食源性致瀉大腸桿菌血清型及毒力基因的研究
2017-06-05 15:10:48余曉豐梅玲玲陳國剛
食品工業科技 2017年9期
余曉豐,楊 勇,占 利,梅玲玲,陳國剛,*
(1.石河子大學食品學院,新疆石河子 832000; 2.浙江省疾病預防與控制中心微生物所,浙江杭州 310051)
食源性致瀉大腸桿菌血清型及毒力基因的研究
余曉豐1,楊 勇2,占 利2,梅玲玲2,陳國剛1,*
(1.石河子大學食品學院,新疆石河子 832000; 2.浙江省疾病預防與控制中心微生物所,浙江杭州 310051)
了解浙江地區食源性致瀉大腸桿菌的血清型分布特點及毒力基因攜帶狀況。收集食品安全風險監測點的致瀉大腸桿菌分離株,采用玻片凝集法對受試菌株進行血清分型,運用多重PCR檢測其攜帶的毒力基因。在69株受試菌株中,優勢血清型包括O148(16株)、O159(11株)、O6(4株)、O15(4株)、O63(4株)和O78(3株),這六種血清型菌株共計42株,占菌株總數的60.9%。菌株主要攜帶的毒力基因為astA(42株)、estIb(25株)、pic(15株)、escV(14株)、aggR(14株);最常見的毒力基因組合為astA+estIb(25株)。進一步分析發現,攜帶astA、estIb基因的O148和O159的菌株對食品安全存在較大威脅。浙江地區食源性致瀉大腸菌的類型以EAEC和ETEC為主,攜帶的主要毒力基因為astA和estIb,優勢血清型為O148和O159。
食源性致瀉大腸桿菌,多重PCR,血清型,毒力基因
大腸埃希氏菌(Escherichiacoli)通常被稱為大腸桿菌,是存在于食物、溫血動物腸道和環境中的常見細菌[1]。大腸菌群數能夠反映食品是否被污染以及被污染的程度,是評價食品衛生質量的重要指標[2]。盡管普通大腸桿菌屬于正常腸道菌群的組成部分,但是獲得特殊毒力因子的大腸桿菌可引起食物中毒,臨床表現為腹瀉和敗血癥,引起腹瀉的這類菌株稱之為致瀉大腸桿菌[3-4]。根據攜帶毒力基因的不同,致瀉大腸桿菌分為:腸粘附性大腸桿菌(EAEC)腸出血性大腸桿菌(EHEC)、腸侵襲性大腸桿菌(EIEC)、腸致病性大腸桿菌(EPEC)和腸產毒性大腸桿菌(ETEC)[5]。
近年來,食源性致瀉大腸桿菌引起的食物中毒事件屢見不鮮,尤其是在發展中國家[6]。其中,飲用水、面制品和肉制品,是致瀉大腸桿菌引起人類食物中毒的主要媒介[7]。一直以來,對這類高危食品中致瀉大腸桿菌進行監測是預防食物中毒事件發生的有效手段[8]。之前關于食品中致瀉大腸桿菌的研究,關注點是食品的衛生狀況,很少有對分離株基因型和表現型的研究。本研究收集浙江地區2014年以來的分離株,運用玻片凝集法和多重PCR,首先對菌株進行血清分型,然后并對其種屬基因(uidA)和毒力基因(pic、bfpB、invE、elt、escV、aggR、stx2、stx1、estIb、astA)進行檢測,最后對其血清型分布特點及毒力基因攜帶狀況進行統計和分析,為食物中毒事件的防控提供數據支撐。
1 材料和方法
1.1 材料和儀器
致瀉大腸桿菌 均由浙江省疾病預防與控制中心從食品安全風險監測點收集的樣本中分離;營養瓊脂培養基 青島海博生物技術有限公司;組織和血液DNA提取試劑盒 德國Qiagen公司;五種致瀉性大腸埃希氏菌多重PCR檢測試劑盒 北京卓誠惠生生物科技股份有限公司;100 bp DNA Ladder 日本TaKaRa公司;瓊脂糖 上海生工;病原大腸菌免疫血清 日本生研。
PCR擴增儀 德國Eppendorf公司;穩壓電泳儀,水平電泳槽,凝膠成像系統 美國Bio-Rad公司。
1.2 實驗方法
1.2.1 DNA模板提取 實驗菌株接種于普通營養瓊脂平板37 ℃培養18 h后,按試劑盒說明書操作步驟提取細菌基因組DNA,并存于-20 ℃凍存備用。
1.2.2 毒力基因的測定多重PCR 采用25 μL的反應體系:2×PCR Buffer 12.5 μL、10×Multiplex Assay 2.5 μL、25×PCR Enzyme 1 μL、Nuclease-free Water 7 μL和DNA模板2 μL。擴增條件:95 ℃預變性4 min;95 ℃變性30 s,62 ℃退火30 s,72 ℃延伸90 s,共30個循環;最后72 ℃延伸5 min[9]。取4 μL擴增產物在含1%瓊脂糖凝膠中以6 V/cm恒壓電泳40 min。用凝膠成像系統拍攝圖片,觀察擴增條帶。在含有uidA條帶的基礎上,根據100 bp DNA Mark和陽性對照判斷擴增條帶大小,進而確定毒力基因種類,結合毒力基因的組合方式最終判定菌株的致瀉類型。
1.2.3 血清分型 實驗菌株接種于普通營養瓊脂平板37 ℃培養18 h,用無菌棉簽挑取單菌落均勻磨到含3 mL無菌生理鹽水的比色管中,使得菌懸液的終濁度為4;然后置于121 ℃高壓2 h,8000 r/min離心10 min后去上清;最后將米粒大小菌體重懸于含500 μL無菌生理鹽水的離心管中備用。取20 μL免疫血清于潔凈載玻片,將20 μL菌懸液滴在血清上混勻,緩慢搖動載玻片30 s后觀察,同時以無菌生理鹽水作為對照。在1 min內,若實驗組明顯凝集現象,對照組無凝集現象,則為實驗陽性;若兩者均出現明顯凝集現象,則該菌株為自凝型(Orough);若兩者未現明顯凝集現象,則為實驗陰性(ONT)。
2 結果與分析
2.1 菌株類型及毒力基因的分布
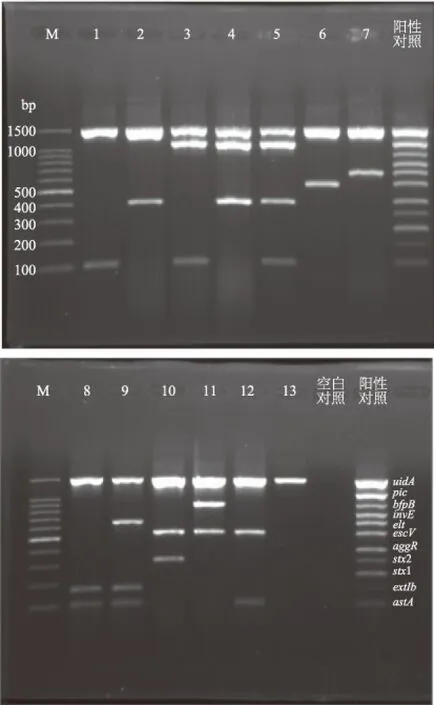
圖1 12種代表性菌株的多重PCR產物凝膠電泳圖Fig.1 Agarose gel electrophoresis of 12 isolates that were selected as the representatives of the strains tested注:泳道1~7對應的毒力基因片段分別為astA、aggR、astA+pic、pic+aggR、astA+pic+aggR、escV、elt;泳道8~12對應的毒力基因片段分別為astA+estIb、astA+elt+estIb、escV+stx2、escV+bfpB、astA+escV;泳道13對應參考菌株ATCC25922的種屬基因片段uidA;M為100 bp DNA Ladder Mark。
在69株致瀉大腸桿菌中,42株astA基因陽性;25株estIb基因陽性;15株pic基因陽性;14株escV基因陽性;14株aggR基因陽性;6株elt基因陽性;1株bfpB基因陽性;1株stx2基因陽性;未檢出invE和stx1基因。攜帶stx1基因的致瀉大腸桿菌通常會導致溫血動物腸道出血,但是這類菌株在食品樣本中陽性率很低。之前李毅等也報道在食品樣本未發現攜帶該毒力基因的菌株,這與本實驗結果一致[10]。此外,根據實驗結果可以得出:浙江地區食源性致瀉大腸菌株主要攜帶的毒力基因是astA;常見的毒力基因組合為astA+estIb,兩者都屬于熱穩定腸毒素基因。由于熱穩定腸毒素分子量小,無免疫原性,這無疑增加了防控的難度。
根據毒力基因的組合方式的不同,實驗菌株可分為EAEC 25株(36.2%)、EPEC 11株(15.9%)、ETEC 5株(7.2%)、EHEC 1株(1.4%)、EAEC和ETEC 25株(36.2%)、EAEC和EPEC 2株(2.9%)、未檢出EIEC。多重PCR產物電泳后共得到12種不同的條帶組合如圖1所示;各種毒力基因組合菌株的種類和數量見表1。由實驗結果可知,根據各類菌株的數量,可以得出EAEC和ETEC是浙江地區食品中的致瀉大腸桿菌的主要類型。盡管EAEC的檢出率高于ETEC,但是由于ETEC易引起嬰幼兒和旅行者腹瀉,更常見于報道。Canizalez等對墨西哥城市錫那羅亞州的5162份食品樣本進行調查后發現當地ETEC陽性率為1.7%,而張淑紅等對國內24個城市559份食品樣本進行研究,結果顯示ETEC的陽性率為6.4%[11-12]。遇曉杰等報道黑龍江地區食源性致瀉性大腸埃希氏菌中EAEC、ETEC、EPEC的占比分別為38.8%、37.5%、15.0%[13]。這些數據表明,EAEC、ETEC和EPEC是目前威脅我國食品安全的主要致瀉大腸桿菌類型。

表1 不同類型食源性致瀉大腸桿菌的毒力基因分布
注:“+”表示相應的基因檢測結果為陽性。

表2 不同類型食源性致瀉大腸桿菌的血清型分布
注:“Orough”表示菌株為自凝型;“ONT”表示血清分型時菌株無凝集現象。
2.2 血清型分布
在69株致瀉大腸桿菌中,O148有16株、O159有11株、O6有4株、O15有4株、O63有4株、O78有3株、以上6種血清型合計42株,占總數的60.9%。11株O159中,81.8%菌株為EAEC和ETEC雜合株中;16株O148中,62.5%的菌株為EAEC,31.3%的菌株為EAEC和ETEC雜合株;血清型的分布特點詳見表2。
實驗結果表明,浙江地區食品中致瀉大腸桿菌的優勢血清型為O148和O159。作為致瀉大腸桿菌的保護性抗原,O-抗原能激發宿主產生抗體從而發揮免疫作用,是疫苗的靶點[14]。由此可以推測,浙江地區市面上針對O148和O159菌株的疫苗效果并不明顯,需要作進一步評估。臨床上關于這兩種血清型陽性分離菌株也有報道,并證實這類菌株會導致感染者腹瀉[15-16]。在日本,這兩種血清型菌株也曾多次引起食物中毒事件的發生,經證實水龍頭和井水是病原體污染食品的主要途徑[17-18]。眾所周知,郊區和偏遠農村的生活用水很少經過殺菌處理,很多農副產品經過簡單清洗處理后直接銷售到城市,一旦被污染,很容易引起集體食物中毒事件,因此,加強這類食品的監測很有必要。此外,在食源性致瀉大腸桿菌的監測中,對這兩種血清型進行針對性鑒定,可以節約時間成本。
2.3 毒力基因與血清型的關系
結合多重PCR和血清分型的實驗結果可知,16株優勢血清型O148陽性菌株的毒力基因分布如下:astA(6株)、astA+estIb(5株)、astA+pic(2株)、astA+pic+aggR(1株)、pic+aggR(1株)、elt(1株);11株O159陽性菌株的毒力基因分別為:astA+estIb(9株),elt(2株)。
O-抗原多糖由細菌染色體上的抗原合成基因簇編碼合成,該基因簇易獲取噬菌體或質粒上抗原修飾基因;在基因的水平轉移過程中,菌株可能同時獲得某種毒力基因[19]。因此,菌株攜帶的毒力基因和血清表型之間存在一定的關聯。本研究發現某些毒力基因集中出現在幾種血清型中;例如,陽性率較高的astA、estIb基因主要出現在O148和O159菌株中。之前也有研究表明大腸桿菌的攜帶的毒力基因和血清表型之間存在密切的聯系[20-21]。盡管如此,并不能根據菌株攜帶的毒力基因直接推測出該菌株的血清型;例如本研究中,攜帶estIb基因的兩株菌,血清表型可能一致(同為O148或同為O159),也可能不一致(一株為O159,另一株為O148)。由此可知,毒力基因會集中出現在幾種血清型中,但是不存在嚴格對應關系。可能是由于抗原多糖由多個基因協同編碼合成,存在其他未知基因的調控[22]。
3 結論
浙江地區食源性致瀉大腸菌的類型以EAEC和ETEC為主;攜帶的主要毒力基因為astA(60.9%)和estIb(36.2%);優勢血清型為O148(23.2%)和O159(15.9%);未發現攜帶invE、stx1基因的菌株。盡管毒力基因astA在血清型O148、O159菌株中檢出率高達87.5%、81.8%,但是兩者不存在嚴格對應關系。
[1]姬華,張玫,盧士玲,等.食源性大腸桿菌耐藥性與毒力特征的研究進展[J].食品工業科技.2014,35(07):364-367.
[2]武娟,張祺,李丹,等.青島市售雙殼貝類大腸菌群/大腸桿菌檢出水平與當地氣溫的相關性[J].食品工業科技.2014,35(17):147-150.
[3]Hemmatinezhad B,Khamesipour F,Mohammadi M,et al.Microbiological investigation of O-serogroups,virulence factors and antimicrobial resistance properties of shiga toxin-producingEscherichiacoliisolated from Ostrich,Turkey and Quail meats[J]. Journal of Food Safety.2015,35(4):491-500.
[4]Da S S A,Gomes R F,Yukie S L,et al.Escherichiacolifrom Crohn’s disease patient displays virulence features of enteroinvasive(EIEC),enterohemorragic(EHEC),and enteroaggregative(EAEC)pathotypes[J].Gut Pathogens.2015,7(1):2.
[5]Levin R E.Molecular methods for detecting and discriminating Shigella associated with foods and human clinical infections-A review[J].Food Biotechnology.2009,23(3):214-228.
[6]Gould L H,Walsh K A,Vieira A R,et al.Surveillance for foodborne disease outbreaks-United States,1998-2008[J]. Morbidity and Mortality Weekly Report.Surveillance Summaries. 2013,62(2):1-34.
[7]Macdonald E,Moller K E,Wester A L,et al.An outbreak of enterotoxigenic Escherichia coli(ETEC)infection in Norway,2012:a reminder to consider uncommon pathogens in outbreaks involving imported products[J].Epidemiology and Infection.2015,143(3):486-493.
[8]郭玉梅,秦麗云,徐保紅,等.2003至2012年石家莊地區細菌性食物中毒病原菌特征研究[J].微生物學雜志.2013,33(5):73-77.
[9]Muller D,Greune L,Heusipp G,et al.Identification of unconventional intestinal pathogenicEscherichiacoliisolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR[J].Applied and Environmental Microbiology.2007,73(10):3380-3390.
[10]李毅,章樂怡,洪程基,等.溫州市食品中腸出血性大腸桿菌O157∶H7污染狀況調查[J].中國食品衛生雜志.2012,24(4):369-371.
[11]Canizalez-Roman A,Gonzalez-Nunez E,Vidal J E,et al.Prevalence and antibiotic resistance profiles of diarrheagenicEscherichiacolistrains isolated from food items in northwestern Mexico[J].International Journal of Food Microbiology.2013,164(1):36-45.
[12]Zhang S,Wu Q,Zhang J,et al.Prevalence,genetic diversity,and antibiotic resistance of enterotoxigenicEscherichiacoliin retail ready-to-eat foods in China[J].Food Control.2016,68:236-243.
[13]遇曉杰,閆軍,薛成玉,等.黑龍江省食品風險監測致瀉性大腸埃希氏菌致病性分析[J].中國公共衛生管理.2016,02(32):250-252.
[14]潘超,朱力,王恒樑.病原細菌多糖疫苗和多糖結合疫苗研究進展[J].生物技術通訊.2013,24(05):705-709.
[15]羅淑華,黃鵬飛,李燕,等.產毒性大腸埃希菌血清型與毒力流行特征及耐藥分析[J].中國衛生檢驗雜志.2013,23(09):2176-2179.
[16]Hussain M I,Wani S A,Nagamani K,et al.Isolation and characterization of enteroaggregative,enterotoxigenic,diffusely adherentEscherichiacoliand Salmonella Worthington from human diarrhoeic faecal samples in Kashmir and Secunderabad,India[J]. World Journal of Microbiology and Biotechnology.2010,26(10):1883-1889.
[17]Konishi N,Obata H,Monma C,et al.Bacteriological and epidemiological characteristics of enterotoxigenicEscherichiacoliisolated in Tokyo,Japan,between 1966 and 2009[J].Journal of Clinical Microbiology.2011,49(9):3348-3351.
[18]Konishi N,Obata H,Shimojima Y,et al.Colony-sweep PCR efficacy in food-borne outbreak associated with six types of enterotoxigenicEscherichiacoli[J].The Journal of the Japanese Association for Infectious Diseases.2009,83(5):490-495.
[19]蔡成松,郭曉奎.細菌O抗原基因簇的研究進展[J].微生物學雜志.2008,28(03):64-67.
[20]Rodas C,Mamani R,Blanco J,et al.Enterotoxins,colonization factors,serotypes and antimicrobial resistance of enterotoxigenicEscherichiacoli(ETEC)strains isolated from hospitalized children with diarrhea in Bolivia[J].Brazilian Society of Infectious Diseases.2011,15(2):132-137.
[21]陳祥,趙李祥,高崧,等.豬源大腸桿菌(ETEC、STEC、AEEC)毒力基因及其與O抗原型的關系[J].微生物學報.2008,48(7):857-862.
[22]劉璨穎,張濟培,王丙云,等.大腸桿菌O-抗原血清型鑒定研究進展[J].中國人獸共患病學報.2016,32(10):928-933.
Study on serotypes and virulence genes of foodborne diarrheogenicEscherichiacoli
YU Xiao-feng1,YANG Yong2,ZHAN Li2,Mei Ling-ling2,CHEN Guo-gang1,*
(1.College of Food,Shihezi University,Shihezi 832000,China; 2.Department of Microbiology,Zhejiang Provincial Center for Disease Control and Prevention,Hangzhou 310051,China)
The main purpose of the research is to define serotypes and virulence factors of foodborne diarrheogenicEscherichiacoliin Zhejiang Province. The distribution characteristics of serotypes and virulence factors of strains collected from food surveillance were detected by serotyping and multiple polymerase chain reaction(PCR).Of all 69 strains,the most frequent identified serotypes were O148(16),O159(11),O6(4),O15(4),O63(4)and O78(3),the total number of these serotypes was 42,which accounted for 60.9% of the strains. And 42,25,15,14,14 strains were positive forastA,estIb,pic,escV,aggRgene,respectively. Among these,25 strains harbored bothastAandestIbgenes,representing the common type. Furthermore,O148 and O159 strains harboringastAandestIbgenes posed a potential threat to food safety. Results showed that EAEC and ETEC harboringastAandestIbgenes were the dominant bacteria types,mainly belonging to serotype O148 and O159,among foodborne diarrheogenicEscherichiacoliin Zhejiang Province.
foodborne diarrheogenicEscherichiacoli;multiplex polymerase chain reaction;serotypes;virulence genes
2016-11-14
余曉豐(1990-),男,碩士,研究方向:食源性致病微生物,E-mail:yuxiaofeng@foxmail.com。
*通訊作者:陳國剛(1977-),男,博士,教授,研究方向:食品微生物及果蔬貯藏,E-mail:cgg611@163.com。
國家自然基金(31560468);中國博士后科學基金(2015M570525)。
TS201.1
A
1002-0306(2017)09-0124-04
10.13386/j.issn1002-0306.2017.09.015
