狨猴B2m基因沉默位點在細胞水平的驗證
2017-06-09 08:58:47鄧怡晨向志光滕永康劉云波
中國比較醫學雜志 2017年5期
關鍵詞:實驗
鄧怡晨,張 晨,向志光,滕永康,劉云波*
(1.中國醫學科學院醫學實驗動物研究所,北京 100021;2.北京大學生命科學學院,生物膜及膜生物工程國家重點實驗室,北京大學麥戈文腦研究所,北京 100871)
研究報告
狨猴B2m基因沉默位點在細胞水平的驗證
鄧怡晨1,張 晨2,向志光1,滕永康1,劉云波1*
(1.中國醫學科學院醫學實驗動物研究所,北京 100021;2.北京大學生命科學學院,生物膜及膜生物工程國家重點實驗室,北京大學麥戈文腦研究所,北京 100871)
目的 在細胞水平篩選狨猴B2m基因的有效沉默靶點,并進行驗證。方法 查詢人源B2m驗證過的有效siRNA靶位點序列,與狨猴B2m基因序列進行同源性比較,選擇匹配靶點合成shRNA序列。將體外合成的2條干擾序列分別與慢病毒載體 FUGW-TDT連接,構建FUGW-TDT-shb2m干擾表達質粒,在聚乙烯亞胺(polyethylenimine,PEI)介導下轉染293T細胞,轉染后48h,用實時熒光定量法檢測轉染細胞中B2m基因mRNA的水平。結果 篩選出2個與狨猴完全同源的B2m沉默靶位點,分別位于B2mmRNA 的290~310 bp,665~685 bp;B2m兩個靶點在轉錄水平的沉默效率分別為(46.54±7.91)%(P< 0.05)和(83.22±4.37)%(P< 0.0001),差異有顯著性。結論 成功構建成FUGW-TDT-shb2m重組質粒;在細胞水平篩選得到2個有效的B2m基因沉默靶點;為后續有關介導狨猴B2m基因沉默的研究奠定了基礎。
基因沉默;狨猴;β2-微球蛋白;短發夾RNA
β2-微球蛋白(beta2-microglobulin,B2M)是一種內源性低分子量血清蛋白質,廣泛存在于血漿、尿液、腦脊液、唾液以及初乳中。它是人細胞表面主要組織相容性復合體Ⅰ類分子(major histocompatibility complex class I molecule,MHCI)的β鏈,由99個氨基酸組成,無多態性且高度保守,是CD8分子的結合區域[1]。
已有研究表明B2m基因缺陷小鼠在胸腺和外周淋巴器官幾乎缺乏所有成熟的CD4-8+T細胞,導致免疫功能缺陷[2]。目前免疫缺陷疾病動物模型的研究主要是使用大鼠和小鼠作為動物平臺,而非人靈長類動物模型在生物醫學研究中的應用較少。與嚙齒類動物相比較,非人靈長類動物與人類親緣關系和生理生化指標更加接近,因此,研究建立非人靈長類動物模型對于相關疾病的研究有重要意義[3]。
狨猴作為一種較小的新大陸猴,與實驗室內其他常用的靈長類實驗動物,如狒狒、獼猴相比較,由于其體型較小(成年體重300~500 g)、易在實驗室內籠養、便于實驗操作、具有較高的繁殖率、對人類細胞因子或激素具有交叉反應性、具有獨特的行為和認知特性,其在生物醫藥科學和神經科學方面引起廣泛關注[4, 5]。本文從細胞水平篩選及驗證狨猴B2m基因的沉默靶位點,為進一步探究B2m基因在狨猴體內的功能奠定基礎。
1 材料和方法
1.1 材料
293T細胞來至ATCC細胞庫;E.coliDH5α感受態細胞購自北京全式金公司;細胞培養基 DMEM 購自Hyclone;胎牛血清(FBS)、胰蛋白酶為Invitrogen-Gibco公司產品;無內毒素質粒小提試劑盒購自康為世紀公司;逆轉錄試劑盒購購自TaKaRa公司;PCR試劑盒購自TaKaRa公司;shRNA合成由 Invitrogen 公司提供;引物合成由北京擎科新業生物技術有限公司提供;Trizol試劑由 Invitrogen 公司提供;Xho I和Xba I 購自New England Biolabs公司。
1.2 實驗方法
1.2.1 shRNA序列設計
從broad institute 網站中(http://www.broadin stitute.org/rnai/public/)查詢人源B2m基因有效siRNA靶位點序列,通過與狨猴B2m基因序列進行同源比對,選擇完全匹配的兩條siRNA序列,設計shRNA序列[6, 7],由Invitrogen 公司合成。
1.2.2 B2m-RNA干擾慢病毒載體的構建和鑒定
慢病毒載體FUGW-TDT含有Ubi啟動子(Ubiquitin promoter)調控的紅色熒光蛋白(tandem dimer tomato,TDT)報告基因,可與U6啟動子調控的RNA干擾片段共表達[8]。FUGW-TDT 載體經Xho I和Xba I雙酶切線性化后回收,將兩條shRNA分別連入載體FUGW-TDT的U6啟動子下游,即構建重組質粒[9]。連接產物轉化至E.coliDH5α 感受態細胞中,涂布于50 μg/mL氨芐青霉素的LB平板上,37℃培養12 h,挑取單菌落,接種至含有50 μg/mL氨芐青霉素的LB液體培養基中,37℃培養不超過16 h,進行菌液鑒定(菌液PCR及測序引物均為F: 5’ -AGGAAGATGGCTGTGAGG- 3’;R: 5’-GCCTTGTATCGTATAAGC- 3’)。選擇陽性克隆菌液進行質粒抽提,經過質粒測序(北大儀器中心),將成功插入目的片段的質粒命名為FUGW-TDT-shb2m。本實驗的陰性對照為未導入shRNA的FUGW-TDT空質粒。
1.2.3 細胞培養及轉染
用含有10%胎牛血清的DMEM培養液,在6孔板中培養293T細胞,每孔加入5×105個細胞,置5% CO2培養箱中37℃常規培養。24 h內細胞密度達70%~80%時進行轉染。在轉染試劑聚乙烯亞胺(polyethylenimine,PEI)的介導下,按PEI∶質粒=3∶1的比例配制轉染混合物,將重組質粒和空載質粒轉進細胞[10]。實驗設為重組質粒組和空載質粒組,其中,重組質粒組包括FUGW-TDT-shb2m-1和FUGW-TDT-shb2m-2兩個,空載質粒組為陰性對照,每個質粒做3個復孔。48 h后觀察熒光并進行流式分選,收集帶有紅色熒光的細胞。
1.2.4 RT-PCR法檢測B2m基因mRNA表達水平
采用實時熒光定量PCR法分析各組轉染細胞中B2m基因mRNA的表達情況[11]。按照 Invitrogen Trizol Reagent說明書分別提取每組細胞總RNA,然后進行逆轉錄反應,逆轉錄條件為: 37℃ 15 min,85℃ 5 s,得到的cDNA用于進行實時熒光定量PCR反應。B2m基因引物為:上游5’-GGCTATCCAGC GTACTCCAAA-3’下游5’- CACGGCAGGCATACTC ATCTT -3’,擴增長度為248 bp。內參GAPDH的引物為:上游5’- TGACTTCAACAGCGACACCCA -3’,下游5’- CACCCTGTTGCTGTAGCCAAA -3’,擴增長度為121 bp。兩條引物均由primer5軟件設計得來。PCR反應程序:94℃預變性5min,94℃變性30 s,60℃退火30s,72℃延伸30 s,40個循環,72℃延伸10 min。
1.3 統計學方法
采用GraphPad Prism 5.0 軟件進行統計學分析,實驗數據以平均值±標準差(±s)表示,組間比較采用t檢驗分析。以P< 0.05表示差異有顯著性。
2 結果
2.1 狨猴B2m基因shDNA寡核苷酸序列
根據狨猴B2m基因序列,依照shRNA的設計原則和broad institute網站中人源B2m基因有效siRNA靶位點序列,與狨猴B2m基因序列進行同源性比較,選擇完全匹配的兩個靶點(圖1)合成shRNA序列,共兩條核苷酸序列見表1。
2.2 B2M慢病毒沉默載體構建
在靶點序列的正反向組合中間添加一個loop環結構(序列為CTCAAGAGA)以便形成shRNA發夾結構。另外在正義鏈模板的 5’端添加tcga,可與XhoI酶切后的粘性末端互補,在反義鏈模板的 5’端添加ctag,可與XbaI酶切后的粘性末端互補,使其能與空的干擾質粒連接,由此合成靶序列的oligoDNA(圖2A)。之后將oligoDNA連接在U6啟動子的下游,成功構建FUGW-TDT-b2m的shRNA載體(圖2B)。

圖1 狨猴B2m基因序列同源性比對分析Fig.1 Homology analysis of marmoset B2m gene sequence
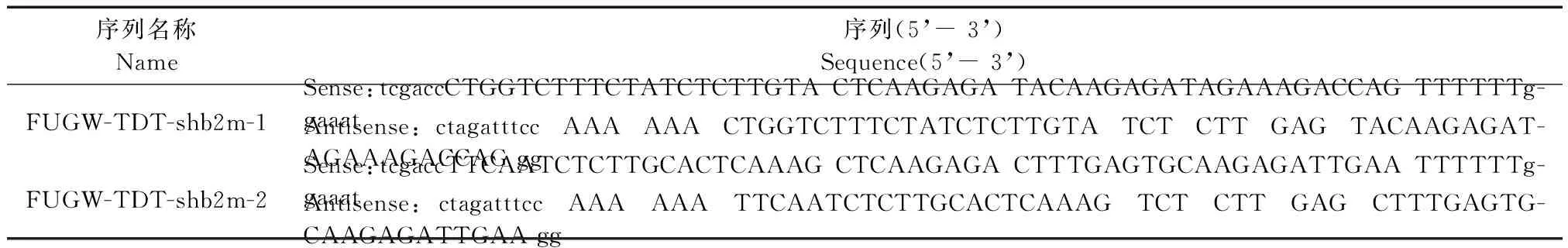
序列名稱Name序列(5’-3’)Sequence(5’-3’)FUGW-TDT-shb2m-1Sense:tcgaccCTGGTCTTTCTATCTCTTGTACTCAAGAGATACAAGAGATAGAAAGACCAGTTTTTTg-gaaatAntisense:ctagatttccAAAAAACTGGTCTTTCTATCTCTTGTATCTCTTGAGTACAAGAGAT-AGAAAGACCAGggFUGW-TDT-shb2m-2Sense:tcgaccTTCAATCTCTTGCACTCAAAGCTCAAGAGACTTTGAGTGCAAGAGATTGAATTTTTTg-gaaatAntisense:ctagatttccAAAAAATTCAATCTCTTGCACTCAAAGTCTCTTGAGCTTTGAGTG-CAAGAGATTGAAgg
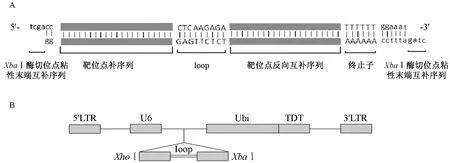
注:(A)shRNA-1的雙鏈寡核苷酸DNA序列結構;(B)FUGW-TDT-shb2m慢病毒載體。圖2 結構示意圖Note.(A)Double-stranded DNA oligonucleotide sequence structure of shRNA-1;(B)FUGW-TDT-shb2m lentiviral vector.Fig.2 The frame of FUGW-TDT-shRNA
2.3 293T細胞中B2M豐度的檢測
選取293T細胞提取總RNA后逆轉錄為cDNA,經普通PCR,電泳檢測細胞中B2m基因的豐度(圖3)。結果顯示293T細胞中B2m基因的表達,說明能夠選用293T細胞于基因抑制效率檢測。
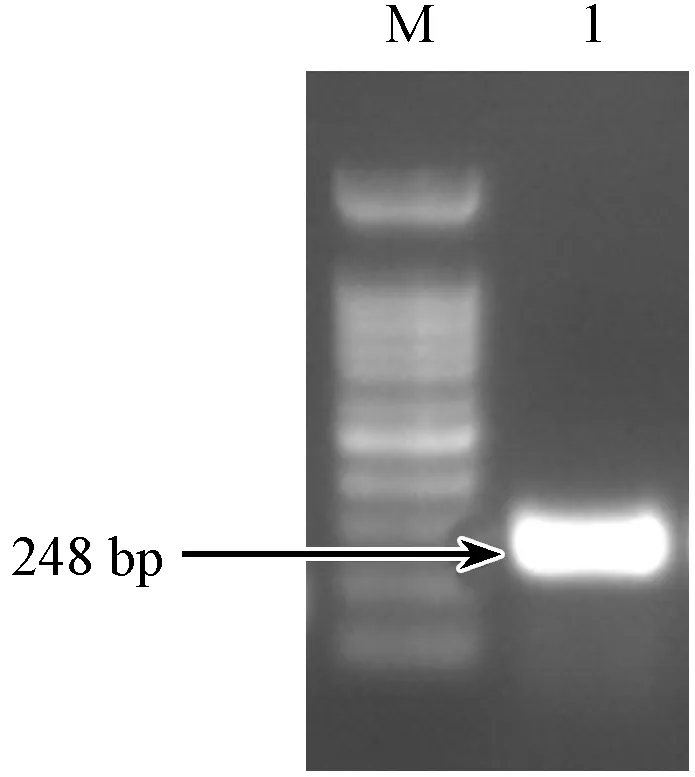
注:(M)Marker;(1)293T。圖3 B2m含量檢測Note.(M)Marker;(1)293T.Fig.3 Detection of B2m abundance
2.4 慢病毒表達載體FUGW-TDT的XhoI和XbaI雙酶切結果
將慢病毒表達載體FUGW-TDT經XhoI和XbaI雙酶切后,產物于1%的瓊脂糖凝膠電泳,結果(如圖4)表明:酶切片段大小10600 bp左右,與預期相符合。然后將其酶切產物采用膠純化回收試劑盒回收目的片段,用于后續的連接反應。
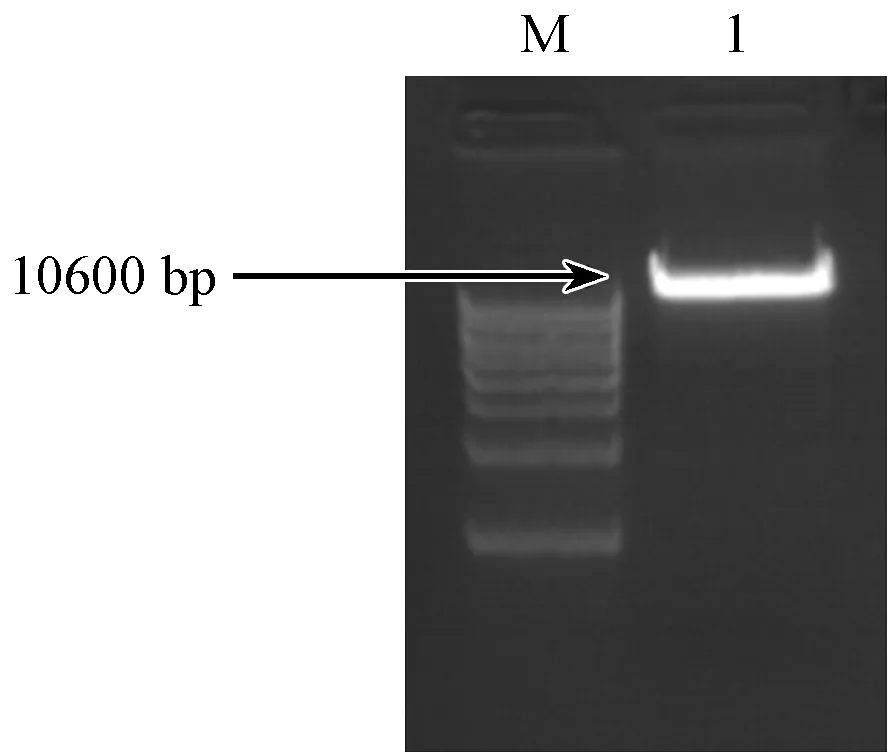
注:(M)DNAMarker;(1)經XhoI和XbaI雙酶切線性化后的質粒載體。圖4 FUGW-TDT載體的雙酶切電泳結果Note.(M)DNA Marker;(1)Linearized plasmid XhoI and XbaI double digestion.Fig.4 Electrophoresis for production of FUGW-TDT vector by double digestion
2.5 菌液PCR鑒定陽性克隆
B2m基因shRNA的寡核苷酸序列,經退火形成雙鏈DNA,與經XhoI和XbaI雙酶切后的FUGW-TDT載體連接,連接產物轉化大腸桿菌DH5α,每個平板挑取3個重組陽性克隆,進行菌液PCR鑒定陽性克隆。重組細菌克隆的PCR產物351 bp(插入片段為61 bp),而經雙酶切后沒有插入片段的FUGW-TDT空載體PCR產物為290 bp。作為對照(圖5),鑒定結果與預期相符。證明B2m基因的shRNA已經定向連入慢病毒表達載體中。
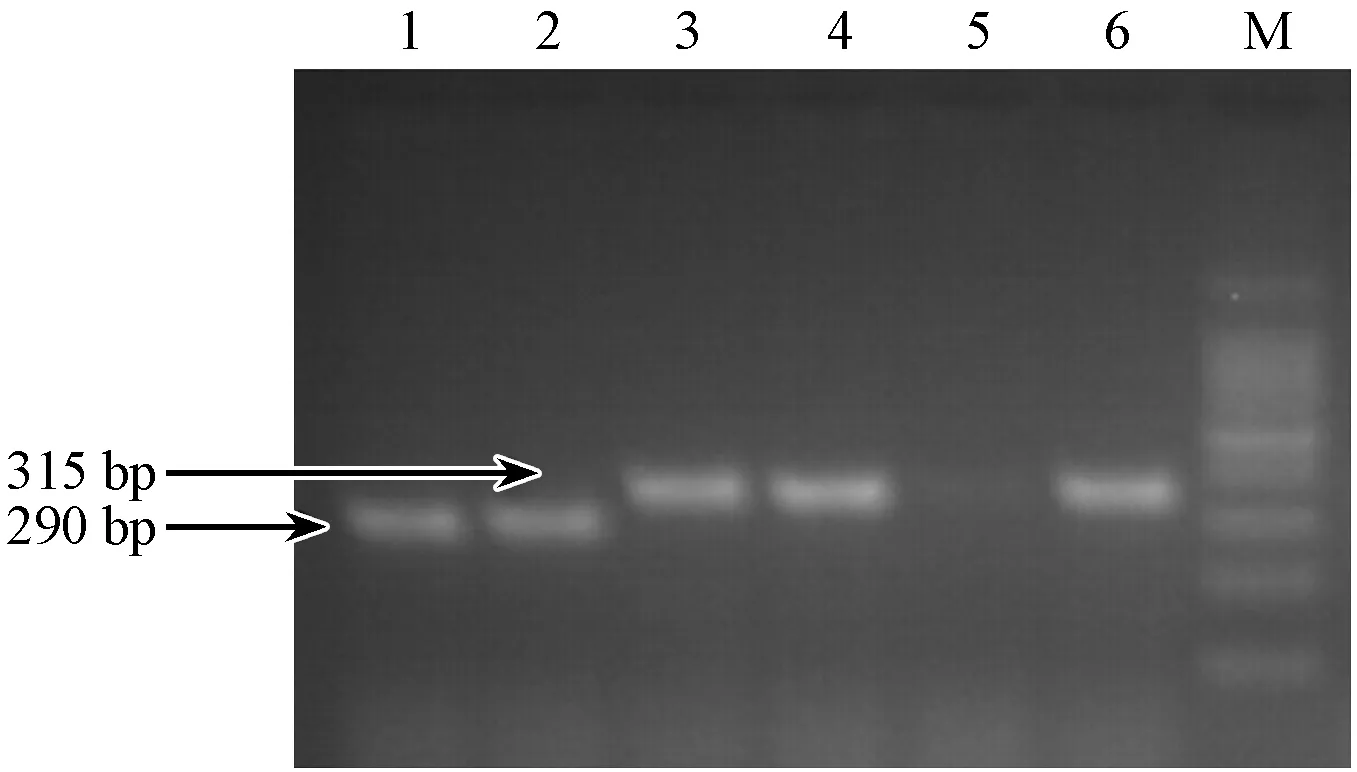
注:(M)DNAMarker;(1~3)FUGW-TDT-shb2m-1;(4~6)FUGW-TDT-shb2m-2。圖5 FUGW-TDT-shb2m載體菌液PCR鑒定結果Note.(M)DNA Marker;(1~3)FUGW-TDT-shb2m-1;(4~6)FUGW-TDT-shb2m-2.Fig.5 Results of PCR FUGW-TDT-shb2m identification
2.6 RT-PCR檢測293T細胞B2m基因的表達
RT-PCR檢測結果顯示,實驗組中2個插入shRNA序列的重組質粒對應的B2m基因mRNA表達量都明顯低于對照組,其中,FUGW-TDT-shb2m-1組與對照組相比,其沉默效率為(46.54±7.91)%(P< 0.05),FUGW-TDT-shb2m-2組與對照組相比,其沉默效率為(83.22±4.37)%(P< 0.0001),實驗批數n=3,差異有顯著性(圖6)。
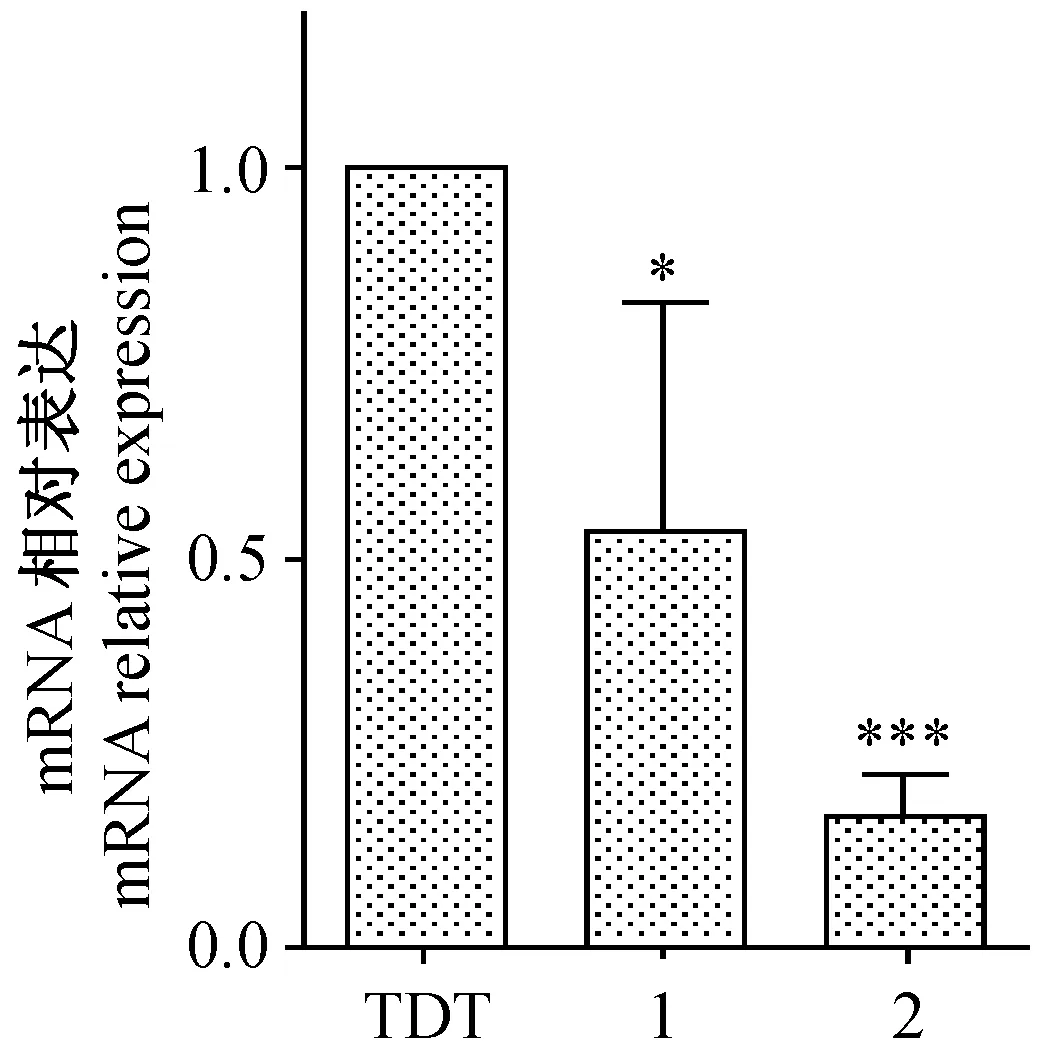
注:(TDT)對照組FUGW-TDT;(1)FUGW-TDT-B2m-1;(2)FUGW-TDT-b2m-2;與對照組FUGW-TDT相比,*P<0.05,***P< 0.0001。圖6 SYBR-Green法 Real-time PCR檢測B2m基因mRNA的表達(n=3)Note.(TDT)Control group FUGW-TDT;(1)FUGW-TDT-b2m-1;(2)FUGW-TDT-B2m-2; Compared with control group FUGW-TDT,*P<0.05,***P< 0.0001.Fig.6 Detection of B2m mRNA by SYBR-Green Real-time PCR
3 討論
很多種類的免疫缺陷動物模型已經建立起來,在免疫學、遺傳學、腫瘤學、自身免疫性疾病、微生物學等諸多方面發揮重要作用[12]。實驗室常用的免疫缺陷動物包括裸小鼠、SCID小鼠等,但相對于嚙齒類動物,非人靈長類動物在遺傳、進化、生理等方面與人類高度相似,被認為是最理想的疾病治療和預防藥物研究的模型,并已廣泛應用于評估抗病藥物和疫苗的安全及藥代工作[13, 14]。
狨猴現在已經成為重要的研制人類疾病模型的動物,在基因修飾動物研究方面有很多優勢條件,比如相對成熟的輔助生殖技術和與人類進化的親緣性。近年來,狨猴在神經科學方面及帕金森、肝炎等疾病動物模型方面具有重要地位[15]。但是, 狨猴疾病模型無法替代舊大陸猴在構建HIV、結核病等人類疾病模型的獨特優勢, 所以應根據課題的需求和具備的條件,采用相應的動物品種[16]。
B2m是通過非共價鍵與MHCI類分子的α鏈胞外段相互作用,不參與MHCI類分子同抗原多肽的結合,但對維持MHCI類分子天然構型的穩定性及其表達起關鍵作用。有文獻報道B2m基因缺陷小鼠表現為免疫缺陷,且B2m基因缺陷小鼠在無病原體環境下培養,可正常發育及繁殖[17],為探究狨猴B2m基因沉默后是否有免疫降低甚至缺陷的現象,首先從細胞水平上篩選沉默效率高的B2m基因靶點。本文中,將篩選的B2m基因沉默靶點插入帶紅色熒光的慢病毒質粒(FUGW-TDT)后轉染HEK293T細胞,通過real-timePCR方法分析,結果表明在RNA干擾作用下,轉染的細胞中B2m基因在轉錄水平比對照組顯著降低,說明RNA干擾靶點有效,可以經過病毒包裝后進行狨猴胚胎和個體水平上驗證實驗,為構建狨猴免疫缺陷模型奠定基礎。
[1] Chiou SJ,Chen CH. Decipher beta2-microglobulin: gain- or loss-of-function (a mini-review)[J]. Med Sci Monit Basic Res, 2013, 19: 271-273.
[2] Koller BH,Marrack P,Kappler JW,etal. Normal development of mice deficient in beta 2M, MHC class I proteins, and CD8+ T cells. 1990[J]. J Immunol, 2010, 184(9): 4592-4595.
[3] Van Rompay KK. Evaluation of antiretrovirals in animal models of HIV infection[J]. Antiviral Res, 2010, 85(1): 159-175.
[4] Sasaki E. Prospects for genetically modified non-human primate models, including the common marmoset[J]. Neurosci Res, 2015, 93: 110-115.
[5] Kishi N,Sato K,Sasaki E,etal. Common marmoset as a new model animal for neuroscience research and genome editing technology[J]. Dev Growth Differ, 2014, 56(1): 53-62.
[6] Grinev VV. [Design and quality control of short interfering RNAs][J]. Mol Biol (Mosk), 2012, 46(6): 827-845.
[7] Zhao H,Zhang JY,Xu WH,etal. [Construction and identification of lentiviral vector for RNA interference targeting STUB1 gene][J]. Zhejiang Da Xue Xue Bao Yi Xue Ban, 2010, 39(6): 623-627.
[8] Cheng L,Shu R,Song ZC,etal. [Reconstruction and expression of recombinant lentiviral vector for human amelogenin in 293T cell line][J]. Shanghai Kou Qiang Yi Xue, 2008, 17(1): 45-50.
[9] 蘇靜芬,張晨,劉云波, 等.恒河猴p53基因沉默靶點在細胞水平的驗證[J].中國比較醫學雜志, 2014, 24(8): 7-10.
[10] Longo PA,Kavran JM,Kim MS,etal.Transient mammalian cell transfection with polyethylenimine (PEI)[J].Methods Enzymol, 2013, 529: 227-240.
[11] Alfonso J,Pollevick GD,Castensson A,etal. Analysis of gene expression in the rat hippocampus using Real Time PCR reveals high inter-individual variation in mRNA expression levels[J].J Neurosci Res, 2002, 67(2): 225-234.
[12] Mosier DE. Small animal models for acquired immune deficiency syndrome (AIDS) research[J]. Lab Anim Sci,1996, 46(3): 257-265.
[13] Schiel N,Souto A.The common marmoset: An overview of its natural history, ecology and behavior[J].Dev Neurobiol, 2017,77(3):244-262.
[14] Watakabe A,Sadakane O,Hata K,etal. Application of viral vectors to the study of neural connectivities and neural circuits in the marmoset brain[J].Dev Neurobiol,2017,77(3):354-372.
[15] Hashikawa T,Nakatomi R,Iriki A. Current models of the marmoset brain[J].Neurosci Res,2015, 93: 116-127.
[16] Chan AW.Progress and prospects for genetic modification of nonhuman primate models in biomedical research[J].ILAR J,2013, 54(2): 211-223.
[17] Bernal M,Ruiz-Cabello F,Concha A,etal. Implication of the beta2-microglobulin gene in the generation of tumor escape phenotypes[J]. Cancer Immunol Immunother, 2012, 61(9): 1359-1371.
Validation of the silencing site of marmosetB2mgene at the cellular level
DENG Yi-chen1,ZHANG Chen2,XIANG Zhi-guang1,TENG Yong-kang1,LIU Yun-bo1*
(1.Institude of Medical Laboratory Animal Science,Chinese Academy of Medical Sciences, Beijing 100021,China;2.State Key Laboratory of Membrane Biology, School of Life Sciences; PKU-IDG/McGovern Institute for Brain Research, Peking University,Beijing 100871)
Objective To screen and determine the effective silencing targets of β2- microglobulin(B2m)gene at the cellular level in marmoset.Methods By homology comparison of the b2m gene in human and theB2mgene in marmoset, choose homology small hairpin RNA(shRNA)sequences targeting marmosetB2mgene were designed, We choose homology small hairpin RNA(shRNA)sequences targeting designedB2mgene to make homology analysis, and insert into lentivirus-based gene silencing constructs FUGW-TDT. The vectors were transfected into HEK293T cells induced by polyethylenimine(PEI). The suppression ofB2mmRNA was detected by real-time PCR. Results Two gene-silencing sequences were screened that lied in 290~310 bp and 665~685 bp of the marmosetB2mmRNA, and have statistical significance in the silencing rate:(46.54±7.91)% (P< 0.05) and(83.22±4.37)%(P< 0.0001). Conclusions Two effective silencing target sequences are screened at cellular level, which can be further used in studies on gene silencing in marmoset.
Gene silence; Marmoset;B2m; shRNA
國家科技支撐計劃(2014BAI03B01)。
鄧怡晨(1991-),女,碩士研究生,研究方向:實驗動物學。E-mail: yichendeng007@126.com
劉云波,教授,研究方向:實驗動物質量控制。E-mail: yunboliu@126.com
R-33
A
1671-7856(2017) 05-0037-05
10.3969.j.issn.1671-7856. 2017.05.010
2016-12-30
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
