基于模型數據融合的中國溫帶和亞熱帶典型森林生態系統碳通量模擬
2017-05-25 00:37:31何洪林任小麗馮艾琳王輝民張軍輝
生態學報 2017年5期
葛 蓉,何洪林,任小麗,張 黎,馮艾琳,王輝民,張軍輝
1 中國科學院地理科學與資源研究所,生態系統網絡觀測與模擬重點實驗室,北京 100101 2 中國科學院大學,北京 100049 3 沈陽農業大學,沈陽 110866 4 中國科學院沈陽應用生態研究所,沈陽 110016
基于模型數據融合的中國溫帶和亞熱帶典型森林生態系統碳通量模擬
葛 蓉1,2,何洪林1,*,任小麗1,張 黎1,馮艾琳3,王輝民1,張軍輝4
1 中國科學院地理科學與資源研究所,生態系統網絡觀測與模擬重點實驗室,北京 100101 2 中國科學院大學,北京 100049 3 沈陽農業大學,沈陽 110866 4 中國科學院沈陽應用生態研究所,沈陽 110016
生態系統碳循環過程對水分響應的研究已成為全球變化關注的焦點問題之一。基于長白山溫帶針闊混交林與千煙洲亞熱帶人工針葉林觀測站2003—2009年生長季的碳通量(NEE)和氣象觀測數據,綜合考慮水分對光合、呼吸作用的影響,構建不同的NEE模型,并應用模型數據融合方法優化模型參數、遴選最適模型,系統分析了水分因子對不同森林生態系統碳循環的影響。結果表明:(1)優化后的模型參數均能被NEE實測數據較好約束。長白山生長季的光合、呼吸參數值均高于千煙洲,未考慮空氣飽和水汽壓差(VPD)的模型高估了千煙洲溫度敏感性參數(Q10)值、低估了千煙洲基礎呼吸速率參數(BR)值;(2)僅考慮VPD對光合作用影響的模型是長白山生長季碳通量模擬的最優模型,但模擬精度提高不顯著。不同模型間碳通量組分模擬結果差異較小;(3)考慮VPD和土壤含水量對光合、呼吸作用共同影響的模型是千煙洲生長季碳通量模擬的最優模型,并且顯著提高了模擬精度。未考慮水分的模型在生長季高估了總生態系統生產力(GEP)總量2.0%(21.85 g C/m2),同時更大幅度地高估了生態系統呼吸(RE)總量4.4%(38.02 g C/m2),從而導致NEE總量低估于實測值7.8%(18.55 g C/m2)。
水分;碳循環;模型數據融合;參數優化;模型選擇
模型是理解碳循環過程及其控制機制的重要手段[1],如何準確地模擬、預測陸地生態系統碳循環變化過程成為目前全球變化最為重要的研究問題之一[2]。通量觀測技術及全球觀測網絡的發展,為陸地生態系統碳循環模擬研究提供了大量連續觀測的數據支持[3- 5],光合、呼吸作用對光照、溫度、水分等環境要素變化的響應模型(碳循環過程環境響應模型)也隨之得到了長足的發展,成為跨尺度生物地球化學循環模型的重要組成部分,在陸地生態系統碳循環過程模擬中起著重要的作用[6- 7]。水分是影響生態系統碳循環過程的重要因素,尤其是那些水分受限的生態系統[8]。它通過影響微生物活性、植物根系生長、營養物質遷移及呼吸的溫度敏感性而限制生態系統呼吸作用[9],也通過調節植物氣孔導度和葉片水勢、影響光合系統的酶活性和CO2固定效率而限制生態系統光合作用[10]。目前在全球變化背景下,降水變率的增大導致干旱影響區域和強度的增大[11],生態系統碳循環過程對水分響應的研究越來越得到關注和重視[12-17]。其中Richarson等[13]、Liu等[14]構建呼吸作用對溫度、水分變化的響應模型,系統分析了水分對生態系統呼吸的影響;Lassop等[15]、Wagle等[16]基于多個通量站數據構建光合作用對光照、空氣濕度變化的響應模型,探討了水分在不同植被光合作用中的重要性。但水分變化同時影響生態系統的光合、呼吸作用,而現有碳循環過程環境響應模型較少考慮水分因子對光合、呼吸的共同影響,無法準確分析水分對生態系統碳循環過程的影響。
碳循環過程對水分響應形式的不同使模型結構存在差異性,同時模型參數也難以準確估算,導致模擬結果存在較大的不確定性。而近年來發展的基于貝葉斯理論的模型數據融合方法(Model-Data Fusion, MDF)[18]能夠充分利用觀測數據和參數先驗信息,優化模型參數、評估模型結構,為準確分析水分對生態系統碳循環影響提供了有效的技術手段[19]。目前MDF在碳通量模擬中已得到廣泛應用,但主要集中在通過馬爾科夫鏈-蒙特卡羅(MCMC,Markov Chain Monte Carlo)等優化方法降低模型參數不確定性方面[20-24]。綜合考慮模型參數及模型結構不確定性的研究較少[25],其中Sacks等[26]、Zobitz等[27]采用貝葉斯信息準則法(BIC)定量比較了不同呼吸、光合模型的優勢和不足,揭示了MDF應用在模型結構評價和選擇中的優越性。
千煙洲亞熱帶人工針葉林(QYZ)和長白山溫帶針闊混交林(CBS)分別代表了我國東部南北森林樣帶上的兩個典型森林生態系統類型[28],并在生長期水分條件差異顯著,這為研究水分對于不同生態系統碳循環過程的影響提供了天然的實驗條件。其中QYZ雨熱不同季造成了季節性干旱,對生態系統碳循環過程存在脅迫現象[29],而CBS在生長季節水分相對充沛,較少受到干旱的影響[30]。因此本文以QYZ和CBS為例,綜合考慮水分對光合、呼吸作用的共同影響,構建不同的碳循環過程環境響應模型,集成MCMC與BIC方法優化模型參數、評價模型結構,遴選兩個生態系統的最適模型,降低碳通量模擬結果的不確定性;并在此基礎上探討水分因子對QYZ、CBS光合、呼吸作用的影響。
1 數據與方法
1.1 站點概況和研究數據
千煙洲(QYZ)觀測站位于江西省紅壤丘陵農業綜合開發實驗站內,多為樹齡約30a的人工針葉林,屬于典型亞熱帶季風氣候;長白山(CBS)觀測站位于長白山自然保護區紅松針闊混交林內,多為樹齡約200a的成熟原始林,屬于典型的溫帶大陸性季風氣候。南北兩個生態系統水熱狀況差異明顯。中國生態系統研究網絡(CERN)和中國陸地生態系統通量研究網絡(ChinaFLUX)對兩個站點都進行了長期的生態監測和碳通量連續觀測,站點的基本特征見表1,其它詳細信息可以參考張雷明等[28]、劉允芬等[29]。

表1 研究站點的基本特征
研究數據包括模型驅動、反演及驗證數據。模型驅動數據為兩個站點2003—2009年的30 min 氣象觀測數據,主要包括氣溫、土壤溫度、土壤含水量、光合有效輻射、空氣飽和水汽壓差,均由常規自動氣象觀測系統測定。反演和驗證數據均為兩個站點經過質量控制后的30 min 渦度相關碳通量觀測數據,具體處理流程參見李春等[31]。其中,2003—2006年生長季(5—9月)的通量數據用于模型參數優化,2007—2009年生長季(5—9月)的通量數據用于模型驗證。
1.2 NEE模型組合
凈生態系統碳交換量(NEE)是總生態系統生產力(GEP)與生態系統呼吸(RE)兩大通量平衡的結果。本研究在溫度響應的呼吸模型與光響應的光合模型的基礎上,綜合考慮土壤水分和空氣水分因子對GEP、RE的影響,構建了4組NEE模型表述碳循環過程對水分響應的不同形式。表2描述了碳通量模型的組合方式。
Lloyd&Tayor模型[32](1)作為生態系統呼吸的溫度響應模型得到廣泛認可和應用,相比Van′t Hoff 方程和Arrhenius方程,它更好地描述了生態系統呼吸的變異性,在低溫和高溫條件下對溫度變化都有較強的響應能力[33]。Q10模型(2)采用溫度和土壤含水量作為模型驅動變量來描述溫度和水分對生態系統呼吸的協同作用[12],并假設生態系統呼吸的溫度敏感性因子Q10與土壤水分狀況密切相關。相比于其他考慮溫度、水分響應的呼吸模型(如連乘模型),Q10模型在干旱土壤條件下對溫度和水分有著更強的敏感性[33]。
(1)
(2)
式中,RE是生態系統呼吸(μmol m-2s-1);BR是參考溫度下的生態系統基礎呼吸(μmol m-2s-1);Q10是生態系統呼吸的溫度敏感性因子,代表溫度升高10℃生態系統呼吸的相對增長量;T是5cm土壤溫度(℃);Tref是參考溫度(℃),在此取10℃;SW是5cm土壤含水量(m3m-3);a,b是試驗參數,當b為正時,表示生態系統呼吸對溫度的敏感性隨水分的增加而增加。
生態系統光合作用強度與光合有效輻射呈顯著相關關系,通常符合直角雙曲線方程,可以用Michaelis-Menten模型(3)來表達[34]。當考慮GEP對光合有效輻射和飽和水汽壓差的協同響應時,可在Michaelis-Menten模型的基礎上將Amax替換成一個考慮VPD的指數遞減函數(4)[15],使Amax在VPD值較大時更符合生態學意義,即空氣越干燥,生態系統對二氧化碳吸收能力越弱。
(3)
(4)
式中,GEP是總生態系統生產力(μmol m-2s-1);LUE是初始光能利用效率(μmol CO2/μmol light);Amax為最大光合速率即光飽和時生態系統CO2同化能力 (μmol CO2m-2s-1); I為光合有效輻射即入射到植被上方的光量子通量密度(μmol light m-2s-1); VPD為冠層高度的空氣飽和水汽壓差(hPa),VPD0設定為10 hPa;Amax0是不受VPD約束下的最大光合速率;k是生態系統碳吸收對VPD響應的最大系數。
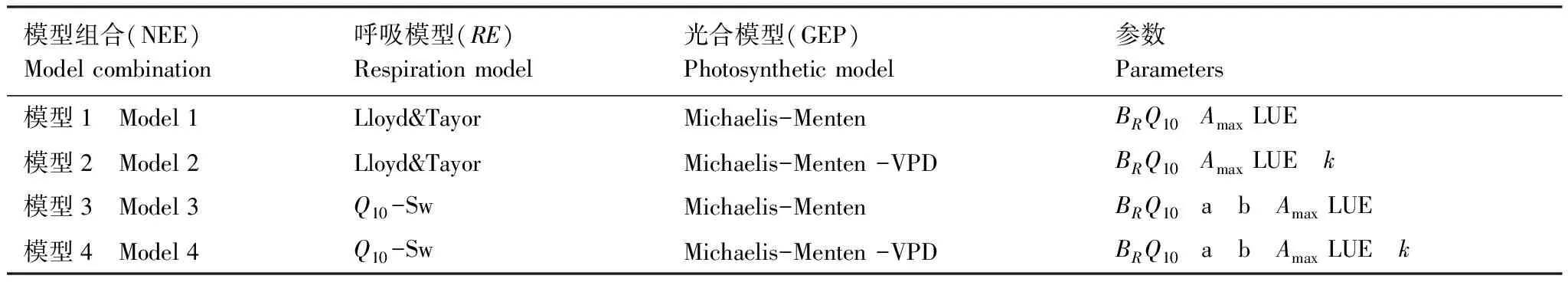
NEE=GEP+RE (5)
1.3 參數估計方法
本文采用的馬爾可夫鏈-蒙特卡羅方法是以貝葉斯定理為基礎,利用計算機進行模擬的蒙特卡羅方法。它通過構造馬爾可夫鏈來推斷參數的后驗信息,不斷迭代搜索到全局最優解。相比最小二乘法、極大似然法等傳統模型參數估計方法,MCMC方法能解決包含大量參數的復雜非線性問題,并結合了生態模型參數先驗知識和觀測樣本的信息,可以更加準確地估計參數。同時它還能提供參數后驗分布和置信區間,便于進行不確定性分析。Metropolis-Hastings算法[35-36]是最常用的MCMC抽樣方法,其主要步驟為:
1)在參數的先驗范圍內取參數初值θ0(可隨機也可指定),將迭代次數t設為0;
2)從建議分布中產生參數建議值Y;
3)產生符合(0,1)均勻分布的隨機數U;
4)計算接受概率α(θt,Y)
(6)
如果α(θt,Y)≥U,則接受參數建議值,即θt+1=Y;否則拒絕參數建議值,即θt+1=θt。
5)令t=t+1,重復2—5步,直到獲得足夠多的樣本。
根據貝葉斯理論,步驟4中參數集后驗概率密度的求解可以轉化為似然函數的求解,模型計算中采用形式簡單的對數似然函數,當參數迭代結束時便能獲取對數似然函數值的全局最優解。本文假設觀測數據隨機誤差服從正態分布,似然函數和其對數值分別如下:
(7)
(8)
式中,N為觀測數據個數,xi和ηi分別為第i個觀測值和模擬值,σ為觀測數據的標準差。
1.4 模型評價方法
傳統的模型評價方法包括比較模擬值與實測值的似然函數值、決定系數R2、均方根誤差(RMSE)以及平均絕對偏差(MAE),但其忽略了不同結構的模型引起的參數組成和數目的差異。本文采用的貝葉斯信息準則法不僅考慮了模型數據匹配情況,還考慮了參數的個數以及參與擬合的數據點個數,從而避免了模型結構的冗余以及數據的過度擬合,能夠從不同的模型中遴選出結構精簡、模擬效果最佳的模型[26-27]。
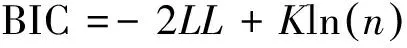
(9)
式中,K是優化參數的個數,n為優化過程中使用的數據點個數,BIC值越小說明模型越佳。
2 結果與討論
2.1 參數優化結果
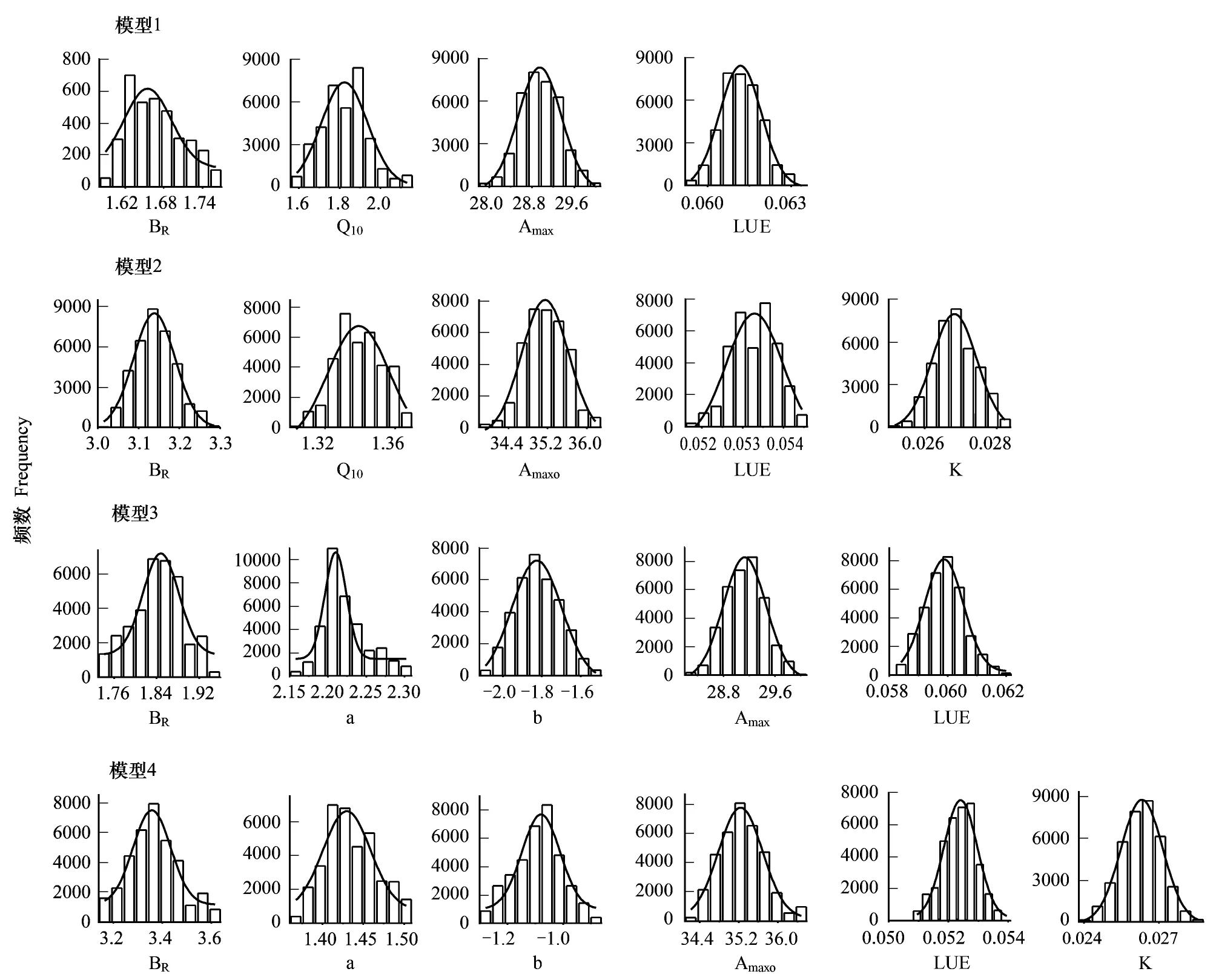
圖1 四組模型參數后驗分布圖(千煙洲森林生態系統)Fig.1 Posterior distribution of parameters in four models at QYZ
基于千煙洲、長白山兩個森林生態系統2003—2006年氣象和碳通量觀測數據,通過MCMC算法對4組模型進行參數優化,獲取參數后驗分布及最優參數集。圖1和圖2中4組模型參數的后驗分布均為正態分布,且各參數的標準差均較小(表3):呼吸參數標準差均控制在參數均值的10%以內,光合參數標準差均控制在參數均值的1%左右,表明優化后的參數均可以被NEE實測數據良好約束。

圖2 四組模型參數后驗分布(長白山森林生態系統)Fig.2 Posterior distribution of parameters in four models at CBS
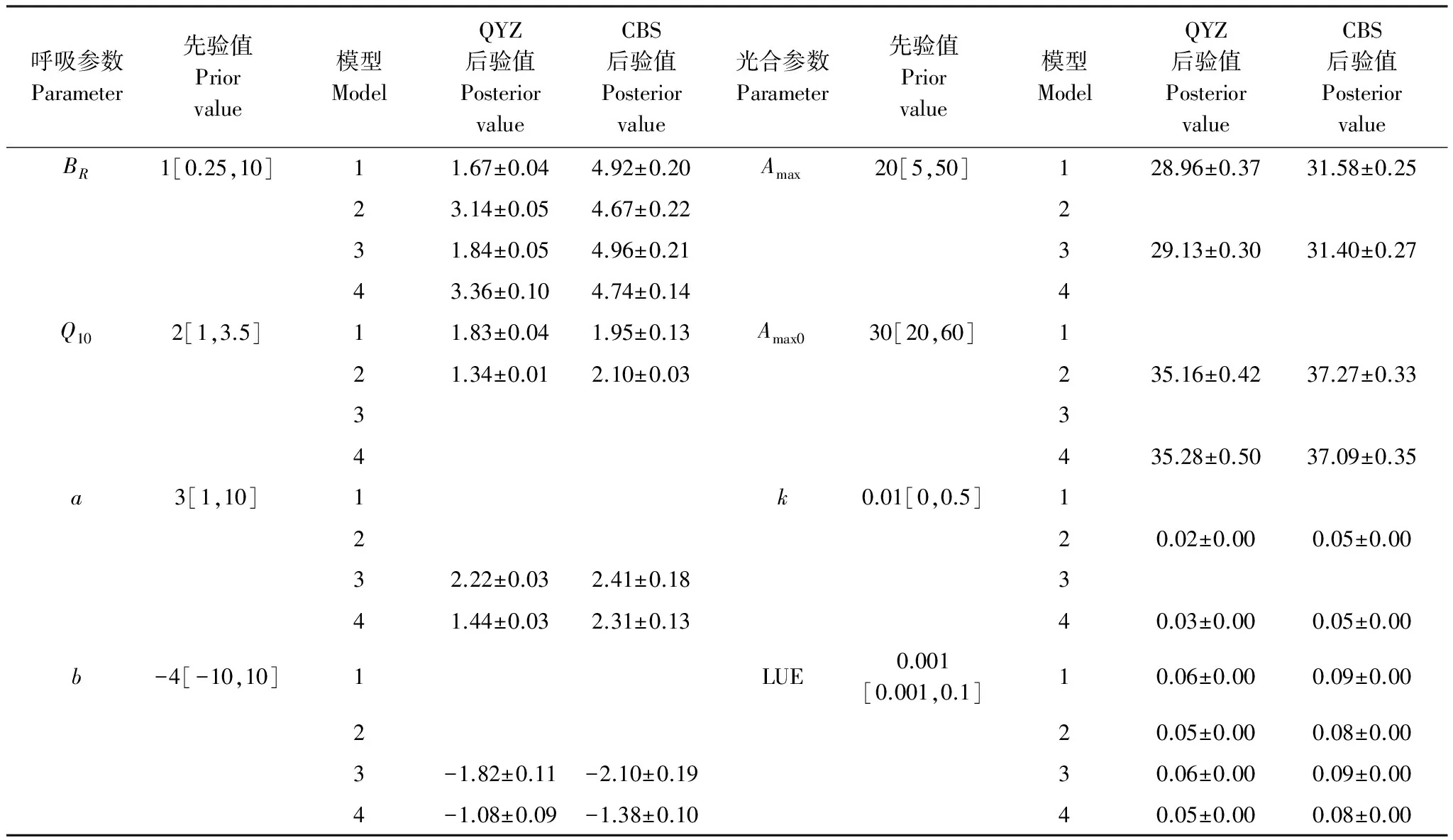
表3 參數優化結果
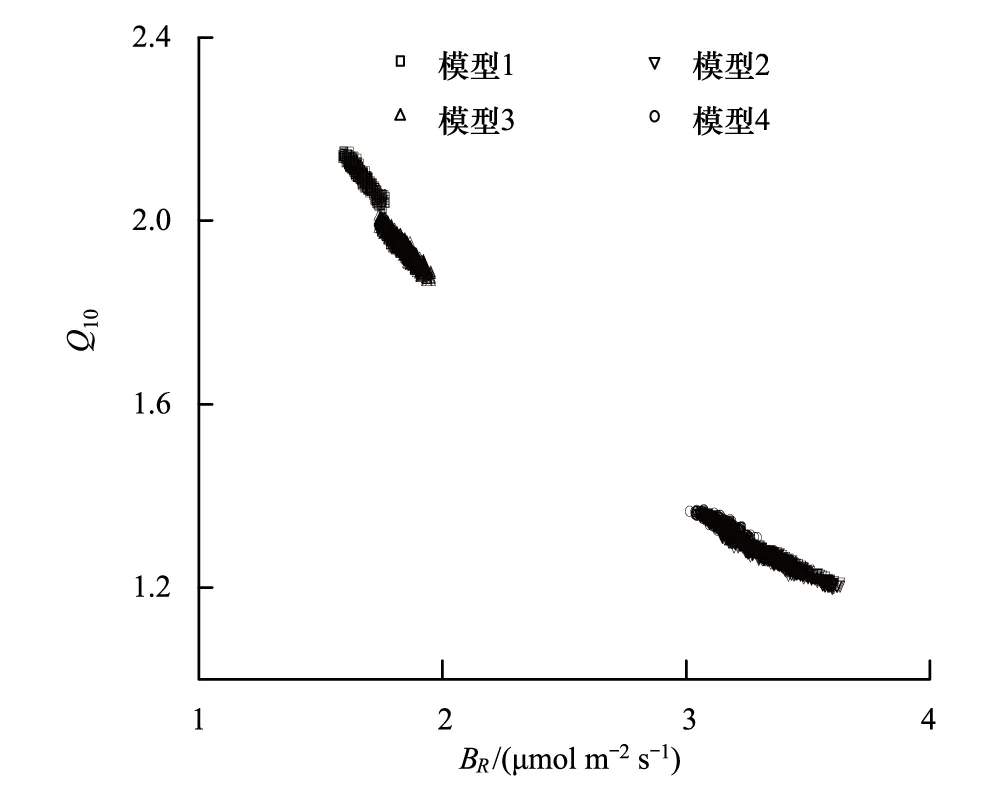
圖3 BR和Q10的最優參數集散點圖 Fig.3 Parameter space plots of basal respiration rate (BR) versus exponential temperature sensitivity (Q10)
對比生態系統光合參數(表3),不同模型間的參數變化微小,而不同生態系統的參數差異明顯:盡管QYZ作為人工林生態系統具有較強的光合能力,但CBS在生長季的光能利用效率LUE和最大光合速率Amax仍高于QYZ。其中,LUE平均約高0.03μmol CO2μmol-1light,Amax平均約高2.20μmol CO2m-2s-1。這是因為CBS溫帶落葉植物不同于QYZ的亞熱帶常綠針葉林,有明顯的生長季節和非生長季節之分,因此在有限生長季節內具有高效利用有效水熱資源的植被生長特性[28,37]。而QYZ在夏季受副熱帶高氣壓帶影響,易形成高溫和干旱天氣限制光合能力[29]。
對比生態系統呼吸參數(表3),不同模型間的的參數值變化較大,不同生態系統間的參數差異也明顯。模型間的參數變化主要表現在QYZ的BR和Q10,CBS的BR和Q10變化較小,反映出水分對QYZ呼吸作用的限制。QYZ生態系統的BR和Q10參數集散點(圖3)表明,四組模型BR和Q10之間均存在很強的負相關性(r=-0.89 — -0.97)。當模型中考慮水分因子后,Q10逐步降低。這是因為呼吸的溫度敏感性與水分呈正相關[38-39],QYZ夏季存在干旱脅迫故Q10值應處在較低的水平[40],表明考慮水分后的模型可修正Q10的高估。同時,圖3中考慮VPD因子的模型2和4散點分布相近,BR在3.02—3.63之間,Q10在1.20—1.37之間,符合參考前人文獻[14,20,24,28,33,41]中 QYZ呼吸參數值范圍,表明在QYZ考慮VPD對光合作用影響的模型能夠獲取合理的、具有物理特征意義的參數;而未考慮VPD因子的模型1和3散點分布相近:BR范圍在1.58—1.95之間,低估了45%;Q10范圍在1.77—2.15之間,高估了37%,該規律與Lasslop等[15]的研究結果一致。因本研究采用NEE數據聯合反演光合、呼吸參數,兩者存在聯動關系,故光合模型中VPD因子的修訂會影響呼吸參數的反演。呼吸參數的生態系統差異表現為:4個模型中CBS生長季的基礎呼吸速率BR、溫度敏感性參數Q10均顯著高于QYZ。其中,BR平均約高2.32μmol m-2s-1,Q10平均約高0.44,符合溫帶森林呼吸參數高于亞熱帶森林的規律。這主要是因為QYZ的緯度低于CBS、溫度高于CBS,而BR和Q10均隨著緯度降低、溫度升高而降低[33,38- 39]。同時,CBS紅松林林齡大于QYZ人工林,凋落物多、土壤有機質含量高[28,30,33],故呼吸底物相對較多,從而呼吸參數值較高。
2.2 最優模型選擇
基于千煙洲和長白山2007—2009年5—9月的氣象觀測數據,利用參數優化后的4組模型對兩個森林生態系統的碳通量分別進行模擬,通過BIC方法對考慮不同水分的模型模擬性能進行了定量對比(表4)。在CBS,僅考慮VPD對光合作用影響的模型2 BIC值最小,成為4組模型中CBS生長季碳通量模擬的最優模型。相比QYZ而言,水分因子對CBS模型性能提升較小,R2僅從0.77提高至0.83,BIC值降低了694:其中,考慮Sw因子使模型BIC平均僅下降了12,考慮VPD因子使模型BIC平均下降了688,表明水分對光合作用的影響更為明顯。這是因為CBS屬于季風影響的溫帶大陸性氣候,降水和溫度表現出一致的單峰季節變化,年均溫度較低,且土壤的持水能力較強,故Sw一直保持在較高的水平,對長白山生態系統的呼吸作用沒有明顯的限制作用[30,33,42]。而由于CBS在5月和9月的降水較少,氣溫卻保持在10℃以上,因此5和9月的VPD較高,6—8月的VPD較低,形成了明顯的雙峰季節變化[28],故VPD對長白山生長季的光合作用有一定限制作用。同時,Poulter等[43]研究也表明北方森林生態系統光合作用對降水變化更為敏感,因此考慮水分對光合作用影響的模型能夠更為準確地模擬溫帶森林生態系統對全球變化響應。盡管模型4的R2高于模型2,但模型4中Sw因子對模擬性能提升貢獻極小,反而增加參數個數使模型結構復雜,僅考慮VPD的模型2 BIC值最小,驗證了BIC方法挑選精簡模型的優越性。
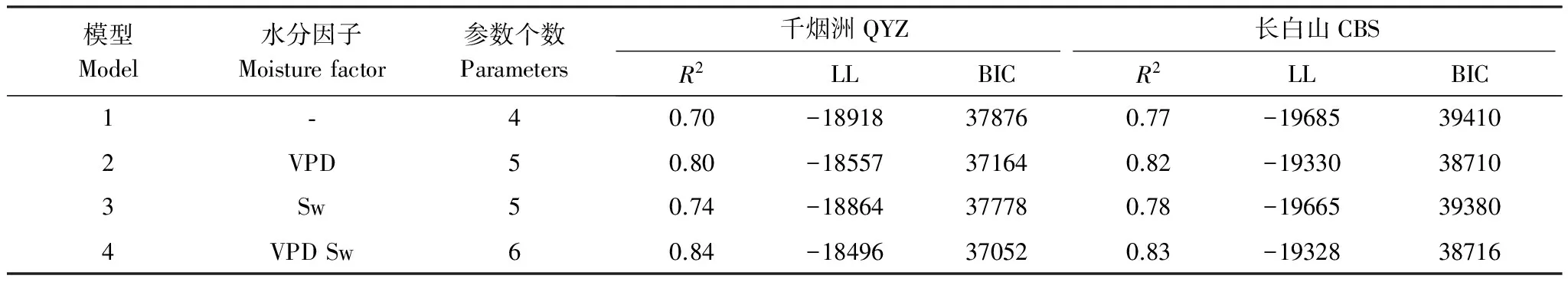
表4 BIC方法模型選擇結果
LL(Log Likelihood)為似然函數對數值,BIC(Bayesian Information Criterion)為貝葉斯信息準則
在QYZ,考慮VPD和Sw對光合、呼吸作用共同影響的模型4 BIC值最小,且模型4中參數相關性約束得更好(圖3),成為QYZ生長季碳通量模擬的最適模型。水分因子對QYZ模型模擬性能有顯著提升,R2從0.70到0.84提升了0.14(21%),同時BIC值降低了824:其中,考慮VPD因子使模型BIC平均下降了719,考慮Sw因子使模型BIC平均下降了106,表明水分是控制QYZ生長季碳通量交換的重要環境因子。這是因為QYZ屬于典型的亞熱帶季風氣候,在夏季受副熱帶高氣壓帶的控制和影響,水熱不同步性明顯,易形成伏旱[44-45]。VPD呈單峰特征,在生長季內可明顯上升至2kPa,同時表層Sw可顯著下降到0.1 m3/m3,故VPD和Sw對QYZ生長季的光合、呼吸作用均有明顯的限制作用[28,33,40,46-47]。
2.3 水分因子對碳通量模擬的影響
利用參數優化后的4組光合、呼吸模型對CBS、QYZ 2003—2009年生長季模型碳通量模擬值進行拆分,并取其多年平均值(表5)分析不同生態系統碳通量組分(NEE、RE、GEP)對水分的響應。
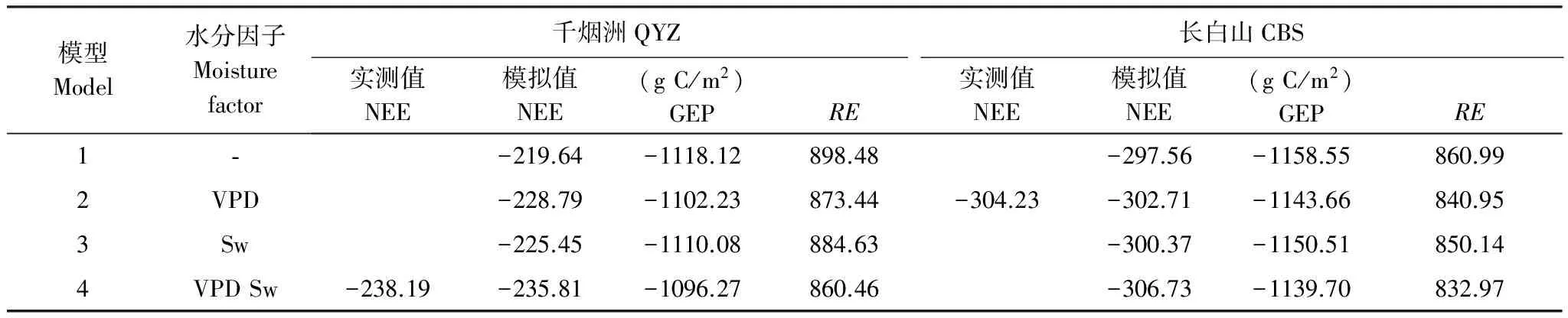
表5 不同模型模擬的碳通量組分生長季總量
在CBS,僅考慮VPD對光合作用影響的最優模型2各年NEE模擬值均與實測值最為接近,多年平均的NEE生長季總量模擬值僅低于實測值0.5%(1.52g C/m2)。與之相比,未考慮水分的模型1模擬效果最差,在生長季平均高估了GEP總量1.3%(14.89 g C/m2),同時高估了RE總量2.4%(20.04 g C/m2),但NEE總量僅低估于實測值2.2%(6.67 g C/m2),表明在CBS不同的水分響應模型之間模擬差異較小。
在QYZ,考慮水分對光合、呼吸作用共同影響的最優模型4各年NEE模擬值均與實測值最為接近,多年平均的NEE生長季總量模擬值僅低于實測值1.0%(2.38g C/m2)。與之相比,未考慮水分的模型1模擬效果最差,在生長季平均高估了GEP總量2.0%(21.85 g C/m2),同時更大幅度地高估了RE總量4.4%(38.02 g C/m2),從而導致整體上NEE總量低估于實測值7.8%(18.55 g C/m2)。模型2、3的NEE模擬值均不同程度地低估于實測值,其中碳通量組分對不同水分因子的響應不同:未考慮Sw因子的模型2主要高估了RE總量1.5%(12.98 g C/m2),同時小幅度高估了GEP總量0.5%(5.96 g C/m2)。表明Sw因子主要影響了QYZ生態系統呼吸,同時也通過調節冠層導度進而影響GEP的大小;未考慮VPD因子的模型3主要高估了GEP總量1.3%(13.81 g C/m2),同時高估了RE總量的2.8%(24.17 g C/m2),表明VPD因子對千煙洲碳通量組分模擬影響更大。這是因為干旱脅迫對千煙洲生態系光合作用的影響較大[41,44-45,48],而高水汽壓差是光合能力下降的主要原因[29]。同時由于千煙洲人工林林齡不長,尚處于較旺盛生長期,植物自養呼吸是千煙洲生態系統呼吸的主要來源[29,45],并且與氣溫相關性較強[33]。VPD作為空氣溫度和濕度的綜合指標直接影響了生態系統呼吸,也通過影響GEP而間接影響生態系統呼吸。
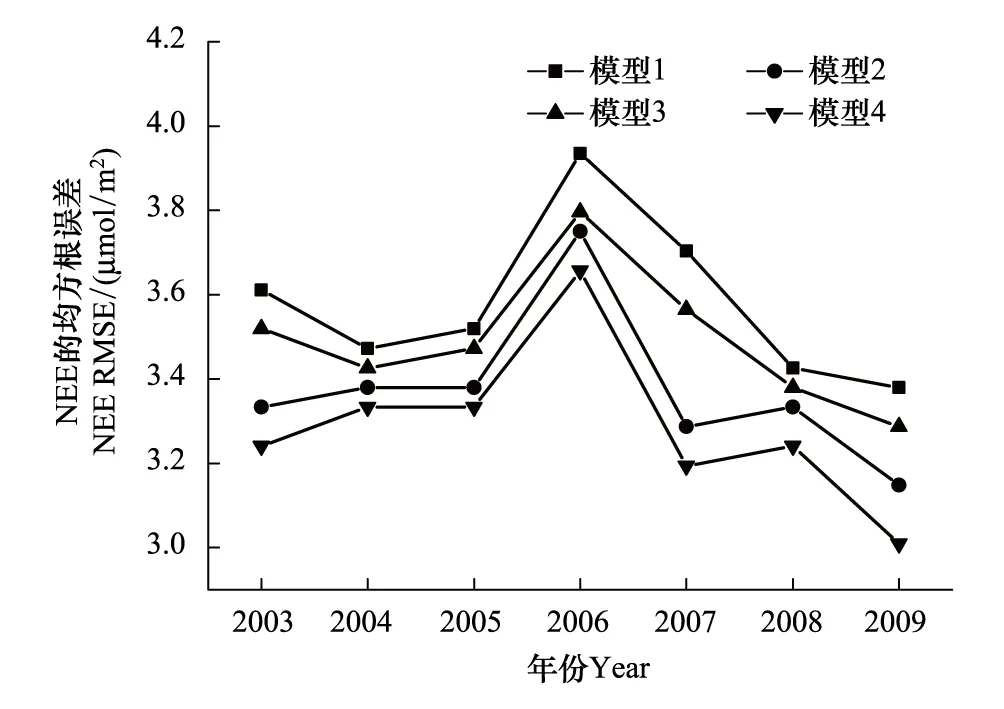
圖4 QYZ四組模型NEE半小時尺度模擬值與實測值的RMSE對比圖Fig.4 Comparison of RMSE of half-hourly modeled NEE from the four models and observed NEE at QYZ
QYZ生長季的水分條件年際差異較大(豐水年、干旱年),對比2003—2009各年QYZ生長季NEE模擬值與實測值的均方根誤差(圖4),發現2003、2007年考慮VPD的模型RMSE下降顯著,其余年份各模型間的模擬效果差異相對較小。而QYZ在這兩年的7月份分別發生了極度干旱和中度干旱[48],表明考慮水分因子(尤其是VPD)可顯著降低模型在QYZ干旱年份的模擬誤差。從QYZ干旱年(2003、2007)生長季模擬值與實測值的散點圖(圖5)中發現,未考慮VPD的模型1、3均有一個共同點,在NEE值較大和較小處散點的趨勢比較平緩,模型模擬存在“高值上不去,低值下不來”的現象,即極值的模擬效果要稍差于中值的模擬效果。尤其當NEE實測值達到-20μmol m-2s-1時,模型1、3的模擬值僅達到-13μmol m-2s-1左右,表明未考慮VPD的模型在干旱時期低估了NEE的峰值。這是因為干旱期高水汽壓差直接驅動氣孔關閉限制生態系統光合作用,導致午間生態系統碳吸收能力急速下降[15,49],因此NEE呈現出非對稱的日變化趨勢:上午時段值要高于下午時段,并在中午之前達到峰值。而未考慮VPD的模型模擬值則表現出隨光照變化的對稱趨勢,從而在上午時段低估NEE、下午時段高估NEE[16],并低估日變化中NEE的峰值,出現了圖5中極值模擬較差的現象。考慮VPD的模型則可以準確模擬出QYZ干旱期NEE日變化的非對稱趨勢,因此在散點圖極值處的平緩現象均有明顯改善,同時數據擬合線也更貼近于1∶1線。
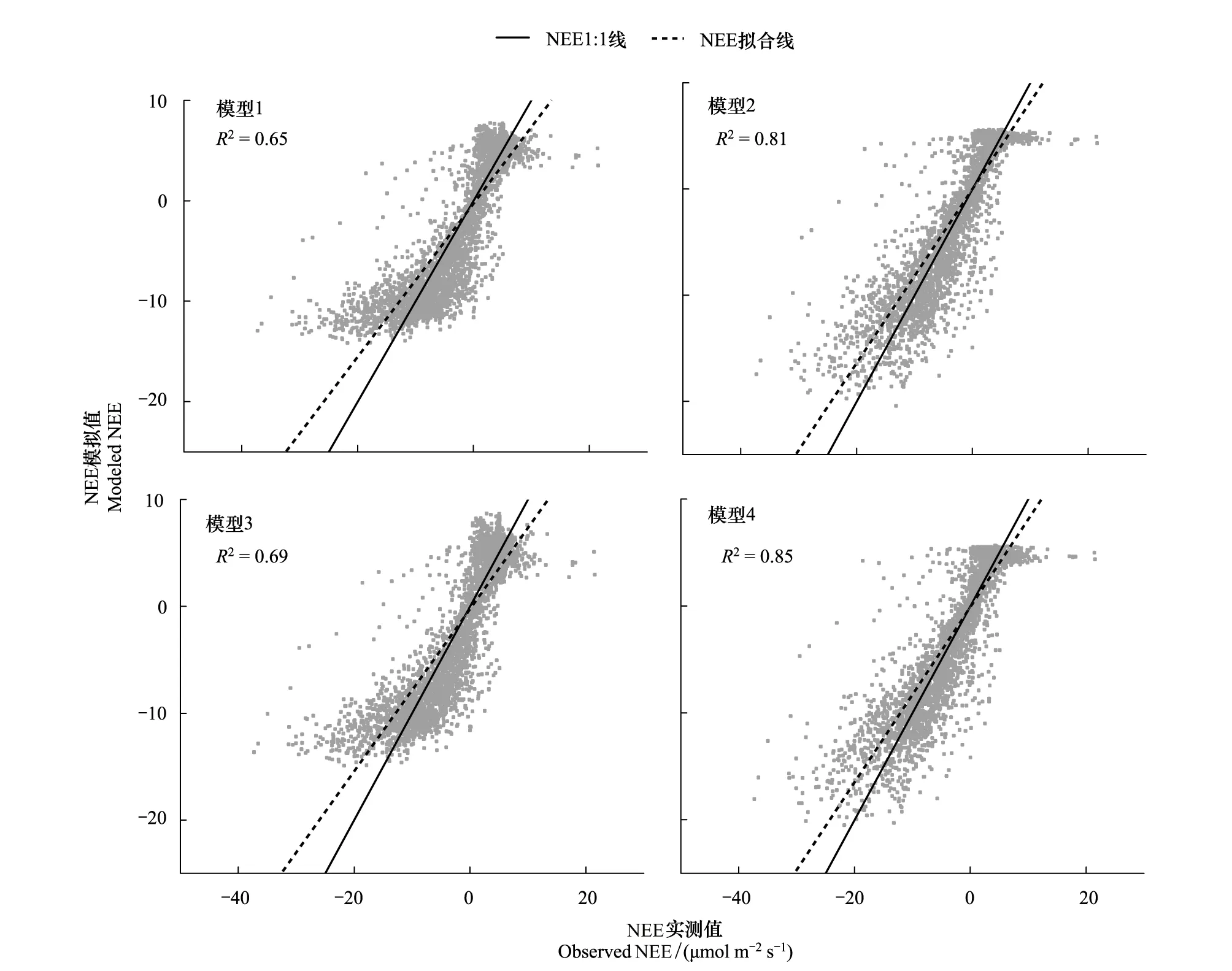
圖5 QYZ干旱年生長季半小時尺度NEE模擬值與觀測值散點圖Fig.5 Half-hourly NEE from observations and the four models at QYZ
3 結論
本文以千煙洲亞熱帶人工針葉林和長白山溫帶針闊混交林生態系統為例,應用模型數據融合方法為不同的碳循環過程響應模型優化參數,并遴選最適模型,系統分析了水分對不同生態系統碳通量模擬的影響。主要結論如下:
(1)優化后的光合、呼吸參數均能被NEE實測數據良好約束,參數后驗為正態分布、標準差較小。光合參數的生態系統差異明顯:CBS生長季的LUE和Amax均高于QYZ,但參數的模型間差異微小;呼吸參數的生態系統差異明顯:CBS生長季的BR和Q10均高于QYZ,同時參數的模型間差異也顯著:未考慮VPD的模型高估了QYZ的Q10值(37%)、低估了QYZ的BR值(45%)。光合、呼吸參數的生態系統差異準確反映了溫帶森林與亞熱帶森林在生長季的碳循環特征。
(2)同時考慮水分對光合、呼吸作用影響的模型和僅考慮水分對光合作用影響的模型分別是QYZ和CBS的最優模型。考慮水分因子對CBS模型的模擬性能并無明顯提升,對QYZ卻顯著提高,其中考慮VPD對光合作用的影響是QYZ模擬效果提升的主要原因。
(3)不同的水分響應模型在CBS碳通量組分模擬差異較小,而在QYZ差異明顯:未考慮水分的模型在生長季高估了GEP總量2.0%(21.85 g C/m2),同時更大幅度地高估了RE總量4.4%(38.02 g C/m2),從而導致整體上NEE總量低估于實測值7.8%(18.55 g C/m2)。其中,未考慮Sw因子主要引起RE高估,而未考慮VPD不僅引起GEP高估,也導致RE高估。同時,在QYZ干旱年VPD因子對碳循環的影響更為顯著,未考慮VPD的模型不能模擬出干旱期NEE日變化的非對稱趨勢并低估了NEE的峰值。
(4)本研究中提出的集成MCMC與BIC的模型數據融合方法可進一步應用于復雜的陸地生態系統過程模型,為改進生態系統模型結構、降低模擬的不確定性提供了有效途徑。
[1] Cao M K,Yu G R,Liu J Y,Li K R.Multi-scale observation and cross-scale mechanistic modeling on terrestrial ecosystem carbon cycle.Science in China Series D-Earth Sciences,2005,48(2): 17- 32.
[2] Heimann M, Reichstein M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature, 2008, 451(7176):289- 292.
[3] 趙士洞.國際長期生態研究網絡(ILTER)——背景、現狀和前景. 植物生態學報, 2001, 25(4): 510- 512.
[4] 于貴瑞, 孫曉敏. 中國陸地生態系統碳通量觀測技術及時空變化特征. 北京:科學出版社, 2008.
[5] Sulkava M, Luyssaert S, Zaehle S, Papale D. Assessing and improving the representativeness of monitoring networks: the European flux tower network example. Journal of Geophysical Research, 2011, 116(G3):278- 278.
[6] 于貴瑞, 孫曉敏.陸地生態系統碳通量觀測的原理與方法. 北京:高等教育出版社, 2006.
[7] 李克讓. 中國陸地生態系統過程及對全球變化響應與適應的模擬研究. 北京:氣象出版社, 2009.
[8] Bonan G B.Ecological Climatology: Concepts and Applications.2nd ed.New York: Cambridge University Press,2008.
[9] Luo, Y Q, Zhou X H. Soil respiration and the environment. Academic Press, an imprint of Elsevier, 2006.
[10] 于貴瑞,王秋鳳. 植物光合、蒸騰與水分利用的生理生態學. 北京:科學出版社, 2010.
[11] 田漢勤, 萬師強, 馬克平. 全球變化生態學:全球變化與陸地生態系統. 植物生態學報, 2007, 31(2):173- 174.
[12] Reichstein M, Tenhunen J D, Roupsard O, Ourcival J M, Rambal S, Dore S, Valentini R. Ecosystem respiration in two Mediterranean evergreen Holm Oak forests: drought effects and decomposition dynamics. Functional Ecology, 2002, 16(1): 27- 39.
[13] Richardson A D, Hollinger D Y. Statistical modeling of ecosystem respiration using eddy covariance: Maximum likelihood parameter estimation, and Monte Carlo simulation of model and parameter uncertainty, applied to three simple models. Agricultural and Forest Meteorology, 2005,131(s 3- 4):191- 208.
[14] Liu M, He H L, Yu G R, Sun X M, Zhang L, Han S J, Wang H M, Zhou G Y. Uncertainty analysis in data processing on the estimation of net carbon exchanges at different forest ecosystems in China. Journal of Forest Research, 2011, 17(3): 312- 322.
[15] Lasslop G, Reichstein M, Papale D, Richardson A D, Arneth A, Barr A, Stoy P, Wohlfahrt G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: critical issues and global evaluation. Global Change Biology, 2010, 16(1): 187-208.
[16] Wagle P, Kakani V G. Seasonal variability in net ecosystem carbon dioxide exchange over a young Switchgrass stand. Global Change Biology Bioenergy, 2013, 6(4): 339- 350.
[17] Nakaji T, Kosugi Y, Takanashi S, Niiyama K, Noguchi S, Tani M, Oguma H, Nik A R, Kassim A R. Estimation of light-use efficiency through a combinational use of the photochemical reflectance index and vapor pressure deficit in an evergreen tropical rainforest at Pasoh, Peninsular Malaysia. Remote Sensing of Environment, 2014, 150(7):82-92.
[18] Global Carbon Project.ScienceFrameworkandImplementation. Earth System Science Partnership (IGBP, IHDP, WCRP, DIVERSITAS) Report No.1. Global Carbon Project Report No.1, Canberra, 2003.
[19] Wang Y P, Trudinger C M, Enting I G. A review of applications of model-data fusion to studies of terrestrial carbon fluxes at different scales. Agricultural & Forest Meteorology, 2009, 149(11):1829- 1842.
[20] Liu M, He H L, Yu G R, Luo Y Q, Sun X M, Wang H M. Uncertainty analysis of CO2 flux components in subtropical evergreen coniferous plantation. Science in China Series D: Earth Sciences, 2009, 52(2): 257- 268.
[21] Richardson A D, Williams M, Hollinger D Y, Moore D J P, Dail D B, Davidson E A, Scott N A, Evans R S, Hughes H, Lee J T, Rodrigues C, Savage K. Estimating parameters of a forest ecosystem C model with measurements of stocks and fluxes as joint constraints. Oecologia,2010, 164(1): 25-40
[22] Ricciuto D M, King A W, Dragoni D, Post W M.. Parameter and prediction uncertainty in an optimized terrestrial carbon cycle model: Effects of constraining variables and data record length. Journal of Geophysical Research-Biogeosciences, 2011,116(G1):104- 121.
[23] Zhang L, Yu G R, Gu F X, He H L. Uncertainty analysis on modeled carbon fluxes in a broad-leaved Korean pine mixed forest by a process-based ecosystem model. Journal of Forest Research, 2012, 17(3):268- 282.
[24] Ren X L, He H L, Moore D J P, Zhang L, Liu M, Li F, Yu G R, Wang H M. Uncertainty analysis of modeled carbon and water fluxes in a subtropical coniferous plantation. Journal of Geophysical Research Biogeosciences, 2013, 118(4): 1674- 1688.
[25] Rayner P J. The current state of carbon-cycle data assimilation. Current Opinion in Environmental Sustainability, 2010, 2(4):289- 296.
[26] Sacks W J, Schimel D S, Monson R K, Braswell B H. Model-data synthesis of diurnal and seasonal CO2fluxes at Niwot Ridge, Colorado. Global Change Biology, 2006, 12(2): 240- 259.
[27] Zobitz J M, Desai A R, Moore D J P, Chadwick M A. A primer for data assimilation with ecological models using Markov Chain Monte Carlo (MCMC). Oecologia, 2011, 167(3): 599- 611.
[28] 張雷明. 中國東部南北森林樣帶典型生態系統碳收支特征及其生理生態學機制 [D]. 北京: 中國科學院研究生院, 2006.
[29] 劉允芬, 于貴瑞, 溫學發, 王迎紅, 宋霞, 李菊, 孫曉敏, 楊風亭, 陳永瑞, 劉琪璟. 千煙洲中亞熱帶人工林生態系統CO2通量的季節變異特征. 中國科學(D輯), 2006, 36(增刊I): 91- 102.
[30] 張軍輝, 于貴瑞, 韓士杰,等. 長白山闊葉紅松林CO2通量季節和年際變化特征及控制機制. 中國科學(D輯), 2006, 36(增刊1):60- 69.
[31] 李春, 何洪林, 劉敏, 蘇文, 伏玉玲, 張雷明, 溫學發, 于貴瑞.ChinaFLUX CO2通量數據處理系統與應用.地球信息科學, 2008, 10(5), 557- 565.
[32] Lloyd J, Taylor J A.On the temperature-dependence of soil respiration. Functional Ecology, 1994, 8(3):315- 323.
[33] 于貴瑞, 溫學發, 李慶康, 張雷明, 任傳友, 劉允芬, 關德新. 中國亞熱帶和溫帶典型森林生態系統呼吸的季節模式及環境響應特征. 中國科學D輯, 2004, 34(增刊2): 84- 94.
[34] Falge E, Baldocchi D, Olson R, Anthoni P, Aubinet M, Bernhofer C, Burba G, Ceulemans R, Clement R, Dolman H, Granier A, Gross P, Grunwald T, Hollinger D, Jensen N O, Katul G, Keronen P, Kowalski A, Lai C T, Law B E, Meyers T, Moncrieff H, Moors E, Munger J W, Pilegaard K, Rannik U, Rebmann C, Suyker A, Tenhunen J, Tu K, Verma S, Vesala T, Wilson K, Wofsy S. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agricultural and Forest Meteorology, 2001, 107(1): 43- 69.
[35] Metropolis N, Rosenbluth A W, Rosenbluth M N, Teller A H, Teller E. Equation of state calculation by fast computing machines. Journal of Chemical Physics, 1953, 21(6), 1087-1092.
[36] Hastings W K. Monte Carlo sampling methods using Markov chain and theirapplications. Biometrika, 1970, 57, 97- 109.
[37] Falge E, Tenhunen J, Baldocchi D, Aubinet M, Bakwin P. Phase and amplitude of ecosystem carbon release and uptake potentials as derived from FLUXNET measurements. Agricultural and Forest Meteorology, 2002, 113(1), 75- 95.
[38] Xu M, Qi Y. Spatial and seasonal variations of Q10 determined by soil respiration measurement at a Sierra Nevadan forest. Global Biogeochemical Cycles, 2001, 15(3), 687-696.
[39] Reichstein M, Rey A, Freibauer A, Tenhunen J, Valentini R, Banza J, Casals P, Cheng Y F, Grunzweig J M, Irvine J, Joffre R, Law B E, Loustau D, Miglietta F, Oechel W, Ourcival J M, Pereira J S, Peressotti A, Ponti F, Qi Y, Rambal S, Rayment M, Romanya J, Rossi F, Tedeschi V, Tirone G, Xu M, Yakir D. Modeling temporal and large‐scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Global Biogeochemical Cycles, 2003, 17(4):15- 1.
[40] Wen X F, Yu G R, Sun X M, Li Q K, Liu Y F, Zhang L M, Ren C Y, Fu Y L, Li Z Q. Soil moisture effect on the temperature dependence of ecosystem respiration in a subtropicalPinusplantation of southeastern China. Agricultural and Forest Meteorology, 2006, 137(3): 166- 175
[41] 任小麗, 何洪林, 劉敏, 張黎, 周磊, 于貴瑞, 王輝民. 基于模型數據融合的千煙洲亞熱帶人工林碳水通量模擬. 生態學報, 2012, 32(23), 7313- 7326.
[42] 趙曉松.渦動相關法估算森林生產力及與測樹學方法的比較. 碩士學位論文, 中國科學院研究生院, 北京, 2005.
[43] Poulter B, Pederson N, Liu H, et al. Recent trends in Inner Asian forest dynamics to temperature and precipitation indicate high sensitivity to climate change. Agricultural and Forest Meteorology, 2013, 178(4):31-45.
[44] 孫曉敏, 溫學發, 于貴瑞. 中亞熱帶季節性干旱對千煙洲人工林生態系統碳吸收的影響. 中國科學(D輯), 2006, 36(增刊2): 103- 110.
[45] 米娜.中亞熱帶人工針葉林生態系統碳水通量的觀測和模擬研究. 博士學位論文,南京信息工程大學,南京,2007年.
[46] 顧峰雪, 于貴瑞, 溫學發, 陶波, 李克讓, 劉允芬. 干旱對亞熱帶人工針葉林碳交換的影響. 植物生態學報, 2008, 32(5): 1041- 1051.
[47] 張舒, 申雙和, 溫學發, 張舒, 申雙和, 溫學發, 張心昱, 孫曉敏, 王輝民. 溫度和水分對中亞熱帶人工林生態系統呼吸的調控作用. 自然資源學報, 2012, 27(12):2057- 2070.
[48] Wen X F, Wang H M, Wang J L, Yu G R, Sun X M. Ecosystem carbon exchanges of a subtropical evergreen coniferous plantation subjected to seasonal drought, 2003- 2007. Biogeosciences, 2010, 7(1): 357- 369.
[49] K?rner C. Leaf diffusive conductances in the major vegetation types of the globe//Ecophysiology of photosynthesis. Berlin: Springer Heidelberg, 1995: 463- 490.
Carbon flux simulation of typical temperate and subtropical forest ecosystems in China based on model-data fusion approach
GE Rong1, 2, HE Honglin1,*, REN Xiaoli1, ZHANG Li1, FENG Ailin3, WANG Huimin1, ZHANG Junhui4
1KeyLaboratoryofEcosystemNetworkObservationandModeling,InstitutionofGeographicSciencesandNaturalresourcesresearch,ChineseAcademyofScience,Beijing100101,China2UniversityofChineseAcademyofSciences,Beijing100049,China3ShenyangAgriculturalUniversity,Shenyang110866,China4InstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China
Moisture effect on the carbon balance of terrestrial ecosystems is a key issue in global change research. It is crucial to accurately analyze the response of terrestrial ecosystem carbon cycle to moisture. However, the carbon flux models responding to environmental factors rarely consider the moisture effects on photosynthesis and respiration simultaneously; meanwhile there are still large uncertainties in model structures and parameters. Thus, this study was designed to (1) choose the optimal carbon flux model with accurate parameters for different ecosystems through model-data fusion approach, reducing the uncertainties of modeled results; (2) systematically analyze the influence of water factors on carbon flux simulation, including gross ecosystem productivity (GEP), ecosystem respiration (RE) and net ecosystem exchange (NEE). To consider the effects of moisture on both GEP andRE, we developed four different NEE models. Then, based on carbon flux and meteorological data during growing season from 2003 to 2009 in Changbaishan temperate mixed forest (CBS) and Qianyanzhou subtropical coniferous plantation (QYZ), Markov Chain Monte Carlo was employed to estimate model parameters, and Bayesian Information Criterion was applied to choose the optimal model for two forest ecosystems. The results showed that (1) the posterior values of model parameters were normally distributed, indicating that the parameters were well constrained by NEE. Photosynthetic and respiratory parameter values of CBS were higher than those of QYZ during the growing season. The model without vapor pressure deficit (VPD) overestimated the value of temperature sensitivity (Q10) and underestimated the value of basal respiration rate (BR) in QYZ; (2) the model considering VPD only was the optimal model for CBS,but its performance was not improved much. The modeled flux components were similar among the four models; (3) the model considering both VPD and soil water content (Sw) was the optimal model for QYZ, and its performance was improved significantly. The model ignored water factors overestimated 2% (21.85 g C/m2) of the total GEP, and 4.4% (38.02 g C/m2) of the totalRE, and therefore, underestimated 7.8% (18.55 g C/m2) of the total measured NEE during the growing season.
moisture effect; carbon cycle; model-data fusion; parameter optimization; model selection
中國科學院戰略性先導科技專項(XDA05050600);國家科技支撐計劃(2013BAC03B00);中國科學院科研信息化“科技領域云”項目(XXH12503-05-03)
2015- 10- 13;
日期:2016- 07- 13
10.5846/stxb201510132066
*通訊作者Corresponding author.E-mail: hehl@igsnrr.ac.cn
葛蓉,何洪林,任小麗,張黎,馮艾琳,王輝民,張軍輝.基于模型數據融合的中國溫帶和亞熱帶典型森林生態系統碳通量模擬.生態學報,2017,37(5):1409- 1420.
Ge R, He H L, Ren X L, Zhang L, Feng A L, Wang H M, Zhang J H.Carbon flux simulation of typical temperate and subtropical forest ecosystems in China based on model-data fusion approach.Acta Ecologica Sinica,2017,37(5):1409- 1420.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
