歐美楊細菌性潰瘍病菌雙組分信號轉導系統LqHK1基因的功能*
2017-06-05 09:07:55李愛寧
林業科學 2017年4期
魏 強 李愛寧 賀 偉
(北京林業大學林學院 北京 100083)
?
歐美楊細菌性潰瘍病菌雙組分信號轉導系統LqHK1基因的功能*
魏 強 李愛寧 賀 偉
(北京林業大學林學院 北京 100083)
【目的】 明確歐美楊細菌性潰瘍病病原菌中雙組分信號轉導系統LqHK1基因的生物學功能,以闡明該病原菌的致病機制。【方法】 采用生物信息學方法鑒定雙組分系統LqHK1,利用同源重組方法獲得該基因缺失突變體菌株△LqHK1,分析突變體及其互補菌株的生長速率、游動性、生物膜形成、致病性等生物學特性。同時,采用qRT-PCR檢測與病原菌游動性相關的基因flgB、flgC、flgE的表達水平,并定量分析接種病原菌后在接種點部位楊樹組織內病原菌的DNA含量。【結果】 鑒定出病原細菌的雙組分系統基因LqHK1,通過同源重組獲得缺失突變體△LqHK1。表型分析顯示: △LqHK1在1年生107楊莖干上的致病力顯著下降; 突變體在寄主體內定殖的菌量明顯少于野生型菌株; 突變體菌株的游動能力比野生型顯著降低,游動性相關基因flgB、flgC、flgE在突變體中的表達量顯著下降;LqHK1突變體生物膜的形成能力顯著降低,生長速率與野生型無顯著差異。【結論】LqHK1基因缺失突變體在107楊上的致病能力顯著下降,歐美楊細菌性潰瘍病病原菌雙組分信號轉導系統LqHK1基因與病原菌的致病性密切相關。
歐美楊細菌性潰瘍病病原菌; 雙組分信號系統; 致病性
歐美楊細菌性潰瘍病是由歐美楊細菌性潰瘍病原菌(Lonsdaleaquercinasubsp.populi)引起的一種毀滅性林木病害,國內于2005年首次在河南省濮陽市發現(郭利民等, 2008)。該病主要在中林46 楊(Populus×euramericanacv.‘Zhonglin 46’)、107 楊(P. ×euramericanacv. ‘74/76’)等歐美楊品種莖干上危害,造成樹木病部枝條和樹干頂梢枯死,影響樹木的健康生長,并導致樹木材積和其他可利用部分減少(Lietal., 2014)。目前對于該病害防治尚未得到有效解決。
微生物是地球上最多變、適應性最強的有機體,為了適應多樣的生態環境,必須具備一套復雜而精密的信號系統來應答外界或體內環境的變化,從而適應環境并得以生存。目前已發現多種細胞信號轉導途徑,其中蛋白質磷酸化信號轉導途徑廣泛存在于各種微生物中 (Mizuno, 1997; 王梁燕, 2008)。雙組分信號轉導系統是Ninfa等(1986)在研究大腸桿菌(Escherichiacoli)氮調節蛋白系統時首次發現的,是廣泛存在于原核和真核細胞中的信號轉導系統,主要由具有保守的組氨酸位點的組氨酸激酶(histidine kinase,HK)和與其偶聯的反應調節蛋白(response regulator,RR)2個基本組分組成。該系統是細菌對外界各種環境信號刺激做出反應的一個重要機制,通常組氨酸激酶(HK)是一個跨膜蛋白,以檢測環境變化,在外界環境刺激下,組氨酸激酶能迅速感受到環境刺激,并催化其自身磷酸化,隨后磷酸基團從高能磷酸化的殘基轉移至RR接收區,接收區則通過磷酸化調節輸出區的活性,以此來調控目的基因表達或使目標蛋白的功能發生適應性變化(Hoch, 2000; 單世平等, 2014)。
雙組分信號轉導系統參與調解細胞分裂、控制病原菌的致病性、抗逆性、雙氧水的耐受性、滲透壓調節等,尤其是在不利的生存環境中起著不可或缺的作用。因此,為了明確歐美楊細菌性潰瘍病病原菌中雙組分信號轉導系統在致病過程中的作用,通過該病原菌全基因組生物信息學檢索,得到16個組氨酸激酶基因序列,采用基因敲除方法獲得1個有表型的基因,將其命名為LqHK1,并對此基因進行功能研究,分析該基因對歐美楊細菌性潰瘍病病原菌的生長速率、生物膜、游動性及致病性等方面的影響,以期為探索歐美楊潰瘍病病原菌的致病機制和信號轉導奠定基礎。
1 材料與方法
1.1 供試菌株、質粒及培養條件
本研究所用菌株、質粒及性狀見表1。大腸桿菌用LB培養基37 ℃培養16 h; 歐美楊細菌性潰瘍原菌N-5-1菌株及其衍生菌株用LB培養基30 ℃培養24 h。所用抗生素終濃度分別為氨芐霉素(AmpR)100 μg·mL-1、氯霉素(CmR) 20 μg·mL-1、慶大霉素(GmR) 30 μg·mL-1、利福平(RifR) 20 μg·mL-1。所用引物在天一輝遠(北京)公司合成。

表1 本研究所使用菌株和質粒①Tab.1 Bacterial strains and plasmids used in this study
①AmpR、CmR、GmR、RifR分別表示氨芐霉素、氯霉素、慶大霉素、利福平抗性。 AmpR,CmR,GmR,RifRrepresent resistance to ampicillin, chloramphenicol, gentamicin and rifampicin, respectivly.
1.2 供試植物
107楊樹苗為采自北京林業大學森保研究室苗圃地的1年生扦插苗。選取相同生長狀況的樹苗,截取30 cm長枝段,每個菌株接種5個枝段,重復3次。
1.3L.quercinaLqHK1敲除突變體的構建
基因敲除采用同源重組方法(圖1),根據LqHK1基因序列設計引物(表2)。使用LqHK1-UF/LqHK1-UR、LqHK1-DF/LqHK1-DR 2對引物從菌株L.quercinaN-5-1基因組中分別擴增LqHK1基因的上、下游同源臂。將純化后的同源臂片段按1∶1混合作為模板,以LqHK1-UF/LqHK1-DR 引物,通過SOE PCR將同源臂連接起來。用Xho I/NotⅠ雙酶切同源片段和自殺質粒pWM91。將酶切后的自殺質粒和融合片段按適當比例混合,加入T4連接酶連接過夜,再將連接產物熱激轉化到E.coli-S17感受態細胞中,構建一個兩側具有LqHK1基因上、下游同源臂的重組自殺性質粒pWM91-ΔLqHK1,并進行雙酶切和測序驗證。
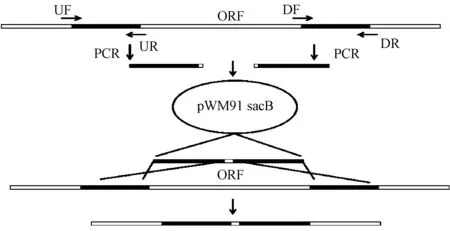
圖1 基因敲除突變體示意Fig.1 Schematic map of the constructions of gene knockout mutants
1.4L.quercinaLqHK1基因敲除突變體的獲得
以攜帶重組自殺質粒pWM91-LqHK1的E.coli-S17為供體菌、L.quercinaN-5-1為受體菌進行自殺性質粒的接合轉移。分別將培養至對數生長期的受體菌與供體菌等體積混勻后,滴加到鋪有LB瓊脂平板表面的無菌微孔濾膜上,37 ℃放置1 h后,轉到30 ℃放置48 h。用LB洗下濾膜上菌液,涂布雙抗性LB瓊脂平板(100 μg·mL-1Amp, 20 μg·mL-1Rif),30 ℃培養72 h,長出的菌落用N-5-1菌株特異性引物Infb238/Infb579(申請專利)和自殺載體上的SacB基因引物SacB1/SacB2作PCR驗證。驗證的菌落轉到含有20 μg·mL-1Rif 5 mL的LB試管中連續搖培,7天后將稀釋1 000倍的100 μL菌液涂到含有5%蔗糖的LB瓊脂平板上,30 ℃培養48 h,各挑取20個有蔗糖耐性的菌落分別點到含有20 μg·mL-1Rif的LB瓊脂平板和含100 μg·mL-1Amp的LB瓊脂平板上,在含有Rif 的LB平板上能生長而在含有Amp的LB平板上不能生長的菌株為缺失突變株。然后用引物LqHK1-UF和LqHK1-DR進行PCR驗證(王薇等, 2014)。
1.5LqHK1 基因互補突變體的獲得
以野生菌株N-5-1基因組為模板,PCR擴增出互補片段,與穿梭質粒pBBR1M-CS-5共同用HindⅢ和XbaⅠ限制性內切酶進行酶切后,將供體菌、受體菌、Helper 菌株取出適量進行三親雜交,將構建正確的互補載體轉化到LqHK1突變菌株中,PCR驗證轉化子,得到互補菌株HBLqHK1。互補菌株的構建可以保證通過回補后表型的恢復來確定該基因的功能(李賓等, 2015)。

表2 本研究所用引物 Tab.2 Oligonucleotides used in this study
1.6 生長曲線測定
將野生菌株、缺失突變體和互補菌株接種于LB液培養基中過夜培養至穩定生長期,用新鮮LB液體培養基將各菌株稀釋到 OD600=0.5,以體積比1∶100的比例將濃度相同的各菌株接種于100 mL的LB液體培養基中,30 ℃、200 r/min 搖培,每隔12 h取1次樣,每個菌株3次重復,測定各菌株在600 nm處的吸光值,繪制生長曲線。
1.7 游動性的測定
將野生菌株、缺失突變體和互補菌株接種于LB液體培養基中過夜培養至穩定生長期,吸取10 μL滴于含有0.3%瓊脂的半固體LB培養基上無菌風吹干,30 ℃靜置培養48 h,測量各菌株游動暈圈直徑,重復3次(Khajanchietal., 2009)。
1.8 生物膜的測定
將野生型菌株N-5-1、△LqHK1和HBLqHK1接種于LB液體培養基中過夜培養至穩定生長期,以體積比1∶100稀釋到含有3 mL新鮮的LB液體培養基的試管中(15 mL),30 ℃靜置培養48 h,將培養液緩慢倒掉,加入0.1%的結晶紫染色15 min,用移液器輕輕吸盡結晶紫溶液,然后用無菌水清洗3遍試管,于37 ℃烘干試管,可觀察到試管壁上一圈紫色痕跡即是固液交界處形成的堅固的生物膜。加入2.5 mL 95% 的乙醇充分溶解與生物膜結合的結晶紫,取1 mL測量OD570(Morohoshietal., 2007)。
1.9 致病性反應測試
將野生菌株、突變體菌株和互補菌株分別接種到含有相應抗生素的LB液體培養基中,30 ℃、200 r·min-1搖床震蕩培養24 h,用無菌水將菌液濃度稀釋到107cfu·mL-1。用自來水清洗歐美楊107枝段,然后用脫脂棉加75%酒精在接種點處進行表面消毒,再用消毒的刀片劃十字傷口,大小為1 cm×1 cm,深度達到木質部,分別用注射器吸取80 μL各菌株菌液接種到枝段傷口處。培養條件: 接種好的枝條放在水桶中,再放入溫濕度氣候箱中(30 ℃、RH80%),接種3天后去掉保鮮膜和棉團。每個菌株接5個枝段,每個枝段接1個點,重復3次,LB液體培養基接種為陰性對照。培養7天后觀察、記錄發病情況(Yangetal., 2014)。
1.10 熒光定量PCR檢測發病部位病原菌含量
1.10.1 歐美楊接種枝條總DNA的提取 按照1.9的方法,接種歐美楊枝條,在發病達到高峰期,第7天,取接種點處100 mg楊樹組織。將組織剪碎,在液氮冷凍下研磨成粉末狀,按照天根植物基因組提取試劑盒的方法提取總DNA。
1.10.2 歐美楊組織中病原菌DNA的Real-time PCR定量分析 應用特異性引物InfB 238/InfB 579擴增L.quercinaDNA,使用ABI 7500 system(Applied Biosystem,Carlsbad,CA,USA)進行Real- time PCR擴增。從LB液體培養基的細菌中提取野生型基因組DNA,用nano-drop 2000定量并10倍梯度稀釋后,用引物InfB 238/InfB 579進行定量PCR,制作標準曲線。根據標準曲線測定樣品中的DNA質量濃度 (Shangetal., 2015)。
1.11 游動性相關基因表達量分析
采用TaKaRa Total RNA 提取試劑盒的方法提取各菌株總RNA。經電泳檢測質量后按照反轉錄試劑盒Prime ScriptTM RT Master Mix(for Real Time)(TaKaRa)的方法合成cDNA。以cDNA為模板,進行實時定量PCR反應,定量PCR反應采用Super Real Premix Plus(天根,中國)、SYBR Green染料和ABI7500實時PCR系統(Applied Biosystem,Carlsbad,CA,USA)。以16S rRNA作為內參基因(引物序列見表2)。每個反應進行3個平行試驗,每個試驗重復3次。相對表達量是由2-△△CT方法計算(Livaketal., 2001)。
2 結果與分析
2.1LqHK1基因缺失突變體的獲得與驗證
以引物LqHK1-UF和LqHK1-DR 進行PCR擴增菌株 N-5-1 和突變體的基因組DNA,電泳檢查證明缺失突變體LqHK1的堿基大小為1 488 bp(圖2A)。
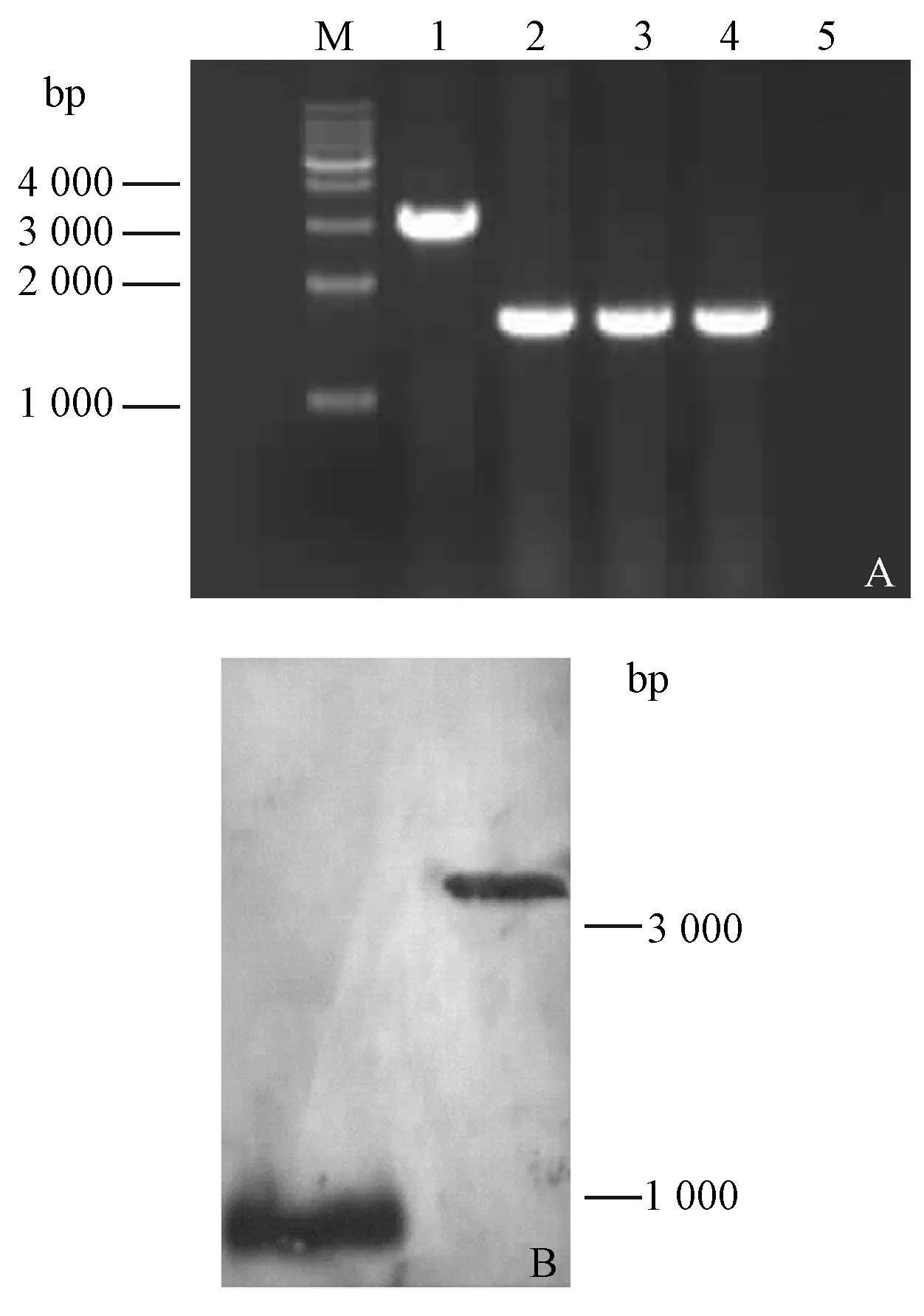
圖2 PCR驗證缺失突變體(A)、Southern blot 驗證突變體(B)Fig.2 PCR analysis of the nutant(A) and Southern blot analysis of the mutant(B) A.M.1 kb maker; 1. N-5-1; 2. LqHK1; 3. LqHK1; 4. H2O.
提取細菌基因組,分別用EcoRI和NotI雙酶切突變體和野生型基因組DNA,以LqHK1-UF/LqHK1-UR擴增產物為探針,進行Southern雜交驗證(圖2B)。由于LqHK1突變體比野生型缺少了1 350 bp,野生型雜交信號帶大小為2 298 bp,而突變體菌株雜交信號帶大小為948 bp,證明確實是突變體。
2.2LqHK1基因互補突變體的獲得與驗證
挑取轉化子,用引物InfB 238/InfB579、引物pBBR1MCS-5多克隆位點上下游引物MCS1/MCS2(圖3)進行PCR驗證。分別可擴增到342 bp和2 182 bp片段,大小與預期一致,即LqHK1基因互補菌株構建正確。
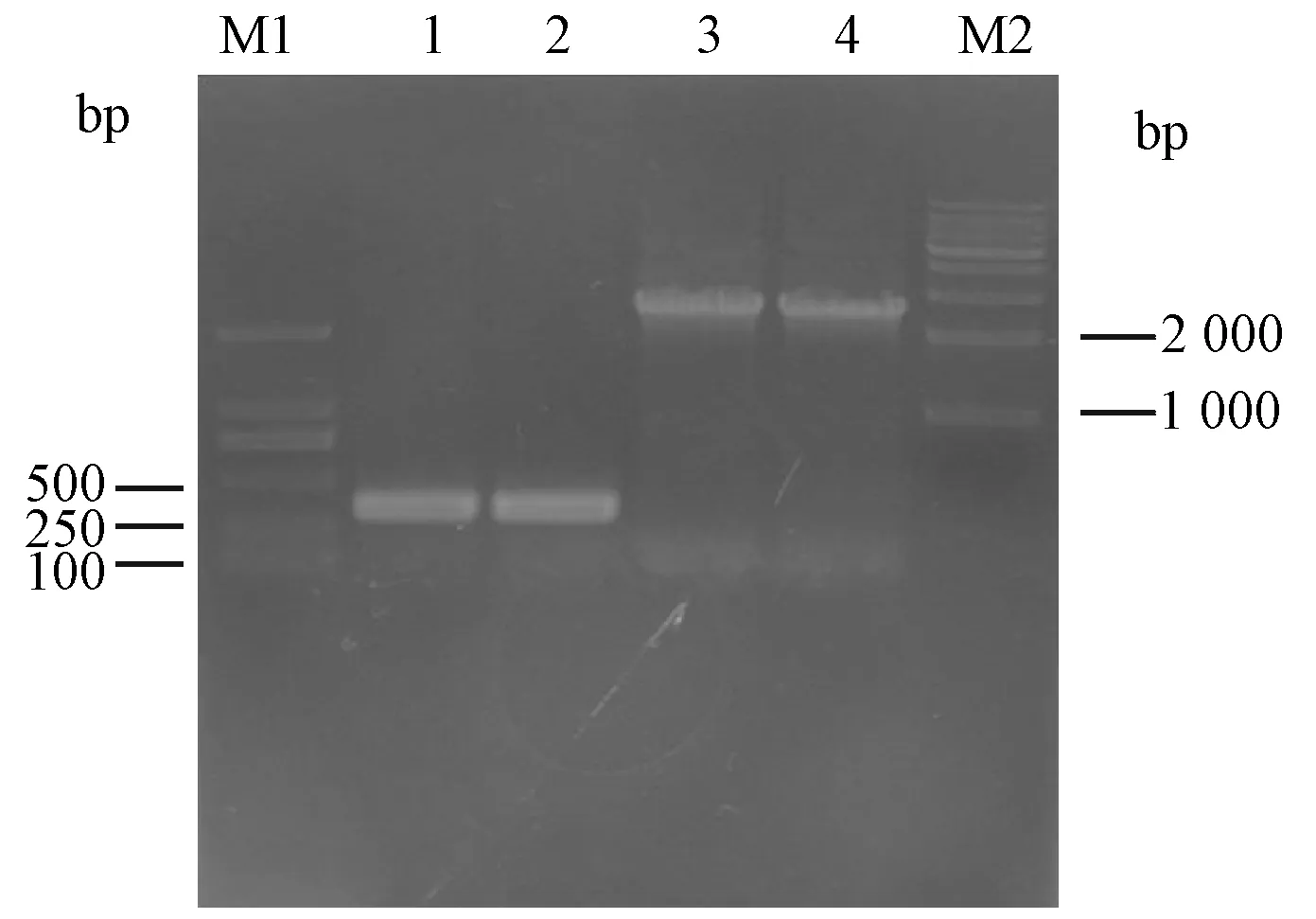
圖3 PCR驗證互補菌株Fig.3 PCR analysis of the complementationM1. D2000; 1. 引物InfB 238/InfB579; 2. 引物InfB 238/InfB579; 3. 引物 MCS 1/MCS 2; 4. 引物 MCS 1/MCS 2; M2. 1 kb maker.
2.3LqHK1缺失突變體的生長速率
以橫坐標為時間、縱坐標為活菌數的對數(OD600值)繪制出一條生長曲線,結果顯示, 野生型菌株N-5-1、△LqHK1 和HBLqHK1的生長曲線無明顯差異,在液體LB培養基中均可正常生長(圖4)。
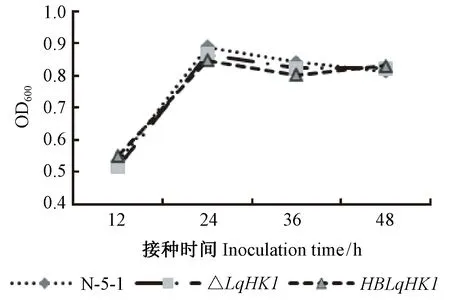
圖4 突變體生長能力的測定Fig.4 Growth test of the mutants
2.4LqHK1基因突變影響生物膜的形成
生物膜是指附著于惰性物體或活動實體表面的細菌包體以及由自身分泌的含水聚合性基質包裹所形成的被膜狀細菌菌落(宋菲等, 2010)。通過結晶紫染色,可以在試管壁上觀察到紫色環狀的生物膜形成。在本試驗中,△LqHK1、HBLqHK1、野生菌株N-5-1均可形成生物膜(圖5A),但突變體形成生物膜的能力顯著降低,OD570的吸光值平均為0.33,明顯小于野生型菌株OD570平均吸光值0.42(P<0.05),對應的互補菌株恢復野生型狀態,說明L.quercinaN-5-1的LqHK1基因在其生物膜形成過程中發揮重要作用(圖5B)。
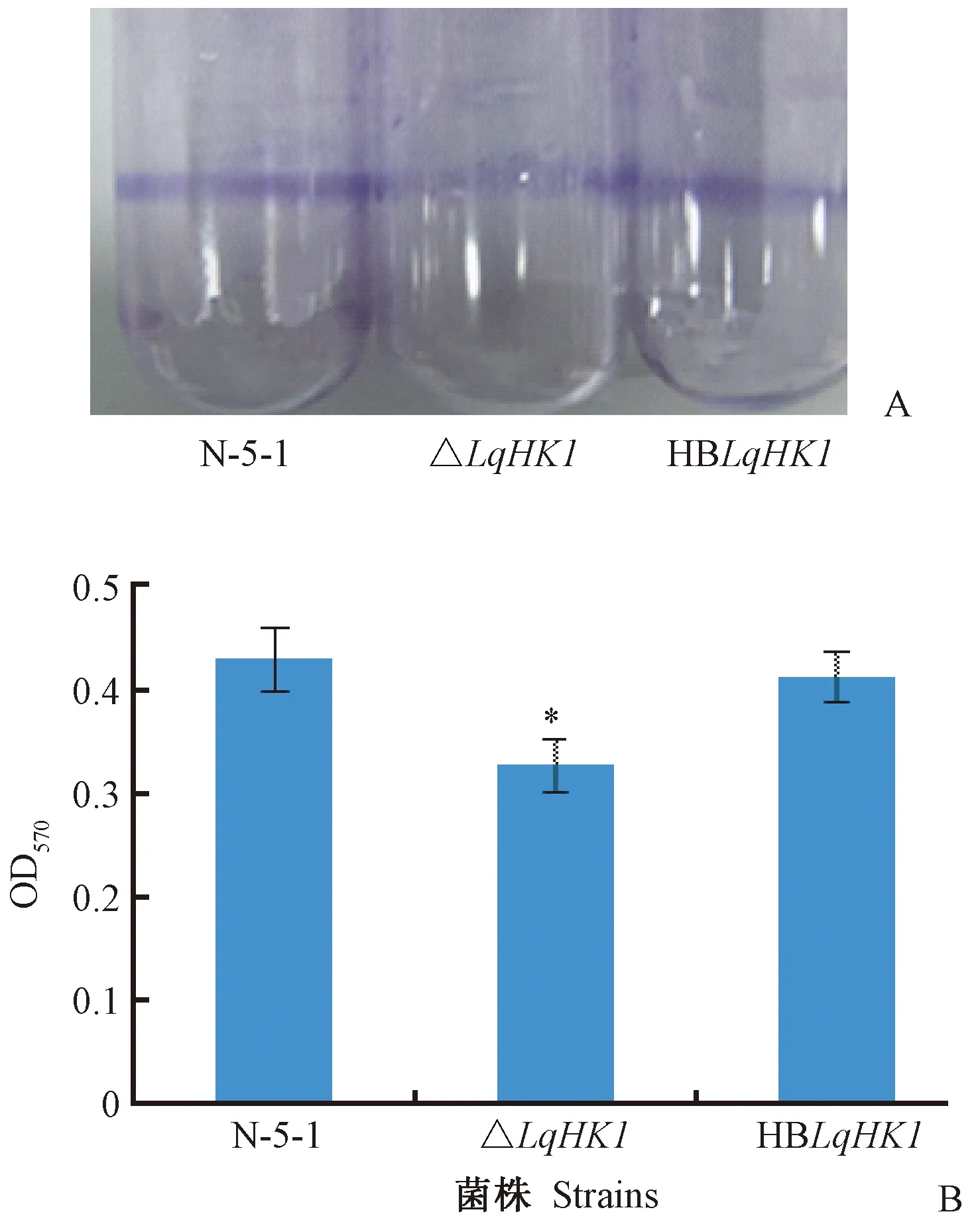
圖5 LqHK1基因突變體對生物膜形成的影響Fig.5 Effect of the LqHK1 mutants on biofilm formationA. 各菌株在試管壁上形成的生物膜Biofilm of strains on the test tube wall;B. 各菌株生物膜的OD570值OD570 of biofilm of strains.
2.5LqHK1缺失影響病原菌游動性及相關基因的表達
經過48 h培養,歐美楊細菌性潰瘍病原菌野生型菌株的游動圈平均直徑為3.54 cm,突變株△LqHK1的游動圈平均直徑為2.51 cm,互補菌株的游動圈平均直徑為3.50 cm(圖6)。數據結果顯示,突變株與與野生型N-5-1的游動能力存在顯著差異(P<0.05),HBLqHK1游動圈大小與野生型無顯著差異,運動能力基本恢復,表明LqHK1在病原菌游動性中具有重要作用。
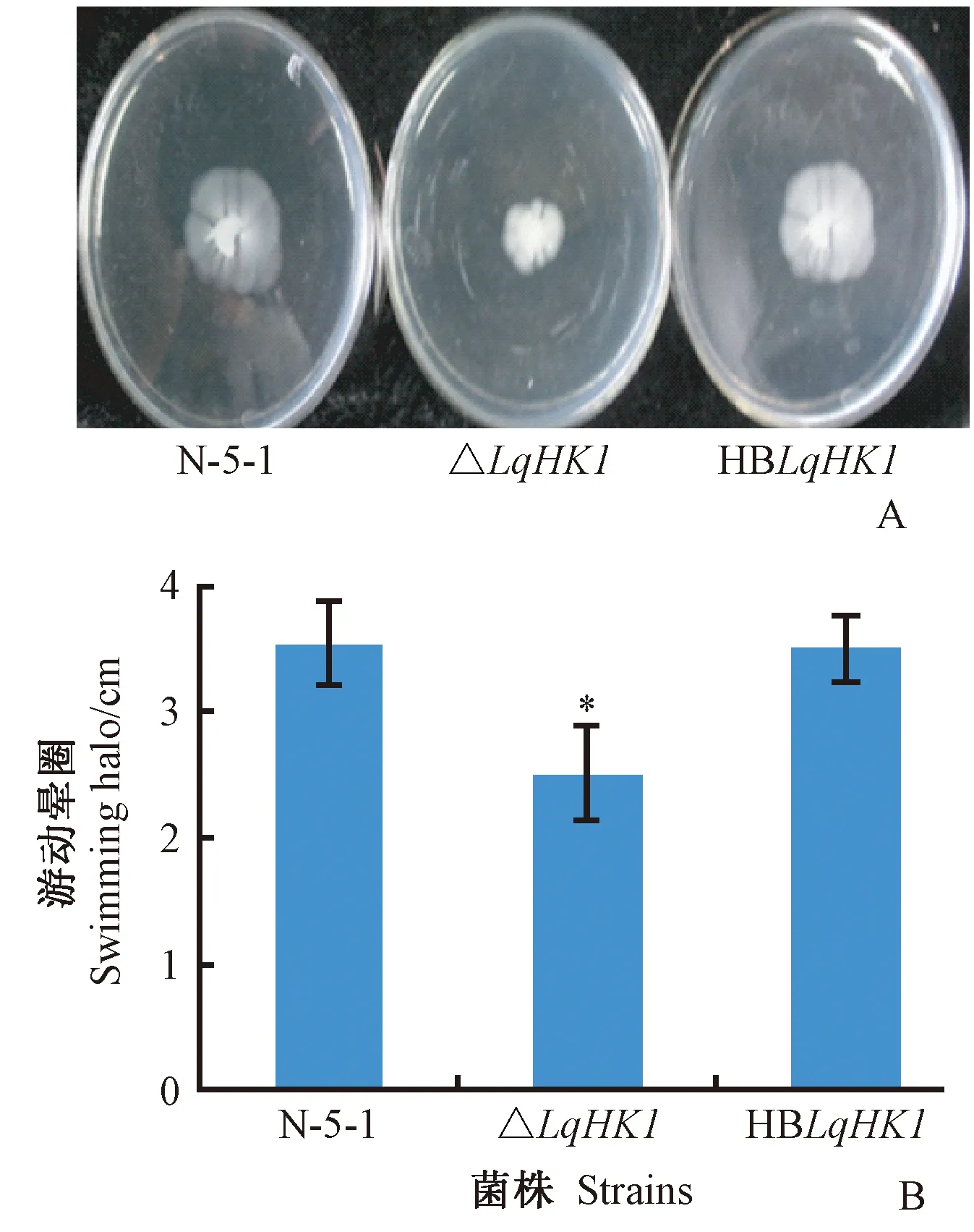
圖6 LqHK1基因突變體的游動能力測定Fig.6 Motility assay of LqHK1mutantsA. 各菌株在半固體培養基中的菌落 The colonies of strains formed on semi-solid medium;B. 菌株的游動暈圈直徑 Diameter of the halos of strains formed.
利用qRT-PCR方法進一步檢測游動性相關基因flgB、flgC、flgE在突變菌株中的表達情況。野生菌株的表達水平設為1。分析表明,游動性相關基因的表達量與野生型菌株相比存在顯著差異(P<0.01)(圖7),說明LqHK1對所選游動性基因為正調控。
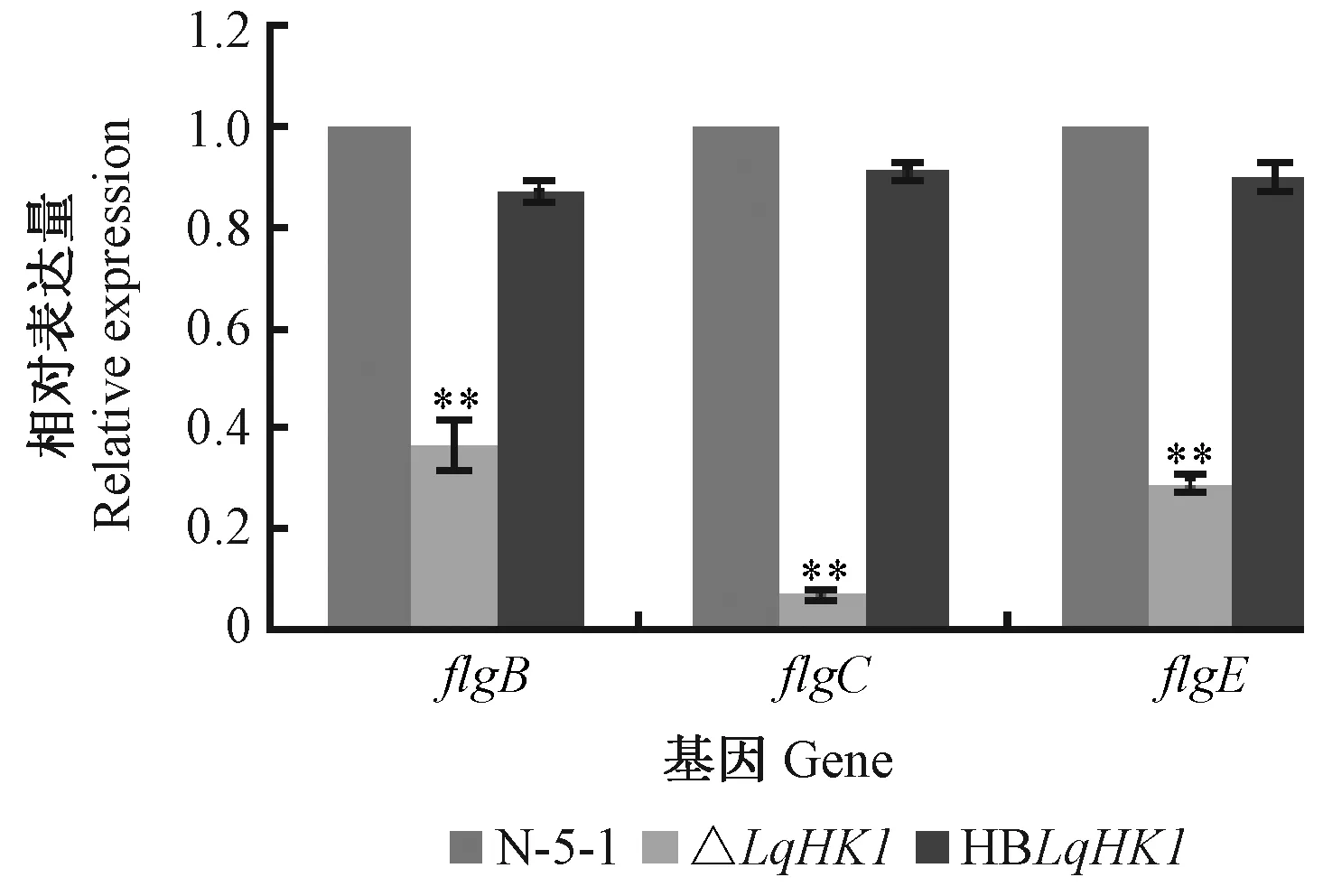
圖7 qRT- PCR分析flgB、flgC、flgE 基因表達量Fig.7 qRT-PCR analysis for the expression of flgB,flgC and flgE genes
2.6LqHK1基因突變體對致病性的影響
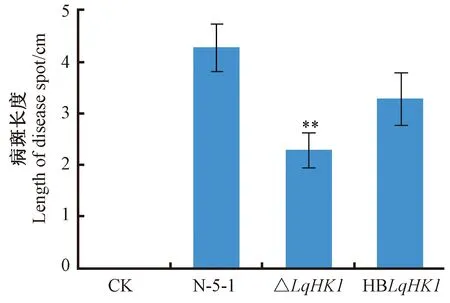
圖8 楊樹枝段接種野生菌株N-5-1、△LqHK1、HBLqHK1后的病斑長度Fig.8 Lesion length on the poplar branches inoculated with wild type strain of L. quercina N-5-1
2.6.2 突變體對病原菌在寄主植物上的定殖能力的影響 將歐美楊潰瘍病病原菌2 ng DNA進行5次10倍梯度稀釋,用引物InfB 238/InfB579進行Real-time PCR分析,制作標準曲線。線性方程為y=-3.605 9x+17.057(R2=0.997 8,P<0.05)。其中,y為CT值,x為細菌性潰瘍病菌DNA濃度常用對數Lg值。
應用Real-time PCR定量分析(圖10)表明,發病達到高峰期時,在接種點部位△LqHK1突變體的DNA含量明顯少于野生菌株 N-5-1和互補菌株,說明在寄主植物上LqHK1基因的缺失導致病原菌的定殖能力下降。
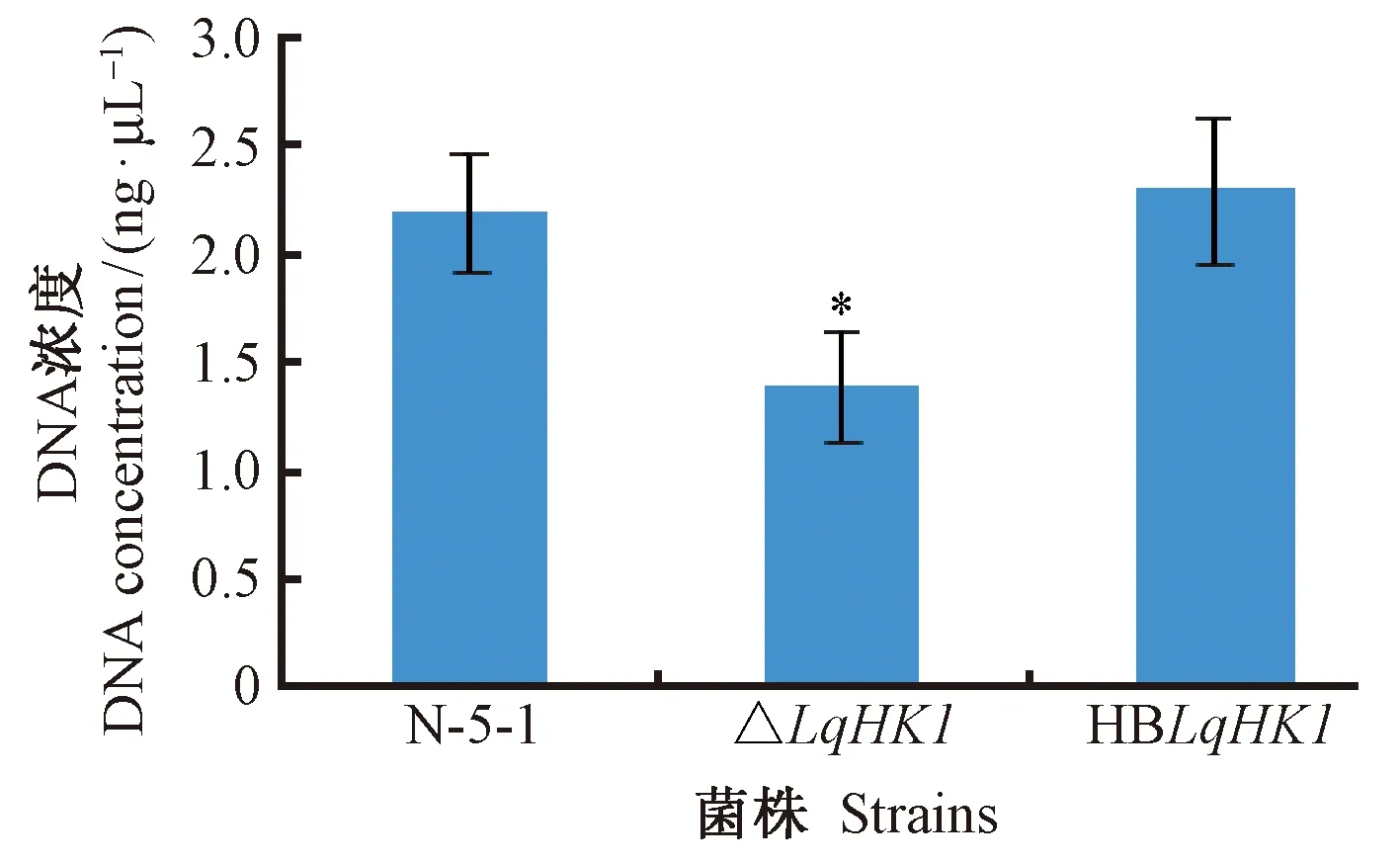
圖9 發病高峰期在接種點處病原菌的Real-time PCR定量分析Fig.9 RT-PCR quantitative analysis of pathogen on inoculation points in the peak period of incidence
3 討論
由歐美楊細菌性潰瘍病原菌引起的歐美楊細菌性潰瘍病是楊樹上一種嚴重的細菌性病害,目前,對于該病原菌的研究主要集中在病原菌鑒定(任飛娟, 2011)、病原菌檢測方法和技術(商靖, 2014)、病原菌的Ⅲ型分泌系統(type Ⅲ secretion system,T3SS)(楊莉等, 2014)等方面。本研究以歐美楊細菌性潰瘍病病原菌雙組分信號系統的LqHK1基因為對象,從野生型菌株N-5-1的全基因組中克隆到相應的雙組分系統基因,然后通過同源重組方法構建了LqHK1基因缺失突變體。通過對L.quercina雙組分信號轉導系統中LqHK1基因的敲除,使突變體菌株在寄主植物上的致病能力降低,且病原菌的游動能力和生物膜形成能力減弱。這說明雙組分系統中的LqHK1基因可對歐美楊細菌性潰瘍病原菌的游動能力、生物膜形成能力造成影響,進而導致病原菌的致病能力減弱,為揭示歐美楊潰瘍病原菌的致病機制和信號轉導提供了試驗依據。并且通過對鞭毛相關基因的表達量檢測,發現LqHK1基因對游動性相關基因起正調控作用,這也在轉錄水平對上述結論進行了進一步印證,從而證明該病原菌中雙組分信號轉導系統發揮著重要的作用。
在許多植物病原細菌中,雙組分調控系統已被證明在病原菌的侵入和致病性等方面起重要作用,如根癌土壤農桿菌(Agrobacteriumtumefaciens)中調控致病性的ChvG/Chvl和VirA/VirG (Charlesetal., 1993; Panetal., 1993)、軟腐病菌(Dickeyadadantii)中,導致寄主植物馬鈴薯(Solanumtuberosum)和胡蘿卜(Daucuscarotavar.sativus)致病性完全喪失的CpxA/CpxR(Bontempsetal., 2015)、梨火疫病菌(Erwiniaamylovora)中EnvZ-OmpR和GrrA-GrrS調節病原菌致病毒力的釋放和致病力(Zhaoetal., 2009)等。在有的病原細菌中,雙組分信號轉導系統對病原菌的其他方面也存在影響,水稻條斑病菌(Xanthomonasoryzaepv.oryzicola)中ColR/ColS的突變,不僅導致病原菌的致病性喪失,而且ColR/ColS也在脂多糖產生、過氧化氧酶活性、環境耐受力(如苯酚、銅和過氧化氫)等方面起著重要作用(Yangetal., 2007)。在本研究中,LqHK1基因并未對病原菌的生長速率、胞外多糖產生、纖維素酶活性等造成影響,這說明不同植物病原細菌的雙組分信號轉導系統所起的生理功能有所不同。此外,在雙組分信號轉導系統中,不同雙組分信號轉導系統之間存在著信號交流,可以使病原細菌中雙組分信號轉導系統中某個組分的缺失不一定會影響病原菌的表型。
歐美楊細菌性潰瘍病菌LqHK1基因與病原菌的致病性相關,為了解該病原菌的致病機制提供了新的研究方向。但是,目前只進行了組氨酸激酶基因研究,并未對其偶聯的反應調節蛋白進行功能研究,開展此項研究將有助于進一步了解雙組分結構與功能。此外,對于L.quercinaN-5-1中LqHK1基因是如何調節病原菌的信號的傳導、毒力以及參與到其他致病基因的調控等方面也需要進行深入研究。
4 結論
LqHK1基因缺失突變體在107楊樹上的致病能力顯著下降,Ream-time PCR檢測表明發病部位的病原菌含量減少; 突變體菌株的游動能力和生物膜的形成能力顯著降低,說明LqHK1基因在病原菌的致病過程中發揮著重要的作用。
郭利民,張鳴放,王洪軒. 2008. 濮陽市發現楊樹新病害. 河南林業科技,28 (3): 36-36.
應用在石油生產運輸中污染土壤的化學修復技術主要包括化學氧化技術、光催化降解技術、化學淋洗技術和化學改良技術。用化學方法修復污染土壤具有見效快、針對性強等優點,但是其最大的弊端在于易引起二次污染。但是其中光催化降解技術是一種新興的土壤處理技術,具有無二次污染的優點。并且在生物表面活性劑應用于化學淋洗技術后,也大大降低了化學淋洗技術的二次污染風險。
(Guo L M, Zhang M F, Wang H X. 2008. New poplar disease was discovered in Puyang of China. Journal of Henan Forestry Science and Technology, 28 (3): 36-36. [in Chinese])
李 賓,李愛寧,魏 強,等. 2015. 歐美楊細菌性潰瘍病菌hrcJ基因的功能分析. 林業科學,51 (12): 71-78.
(Li B, Li A N, Wei Q,etal. 2015. Functional analysis ofhrcJgene inLonsdaleaquercinasubsp.populi. Scientia Silvae Sinicae, 51 (12): 71-78. [in Chinese])
任飛娟. 2011. 歐美楊潰瘍病病原鑒定.北京: 北京林業大學碩士學位論文.
(Ren F J. 2011.Pathogen identifiction ofPopuluseuramericanacanker disease. Beijing: MS thesis of Beijing Forestry University.[in Chinese])
單世平,郭照輝,張德元,等. 2014. 貪銅菌6-5雙組分信號系統czcR2-czcS2的克隆和生物信息學分析. 中國農學通報,30 (36): 148-154.
(Shan S P, Guo Z H, Zhang D Y,etal. 2014. Gene cloning and bioinformatics analysis of two-cczcR2-czcS2 fromCupriavidus.Chinese Agricultural Science Bulletin,30(36):148-154. [in Chinese])
商 靖. 2014. 歐美楊細菌性潰瘍病菌侵染循環及病害流行研究. 北京: 北京林業大學博士學位論文.
(Shang J. 2014. Study on the infection cycle and field epidemic ofLonsdaleaquercinasubsp.populionPopulus×euramericana. Beijing: PhD thesis of Beijing Forestry University. [in Chinese])
宋 菲,向 軍,陸樹良. 2010. 激光共聚焦顯微鏡和改良微孔板法觀察細菌生物膜形成. 中國感染與化療雜志,10(5): 368-372.
(Song F, Xiang J, Lu S L. 2010. The formation of bacterial biofilms studied by using confocal laser scanning microscopy and modified microtiter-plate test. Chinese Journal of Infection and Chemotherapy, 10 (5):368-372. [in Chinese])
王梁燕. 2008. 耐輻射球菌中反應調節蛋白的研究. 杭州:浙江大學博士學位論文.
(Wang L Y. 2008. Study on response regulators inDeinococcusradiodurans. Hangzhou: PhD thesis of Zhejiang University. [in Chinese])
王 薇,胡丹丹,李槿年,等. 2014. 擬態弧菌OmpU蛋白的黏附功能及所介導的致病作用. 水產學報,38(5): 731-740.
(Wang W, Hu D D, Li J N,etal. 2014. Adhension function of OmpU protein fromVibriomimicusand its role in virulence. Journal of Fisheries of China, 38(5):731-740. [in Chinese])
楊 莉,張力群,賀 偉,等. 2014. Ⅲ型分泌系統是歐美楊潰瘍病菌Lonsdaleaquercina重要的致病因子. 植物病理學報, 44 (5): 512-520.
(Yang L, Zang L Q , He W,etal. 2014. Type III secretion system is an essential pathogenic factor in poplar canker pathogenLonsdaleaquercina. Acta Phytopathologica Sinica, 44 (5): 512-520. [in Chinese])
Bontemps G S, Madec E, Lacroix J M,etal. 2015. The two-component system CpxAR is essential for virulence in the phytopathogen bacteriaDickeyadadantiiEC3937. Environmental Microbiology, 17(11): 4415-4428.
Charles T C, Nester E W. 1993. A chromosomally encoded two-component sensory transduction system is required for virulence ofAgrobacteriumtumefaciens. Journal of Bacteriology, 175(20): 6614-6625.
Hoang T T, Karkhoff-Schweizer R A R, Kutchma A J,etal. 1998. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene, 212(1): 77-86.
Hoch J A. 2000. Two-component and phosphorelay signal,transduction. Current Opinion In Microbiol, 3(2): 165-170.
Kessler B, Lorenzo V, Timmis K N. 1992. A general system to integratelacZ fusions into the chromosomes of gram-negative eubacteria: regulation of thePm promoter of the TOL plasmid studied with all controlling elements in monocopy. Molecular and General Genetics MGG, 233(1): 293-301.
Khajanchi B K, Sha J, Kozlova E V,etal. 2009. N-acylhomoserine lactones involved in quorum sensing control the type VI secretion system, biofilm formation, protease production, andinvivovirulence in a clinical isolate ofAeromonashydrophila. Microbiology, 155(11): 3518-3531.
Kovach M E, Elzer P H, Hill D S,etal. 1995. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene, 166(1): 175-176.
Livak K J, Schmittgen T D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2- ΔΔCTmethod. Methods, 25(4): 402-408.
Li Y, He W, Ren F J,etal. 2014. A canker disease ofPopulus×euramericanain China caused byLonsdaleaquercinasubsppopuli. Plant Disease, 98(3): 368-378.
Mizuno T. 1997. Compilation of all genes encoding two-component phosphotransfer signal transducers in the genome ofEscherichiacoli. DNA Research, 4(2): 161-168.
Morohoshi T, Nakamura Y, Yamazaki G,etal. 2007. The plant pathogenPantoeaananatisproduces N-acylhomoserine lactone and causes center rot disease of onion by quorum sensing. Journal of Bacteriology, 189(22): 8333-8338.
Ninfa A J, Magasanik B. 1986. Covalent modification of the glnG product, NRI,by the glnL product, NRII, regulates the transcription of the glnALG operon inEscherichiacoli. Proceedings of the National Academy of Sciences, 83(16): 5909-5913.
Pan S Q, Charles T, Jin S,etal. 1993. Preformed dimeric state of the sensor protein VirA is involved in plantAgrobacteriumsignal transduction. Proceedings of The National Academy of Sciences, 90(21): 9939-9943.
Shang J, Liu B L, He W. 2015. A new method to detectLonsdaleaquercinain infected plant tissues by real-time PCR. Forest Pathology, 45(1): 28-35.
Yang L, Xu B, He W,etal. 2014. The HrpW protein ofLonsdaleaquercinaN-5-1 has pectate lyase activity and is required for full bacterial virulence. Journal of Basic Microbiology, 54(10): 1126-1135.
Yang W, Liu Y, Chen L,etal. 2007. Zinc up-ake regulator (zur) gene involved inzinc homeostasis and virulence ofXanthomonasoryzaepv.oryzaein rice. Curr Microbiol, 54: 307-314.
Zhao Y, Wang D, Nakka S,etal. 2009. Systems level analysis of two-component signal transduction systems in Erwinia amylovora: role in virulence, regulation of amylovoran biosynthesis and swarming motility. BMC Genomics, 10(1): 245.
(責任編輯 王艷娜 朱乾坤)
Functional Analysis ofLqHK1 inLonsdaleaquercinasubsp.populi
Wei Qiang Li Aining He Wei
(CollegeofForestry,BeijingForestryUniversityBeijing100083)
【Objective】 Poplar bacterial canker caused byLonsdaleaquercinasubsp.populiis a disease, which is serious harm to poplar industry. In this study, the biological function of theLqHK1 gene inL.quercinasubsp.populiwas investigated to provide viable knowledge for further understanding the pathogenic mechanism of pathogen.【Method】The bioinformatics method was used to indentify the two component systems LqHK1. The gene deletion mutant strain △LqHK1 was constructed by homologous recombination. Verifying PCR and Southern blot were used to investigate the biological characteristics of the mutant strains and their complementary strains,such as growth rate, motility, biofilm formation, and pathogenicity. At the same time, qRT-PCR was used to test the expression levels of the motility related genesflgB,flgC,flgE, and quantitatively analyze DNA content of poplar tissue pathogen in the vaccination site after pathogen inoculation.【Result】The two-component system geneLqHK1 was identified and the deletion mutant △LqHK1 had been obtained by homologous recombination. Phenotypic analysis showed that pathogenicity test on annual poplar branches was significantly less virulent than wide-type, while the complemented mutant HBLqHK1 restored the virulence to the wild-type level.The mutant had significantly less colonization in host than the wild type strain. Mutant strain had reduced swimming ability compared with the wild-type. The motility related genesflgB,flgC,flgEexpression significantly decreased in mutant; forming abilityLqHK1 mutant biofilms significantly decreased, but the growth rate ofLqHK1 mutant strains had no significant difference from wild type.【Conclusion】 Studies indicate that the bacterial poplar cankerLonsdaleaquercinasubsp.populitwo-component signal transduction systemLqHK1 gene is closely related to pathogenpathogenicity.
Lonsdaleaquercinasubsp.populi; two component system; pathogenicity
10.11707/j.1001-7488.20170412
2016-07-14;
2016-11-08。
國家自然科學基金項目(31400545); 中央高校基本科研業務費專項資金 (BLX2013015)。
S763.13
A
1001-7488(2017)04-0105-08
* 李愛寧為通訊作者。
