氧化氮測試試劑盒(普利萊公司);乙腈、乙酸銨(LC-MS級,美國Fisher Scientific公司);甲醇(HPLC級,美國Fisher Scientific公司);LPS(Sigma 公司);其余試劑至少為分析純。
1.2 細胞來源
小鼠巨噬細胞株RAW264.7購自中國科學院協和細胞資源中心。
2 方法
2.1 細胞培養
RAW264.7小鼠單核巨噬細胞用含10%胎牛血清、青霉素(1×105 U·L-1)及鏈霉素(100 mg·L-1)的DMEM培養基培養于37 ℃,5% CO2的培養箱中培養。六孔板中每孔接種1×106個細胞,孵育24 h后炎癥模型組加入LPS,使其終質量濃度為100 μg·L-1,正常組加入相同體積培養基,每組平行培養6份。培養24 h后進行樣品制備。
2.2 Griess試劑盒檢測培養基中NO含量
按照試劑盒說明書制備NO標準曲線。吸取50 μL培養基上清液于96孔細胞培養板中,加入50 μL Griess R1試劑,室溫避光靜置5 min,各孔再加入50 μL Griess R2試劑,室溫避光靜置5 min,酶標儀540 nm下檢測吸光度,并用校準曲線確定實驗各組NO的濃度。
2.3 細胞樣品制備
去除培養基,用4 mL 37 ℃超純水清洗2遍,液氮淬滅。加入1 mL含有5 μmol·L-1α-氨基庚二酸內標的MeOH-CHCl3-H2O(8∶1∶1)提取溶劑,用細胞刮刀將細胞刮下,該操作置于冰上,提取溶劑轉移至1.5 mL離心管中,渦旋30 s,在液氮-室溫下反復凍融循環2次使細胞破碎,4 ℃以13 500 r·min-1離心15 min后取上清液,在檢測前儲存在-80 ℃冰箱中。檢測前將上清液在室溫下用氮氣吹干,殘渣用50%乙腈復溶,進行LC-MS分析。
2.4 色譜條件
采用ACQUITY UPLC BEH Amide色譜柱(2.1 mm×100 mm,1.7 μm,美國Waters公司);流動相A:含有5 μmol·L-1的乙酸銨水溶液(pH 9.9);流動相B:乙腈;流速:0.4 mL·min-1;梯度洗脫程序:0~1.0 min,0~99%B;1.0~5.0 min,99%~75%B;5.0~6.0 min,75%B;6.0~8.0 min,75%~70%B;8.0~9.5 min,70%~65%B;9.5~10 min,65%~99%B。
2.5 質譜條件
質譜系統采用電噴霧離子源,負離子模式掃描;掃描范圍m/z 50~1 000;離子源參數毛細管電壓為2 000 V,錐孔電壓為40 V,離子源溫度為100 ℃,鞘流氣為氮氣,900 L·h-1,鞘流氣溫度為450 ℃,掃描時間為0.2 s,碰撞氣體為氬氣。低能量掃描時碰撞能量為6 eV,高能量掃描時碰撞能量為10~40 eV。
2.6 方法學考察
在確定MeOH-CHCl3-H2O(8∶1∶1)為最佳細胞代謝物提取溶劑后進行了方法學考察。方法學考察部分包括精密度、重復性、樣品穩定性及系統穩定性4個考察內容。吸取等量的細胞代謝物提取液,混合均勻,作為質控樣本(QC)。精密度考察方法是1個QC樣本平行進樣6次,分別計算峰面積及保留時間的相對標準偏差。重復性考察方法是平行制備6份QC樣本,每份進樣1針,分別計算峰面積及保留時間的相對標準偏差,樣品穩定性考察是平行制備6份QC樣本,4 ℃放置12 h,進樣分析,分別計算峰面積及保留時間的相對標準偏差。系統穩定性考察是每8個樣品隨行一針QC樣本,分別計算峰面積及保留時間的相對標準偏差。
2.7 數據處理與分析
NO含量檢測結果采用SAS 8.2統計軟件進行統計學處理。所有數據用±s表示,組間比較用獨立樣本t檢驗,P<0.05為差異有統計學意義。
將LC-MS采集到的數據導入ProgenesisQI軟件(Waters 公司),對色譜峰進行峰提取、峰匹配、峰對齊、峰識別以及歸一化處理,得到的數據再導入EZinfo2.0軟件進行多變量分析,選取變量權重值VIP>1,并結合S-plot圖篩選正常組和炎癥模型組細胞的代謝物譜差異,采用t檢驗對上述差異代謝產物進行統計學分析,P<0.05有統計學差異。差異性代謝產物的結構依據碎片離子(MSE)信息及HMDB,METLIN,LIPID,KEGG等數據庫信息進行鑒定。
3 結果與討論
3.1 Griess試劑盒檢測培養基中NO含量結果
用一系列對照品濃度與其對應的吸光度A作標準曲線,并進行直線相關回歸分析,得出其直線回歸方程為y=115.41x-6.962 2,R2=0.999 9。將每組樣品測得的A代入該直線回歸方程中,即得到相應的NO含量。加入LPS后培養24 h,正常組RAW264.7 細胞NO分泌量為0.80 μmol·L-1,模型組RAW264.7 細胞NO分泌量為7.01 μmol·L-1,LPS明顯提高了RAW264.7細胞NO的分泌(P<0.01),說明造模成功。
3.2 細胞提取方法的優化
本研究采用超純水沖洗細胞,液氮淬滅,同時考察了4種提取溶劑對細胞代謝物檢測的影響,這4種提取試劑包括MeOH,ACN,MeOH-ACN-H2O(2∶2∶1),MeOH-CHCl3-H2O(8∶1∶1)。按照2.3項樣品制備方法,分別采用4種提取試劑制備樣品并進行檢測,見圖1A,發現MeOH-CHCl3-H2O作為提取溶劑時色譜峰個數較多。本文選擇了部分常見極性代謝物來評價4種提取溶劑的提取效率,每種代謝物的相對提取率采用內標物進行標準化,并將ACN組代謝物的相對提取效率設為100%作為參照,發現采用MeOH-CHCl3-H2O作為提取溶劑時極性代謝物的相對提取率高,見表1。主成分分析是以較少個數的主成分來反映原始數據的主要信息,因此,PCA圖中點的聚集情況可以表明樣品間變異性。由4種提取溶劑制備樣品的PCA圖,見圖1B。發現,采用MeOH-CHCl3-H2O作為提取劑同組樣品間穩定性最高。因此選取MeOH-CHCl3-H2O作為RAW264.7細胞極性代謝產物的提取溶劑。
3.3 分析條件的優化
3.3.1 色譜條件優化 細胞的代謝途徑復雜多樣,研究表明,LPS刺激巨噬細胞后,細胞的糖代謝、氨基酸代謝、脂代謝都發生了改變,為全面表征其代謝產物變化,本研究考察色譜柱類型、流動相組成和離子檢測模式。實驗中分別采用T3色譜柱和BEH Amide色譜柱進行比較,發現BEH Amide因其酰胺官能團同時具有氫原子受體和供體位點,極性代謝物保留時間延長。調整流動相組成,發現使用含5 mmol·L-1乙酸銨的水(pH 9.9)-乙腈流動相體系,采用2.4項所述的流動相梯度進行洗脫,分離效果最佳。
3.3.2 質譜條件優化 該文對毛細管電壓、錐孔電壓、離子源溫度、鞘氣流速及其溫度、掃描時間、低能量通道碰撞能量以及高能量碰撞能量等參數進行了優化,最終以檢出色譜峰的數量和強度為指標確定了質譜檢測條件。針對RAW264.7細胞內極性小分子代謝物組,包括糖酵解產物、三羧酸循環產物、氨基酸代謝產物等極性代謝物,通過考察同一代謝產物在正、負離子2種檢測模式下的出峰情況及準分子離子峰的響應強度,發現三羧酸循環產物和磷酸糖類化合物只在ESI-下有信號,氨基酸類化合物和腺苷類化合物在2種檢測模式下均有信號,因此本研究選擇負離子模式進行數據采集。
3.4 分析方法考察
在總離子流圖上選取內標α-氨基庚二酸其保留時間為6.42 min,m/z為174.079 4,以及tR,m/z分別為4.24 min,218.100 3;5.45 min,124.008 7;6.87 min,565.049 3;7.17 min,306.073 6這5個色譜峰進行代謝組學方法學考察,計算保留時間與峰面積的RSD。保留時間的精密度、重復性、系統穩定性的RSD分別為0.050%~0.19%,0.030%~0.40%,0.060%~1.0%,4 ℃放置12 h穩定性的RSD為0.060%~1.3%。峰面積的精密度、重復性、系統穩定性的RSD分別為0.23%~1.5%,0.16%~0.87%,0.34%~1.9%,4 ℃放置12 h的穩定性的RSD為0.32%~1.2%。方法學驗證結果表明操作和儀器均能進行代謝組學分析。
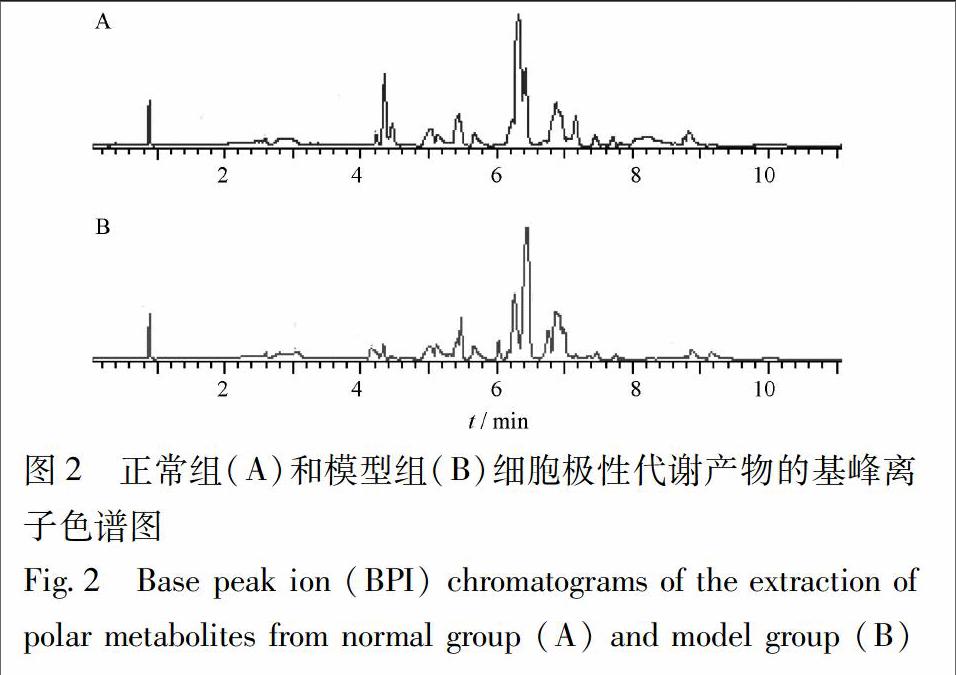
在最優UPLC-MS條件下,分別對正常組、模型組細胞樣品進行檢測,得到細胞代謝指紋圖譜,見圖2,差異部分主要出現在5~7 min處,這些部位可能存在潛在炎癥生物標志物。
3.5 與炎癥相關差異代謝物的尋找
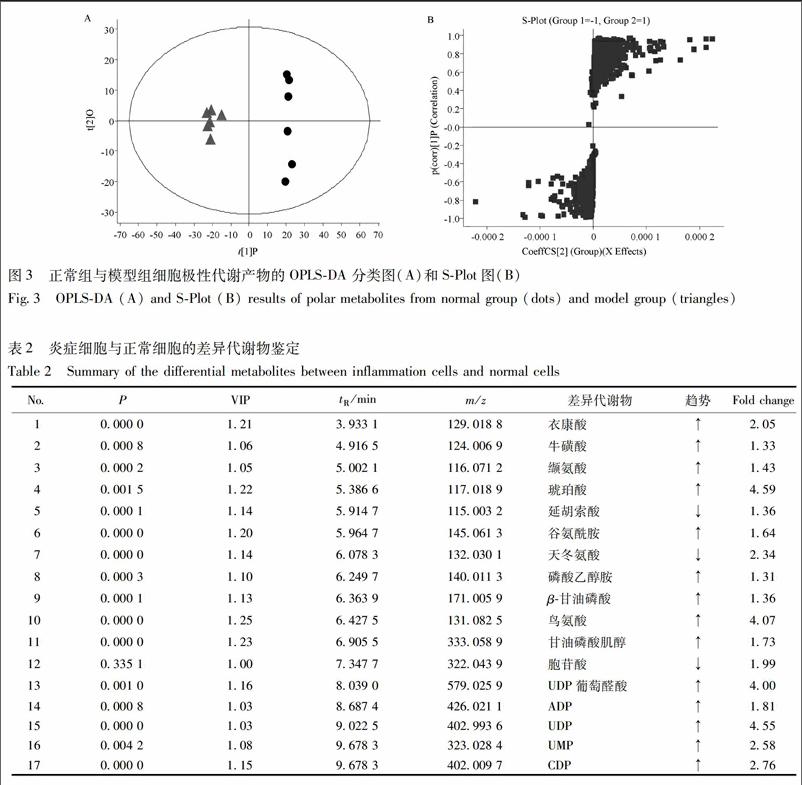
將UPLC-Q-TOF/MS數據導入ProgenesisQI軟件,按照2.7項操作處理數據后,采用OPLS-DA模型進行多維統計分析,篩選VIP>1的峰作為潛在差異代謝物。正常組與模型組的OPLS-DA得分圖和S-Plot圖,見圖3。通過HMDB和METLIN數據庫搜索差異物母離子和子離子的精確質荷比,初步鑒定了17個潛在差異代謝物,見表2。
將鑒定出的差異代謝物輸入MPetA數據庫中,構建代謝通路圖,見圖4。代謝通路分析表明,丙氨酸-天冬氨酸-谷氨酸代謝、三羧酸循環、精氨酸和脯氨酸代謝等代謝通路與LPS誘導的炎癥反應相關。
LPS刺激會導致細胞內發生“Warburg效應”,即糖酵解和磷酸戊糖途徑作用加強,而線粒體內的有氧分解減弱,表現為葡萄糖的大量攝取、高糖酵解速率和核酸合成速率等變化。UDP-葡萄糖醛酸由葡萄糖通過糖醛酸途徑生成,是葡糖醛酸的活性供體,也是UDP-單糖由六碳糖轉變為五碳糖的關鍵底物。與正常組相比,模型組中UDP-葡萄糖醛酸含量顯著升高,是正常組的4倍,表明糖醛酸途徑代謝增強。UDP-葡萄糖醛酸代謝生成的葡萄糖醛酸參與抗壞血酸代謝、磷酸肌醇代謝等代謝活動中。
LPS刺激巨噬細胞使得細胞線粒體氧化磷酸化水平降低,三羧酸循環產物累積,尤其是琥珀酸。Abhishek K Jha等[7]采用平行的代謝組學與轉錄組學數據網絡集成發現被LPS激活的RAW264.7細胞在異檸檬酸脫氫酶處和琥珀酸處發生了代謝中止,即異檸檬酸不生成α-酮戊二酸而生成衣康酸,琥珀酸不再繼續代謝成延胡索酸[8]。累積的琥珀酸主要來源于谷氨酰胺代謝,γ-氨基丁酸旁路對此也有貢獻[9]。實驗結果表明,與正常組相比,炎癥模型組中的琥珀酸、衣康酸含量升高,延胡索酸含量下降,其中模型組琥珀酸含量是正常組的4.59倍,模型組衣康酸含量是正常組的2.05倍,說明琥珀酸和衣康酸可以作為RAW264.7炎癥細胞的生物標志物。
炎癥反應可改變機體的蛋白質代謝和氨基酸需要,這主要是因為細胞因子(如IL-1,IL-6,TNF-α)分泌的增加。在免疫反應中,氨基酸將發生重分配并主要用于合成參與炎癥和免疫反應的蛋白質以及參與免疫細胞增殖和其他免疫反應的重要化合物[10-11]。實驗結果中谷氨酰胺、纈氨酸、鳥氨酸含量升高,天冬氨酸含量降低,涉及了2個代謝通路,即丙氨酸、天冬氨酸、谷氨酸代謝通路,和纈氨酸、亮氨酸和異亮氨酸生物合成通路。谷氨酰胺與谷氨酸可以相互轉化,谷氨酸代謝生成γ-氨基丁酸,γ-氨基丁酸進一步代謝生成琥珀酸。谷氨酰胺在谷氨酰胺轉酰胺酶作用下是將氨基氮轉移至不同底物上,為其他代謝反應提供氮源[12]。纈氨酸、亮氨酸和異亮氨酸屬于支鏈氨基酸,本實驗篩選出的差異代謝物為纈氨酸,與正常組比較,炎癥模型組中纈氨酸含量明顯升高,其原因為細胞因子的合成需要纈氨酸等支鏈氨基酸的參與。
實驗結果中模型組磷酸乙醇胺含量顯著升高,磷酸乙醇胺是神經酰胺的代謝產物之一,表明模型組中神經酰胺代謝高于正常組。神經酰胺是鞘磷脂信號途徑的中心分子,參與激活多種蛋白激酶和蛋白磷酸酶,在調節細胞免疫方面起重要作用。炎癥細胞中上調的花生四烯酸含量,分泌的白介素、腫瘤壞死因子-α等都可以激活中性鞘磷脂酶,水解鞘磷脂,導致細胞中神經酰胺含量升高[13]。
4 結論
本文提出了1種能夠高效、穩定提取RAW264.7細胞中極性代謝產物的方法,并采用UPLC-Q-TOF/MS技術對正常細胞與炎癥細胞進行分析,獲得了代謝物圖譜,結合多元統計分析手段研究正常細胞與炎癥細胞的圖譜變化情況,結果表明,炎癥細胞與正常細胞的代謝物譜圖有顯著變化。通過對一些具有顯著性差異的物質進行初步鑒定,得到了17種代謝差異物。與正常細胞相比,炎癥細胞在蛋白質代謝、磷脂代謝、糖代謝方面發生不同程度的紊亂,與文獻報道的采用CE-TOF/MS、氨基酸分析儀檢測結果基本相符[14],證明該方法可以表征RAW264.7細胞極化,為抗炎藥物研究提供了新的研究方法和思路。
[參考文獻]
[1] Fei F, Bowdish D M E, Mccarry B E. Comprehensive and simultaneous coverage of lipid and polar metabolites for endogenous cellular metabolomics using HILIC-TOF-MS[J]. Anal Bioanal Chem,2014,406(15):3723.
[2] 李瑩,傅超美,任波,等. 基于MI-RI大鼠心肌細胞代謝組學研究四逆湯中附子配伍甘草解毒增效機制[J]. 中國中藥雜志,2014,39(16):3166.
[3] Osborn O, Olefsky J M. The cellular and signaling networks linking the immune system and metabolism in disease[J]. Nat Med,2012,18(3):363.
[4] 趙麗麗. NSAIDs干預RAW264.7細胞炎癥模型甘油磷脂代謝的UPLC-Q-TOF/MS分析[D]. 廣州:廣東藥學院,2012.
[5] Kita Y, Takahashi T, Uozumi N, et al. Pathway-oriented profiling of lipid mediators in macrophages[J]. Biochem Bioph Res Co,2005,330(3):898.
[6] Newsholme P, Curi R, Gordon S, et al. Metabolism of glucose, glutamine, long-chain fatty acids and ketone bodies by murine macrophages[J]. Biochem J,1986,239(1):121.
[7] Jha A K, Huang S C, Sergushichev A, et al. Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization[J]. Immunity,2015,42(3):419.
[8] O Neill L A J. A broken krebs cycle in macrophages[J]. Immunity,2015,42(3):393.
[9] Tannahill G M, Curtis A M, Adamik J, et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α[J]. Nature,2013,496(7444):238.
[10] 井明艷,劉波靜,孫建義,等. 氨基酸代謝與免疫反應[J]. 中國畜牧雜志,2007,43(5):37.
[11] 祖克熱古麗·吾買爾尼亞孜,哈木拉提·吾甫爾, 買吾拉尼江·依孜布拉,等. 異常黑膽質成熟劑對移植性宮頸癌(U27)腫瘤模型小鼠血清的代謝組學研究[J].中國現代應用藥學,2015,32(8):905.
[12] 徐詠全,張蓓,張克旭. 谷氨酰胺磷酸核糖焦磷酸轉酰胺酶研究進展[J]. 生物技術通訊,2003(6):535.
[13] 金道忠,朱興族. 神經酰胺代謝及凋亡信號調節[J]. 生命科學,2006(5):481.
[14] Sugimoto M, Sakagami H, Yokote Y, et al. Non-targeted metabolite profiling in activated macrophage secretion[J]. Metabolomics,2012,8(4):624.
[責任編輯 張燕]
