miR-23a和ESRP1在直腸癌中的作用*
2017-05-18 12:50:54勞玲娟宋新江
中國病理生理雜志 2017年5期
勞玲娟, 宋新江, 徐 佳
(1紹興第二醫院肛腸外科, 浙江 紹興 312000; 2岳陽市一人民醫院胃腸外科,湖南 岳陽 414000)
miR-23a和ESRP1在直腸癌中的作用*
勞玲娟1, 宋新江1, 徐 佳2
(1紹興第二醫院肛腸外科, 浙江 紹興 312000;2岳陽市一人民醫院胃腸外科,湖南 岳陽 414000)
目的: 研究miR-23a和上皮剪接調節蛋白1(epithelial splicing regulatory protein 1,ESRP1)在直腸癌組織及細胞系中的表達,以及對體外直腸癌細胞活力和凋亡的作用。方法: 采用RT-qPCR分析miR-23a在36例直腸癌組織和癌旁組織中的表達,免疫組化檢測ESRP1在直腸癌組織中的表達,分析miR-23a和ESRPl在直腸癌組織中的相關性;利用RT-qPCR檢測miR-23a在直腸癌Caco-2和SW480細胞及人正常結腸上皮細胞株NCM460中的表達;合成miR-23a inhibitor和inhibitor 陰性對照 (inhibitor NC),并將其分別轉染至SW480細胞后,通過CCK-8法檢測miR-23a inhibitor轉染SW480細胞后對細胞活力的影響,流式細胞術檢測轉染后細胞凋亡率,Transwell小室實驗檢測細胞侵襲;通過Western blot技術檢測SW480細胞中ESRPl蛋白的表達;構建野生型pGL3-ESRP1-3’UTR(wt-pGL3-ESRP1-3’UTR)或突變型pGL3-ESRP1-3’UTR(mut-pGL3-ESRP1-3’UTR)質粒,并分別與miR-23a inhibitor或inhibitor NC共轉染至HEK293和SW480細胞中,利用雙螢光素酶報告基因檢測試劑盒說明檢測雙螢光素酶活性;將ESRP1 mimic或mimic NC瞬時轉染SW480細胞后,CCK-8法和流式細胞術分別檢測細胞活力和凋亡;Western blot法檢測瞬轉ESRP1 mimic后對ESRP1、caspase-3、Smac和XIAP蛋白表達的影響。結果: miR-23a和ESRP1在直腸癌組織的表達較癌旁正常組織分別上調和下調,兩者呈明顯負相關(P<0.01);miR-23a的表達與直腸癌的淋巴結轉移和腫瘤浸潤深度相關;與NCM460細胞相比較,miR-23a在SW480細胞中的表達量顯著上調(P<0.01);轉染miR-23a inhibitor后,SW480細胞活力較inhibitor NC組顯著下降(P<0.01);轉染miR-23a inhibitor后SW480細胞早期凋亡率明顯升高,同時細胞體外侵襲能力受到抑制;螢光素酶報告基因結果表明ESRP1是miR-23a的直接靶基因;轉染miR-23a inhibitor至SW480細胞后ESRP1蛋白表達水平明顯升高;ESRP1 mimic轉染SW480細胞后可抑制細胞活力并誘導細胞凋亡,同時上調caspase-3和Smac的表達,下調XIAP的表達。結論: miR-23a可通過負向調控下游靶基因ESRP1從而影響直腸癌細胞生長和凋亡。
直腸癌; miR-23a; 上皮剪接調節蛋白1; 細胞凋亡
近年研究發現,微小RNA(microRNA,miRNA)作為一組高度保守的單鏈非編碼小RNA,能夠與其對應的靶標mRNA發生不完全結合,從而調控靶向調控基因的轉錄或翻譯[1-2]。近年來的研究證實,miRNA在腫瘤細胞中高表達或低表達能夠起到癌基因或抑癌基因的作用,并調控惡性腫瘤細胞的多種惡性生物學行為,從而導致腫瘤的發生和發展[3]。miRNA-23a (miR-23a)是位于染色體19p13.12上的miR-23a/24/27a家族成員。近年來報道稱miR-23a不僅調控正常細胞生長和分化,同時與多種惡性腫瘤細胞生長和轉移等惡性生物學行為關系密切[4-6]。但miR-23a在直腸癌中的表達和作用尚不明確,仍待進一步研究。為此,本次研究擬分析miR-23a和上皮剪接調節蛋白1(epithelial splicing regulatory protein 1,ESRP1)在直腸癌組織和細胞系中的表達,同時進一步研究兩者在直腸癌組織中的聯系及其對體外直腸癌細胞生長和凋亡的影響,探討miR-23a對直腸癌發生發展的生物學意義。
材 料 和 方 法
1 組織標本
收集2014年5月~ 2015年5月間在岳陽市一人民醫院胃腸外科經手術切除的36例新鮮直腸癌組織及癌旁正常組織標本,男性患者共有25例,女性患者共有11例,年齡在36.5歲~72.3歲之間,平均年齡為(61.4±4.3)歲。所有納入本次隊列的患者均經過兩位經驗豐富的病理科專家分別確診為直腸腺癌,且所有患者均未接受新輔助治療。其它排除標準包括下咽困難、急慢性感染、充血性心力衰竭、COPD和肝硬化。
2 試劑
Opti-MEM培養基、DMEM培養基和胎牛血清購自Gibco; Trizol試劑和Lipofectamine 2000購自Invitrogen;miR-23a inhibitor及相應的inhibitor陰性對照(negative control,NC)siRNA序列(inihibitor NC)購自GenePharma;miRNA提取試劑盒、逆轉錄試劑盒、RT-qPCR 試劑盒和細胞蛋白提取試劑盒購自TaKaRa;CCK-8試劑盒和Annexin V/PI凋亡檢測試劑盒購自南京凱基生物科技發展有限公司;抗ESRP1、caspase-3、Smac和X連鎖凋亡抑制蛋白(X-linked inhibitor of apoptosis protein,XIAP)和β-actin抗體購自Epitomics;野生型pGL3-ESRP1-3’UTR (wt-pGL3-ESRP1-3’UTR)或突變型pGL3-ESRP1-3’UTR (mut-pGL3-ESRP1-3’UTR)質粒、pRL-TK質粒及雙螢光素酶報告基因檢測試劑盒購自Promega;ESRP1 mimic及mimic陰性對照(mimic negative control siRNA,mimic NC)購自銳博生物。
3 細胞和實驗動物
人胚腎細胞株HEK-293、人直腸癌高分化腺癌細胞株Caco-2、人直腸癌低分化腺癌細胞株SW480及人正常結腸上皮細胞株NCM460均由上海細胞生物研究所提供。細胞培養于DMEM培養基中,培養基中補充10%胎牛血清、青霉素(1×105U/L)和鏈霉素(0.1 g/L)。細胞在37 ℃、5%CO2的細胞培養箱內進行培養。當細胞生長至占培養瓶底面積90%以上時用胰蛋白酶消化并用于實驗。
4 實驗方法
4.1 miRNA的提取及檢測 miRNA的提取實驗步驟嚴格按照miRNA提取試劑盒操作步驟進行。miR-23a的上游引物序列為5’-CCAGATCAGGCATCCTATTTG-3’,下游引物序列為5’-TATCATTGTCGAATTCCAGTGT-3’;內參照U6的上游引物序列為5’-GCTTCGCCAATTTGGGGTTG-3’,下游引物序列為5’-GTTGGTTCTGAGCGGTATCA-3’。依照PrimeScriptTMRT-PCR試劑盒說明書步驟將cDNA經反轉錄生成。對miRNA進行定量分析檢測步驟為:95 ℃ 2 min; 95 ℃ 45 s, 60 ℃ 60 s,30個循環。每個樣本檢測3次。通過2-ΔΔCt的方法計算miR-23a的相對表達差異倍數,實驗重復3次。
4.2 免疫組織化學法檢測ESRP1的表達 所有直腸癌和癌旁正常組織標本均使用4%的多聚甲醛固定、梯度濃度乙醇脫水、石蠟包埋及切片后,按照SP染色法進行免疫組化染色,使用PBS代替 I 抗作為陰性對照。切片的免疫組化: 烘片、脫蠟與水化、封閉、加 I 抗、加 II 抗、DAB 顯色、鏡檢。ESRP1染色陽性表達于細胞漿內,在低倍鏡下隨機選擇10個視野并拍照,后期使用Image-Pro Plus 6.0 圖像處理軟件分析ESRP1在標本中的陽性表達程度。
4.3 將miR-23a inhibitor、inhibitor NC、ESRP1 mimic及mimic NC轉染至SW480細胞 將對數生長期的SW480細胞接種于24孔板中,每孔2×105個細胞,將5 μL濃度為20 μmol/L的miR-23a inhibitor與5 μL轉染試劑Lipofectamine 2000混勻后加入細胞中,miR-23a inhibitor的轉染終濃度為100 nmol/L,將培養板置于細胞培養箱中孵育48 h后進行相關功能性、RT-qPCR和Western blot等檢測。Inhibitor NC、ESRP1 mimic及mimic NC的轉染方法同miR-23a inhibitor的轉染。
4.4 CCK-8法檢測細胞活力 將SW480細胞以每孔5×103個播種至96孔板中。在每個培養孔中添加完全培養基至100 μL,并將細胞放置于37 ℃、5% CO2細胞培養箱中至細胞融合,每孔添加CCK-8溶液0.01 mL反應1 h,用酶標儀測定490 nm波長處吸光度(A490),實驗重復5次,取平均值。
4.5 流式細胞術檢測SW480細胞凋亡 SW480細胞被冰PBS緩沖液洗滌3次后使用胰酶消化。加入2.5 μL Annexin V試劑和50 μL結合緩沖液在室溫下黑暗中反應15 min,隨后添加5 μL碘化丙啶和50 μL結合緩沖液在室溫下黑暗中孵育5 min。細胞懸浮在Annexin V試劑,細胞濃度為1×109~1×1010/L,所有樣品使用流式細胞儀進行分析,實驗重復3次,取平均值。
4.6 細胞體外侵襲實驗 收集對數生長期SW480細胞,加入無血清培養基中,調整細胞濃度為1×107/L,將300 μL細胞懸液和300 μL含20%胎牛血清的培養基分別加入Transwell上室和下室,放置細胞培養箱中培養24 h后加入結晶紫染色液染色20 min,在倒置顯微鏡下觀察穿過小室的藍染細胞,在高倍鏡下隨機計數10個視野,計算平均值,每組重復3次。
4.7 Western blot檢測細胞中蛋白的表達 SW480細胞在冰冷的PBS液中洗滌3次后重懸于細胞裂解液(每孔100 μL)中。將裂解的細胞置于冰上反應30 min,再在4 ℃、12 000×g離心20 min,收集上清后使用BCA蛋白檢測試劑盒對蛋白質濃度進行定量分析。蛋白質樣品(30 μg)使用10%十二烷基硫酸鈉聚丙烯酰胺凝膠電泳進行后轉移到0.45 μm的硝酸纖維素膜上,并使用5%脫脂奶粉于4℃下孵育過夜,該膜在4 ℃與 I 抗在室溫下反應30 min后,在室溫下與 II 抗再次孵育1 h。以內參照β-actin蛋白條帶灰度值來校正目的蛋白的灰度值。
4.8 螢光素酶報告基因檢測 將wt-pGL3-ESRP1-3’UTR或mut-pGL3-ESRP1-3’UTR質粒與pRL-TK質粒共轉染入HEK293和SW480細胞中,再分別將等量的miR-23a inhibitor或inhibitor NC再轉染至HEK293和SW480細胞中,48 h后收集細胞,根據雙螢光素酶報告基因檢測試劑盒制造商的要求對雙螢光素酶活性進行分析檢測。
5 統計學處理
采用SPSS 17.0統計軟件進行統計學分析。數據均以均數±標準差(mean±SD)表示,使用t檢驗比較2組數據間的差異,運用方差分析法對3組數據間的差異進行分析,運用2檢驗對臨床病理學特征數據進行分析,以P<0.05為差異有統計學意義。
結 果
1 miR-23a和ESRP1在直腸癌組織中的表達
RP-qPCR檢測結果顯示,miR-23a在88.9%(32/36)的直腸癌組織中的表達顯著上調,與癌旁正常組織相比較,有統計學顯著性(P<0.01),見圖1A。同時對miR-23a與36例直腸癌患者臨床病理學特征進行分析可知,在36例直腸癌組織標本中,miR-23a在浸潤漿膜層組織中的表達顯著高于未浸潤漿膜層組織,差異有統計學意義(P<0.05);淋巴結轉移組中miR-23a的表達顯著高于無淋巴結轉移組,差異有統計學意義(P<0.05);但直腸癌組織中miR-23a的表達水平與患者性別、年齡和分化程度無明顯相關性,見表1。利用免疫組化檢測ESRP1蛋白在36例直腸癌及癌旁正常組織標本中的表達可知,ESRP1在直腸癌組織中的表達較正常直腸組織明顯下降,差異有統計學顯著性(P<0.01),見圖1B、C;對miR-23和ESRP1在直腸癌組織中的表達進行相關性分析可知,miR-23和ESRP1在直腸癌組織中表達的Pearson相關系數為-0.794,呈明顯負相關(P<0.01),見圖1D。
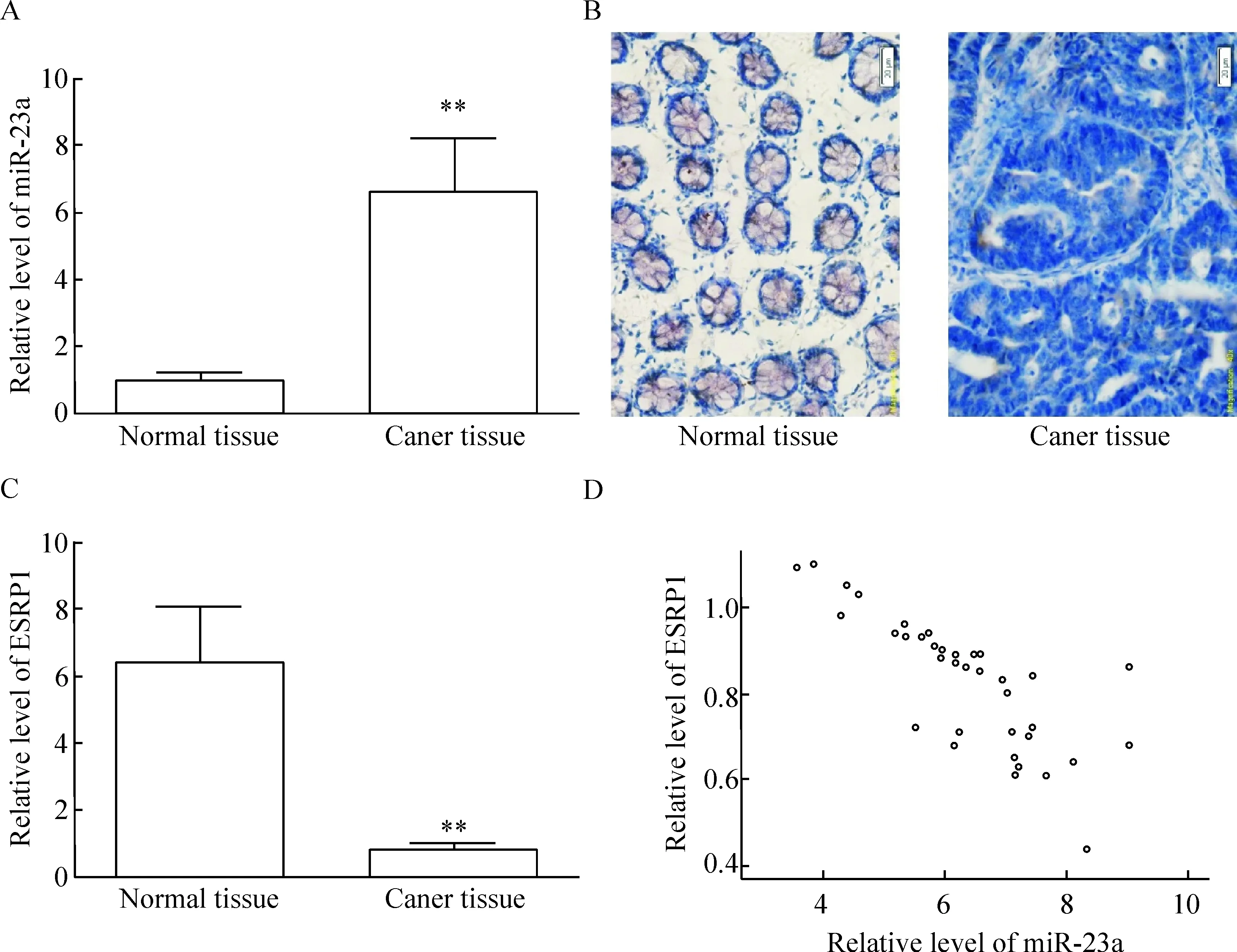
Figure 1.Expression of miR-23a and ESRP1 in human rectal cancer tissues.A: relative miR-23a expression was increased in human rectal cancer tissues compared with the matched adjacent noncancerous tissues; B: representative images showed that immunostaining with the antibody to recognize ESRP1 in the noncancerous and cancer tissues; C: the relative level of ESRP1 was down-regulated in human rectal cancer tissues compared with the matched adjacent noncancerous tissues; D: an inversely correlation between miR-23a and ESRP1 expression in human rectal cancer tissues was observed. Mean±SD.n=36.**P<0.01vsnormal tissue.
圖1 miR-23a 和ESRP1在人直腸癌組織中的表達
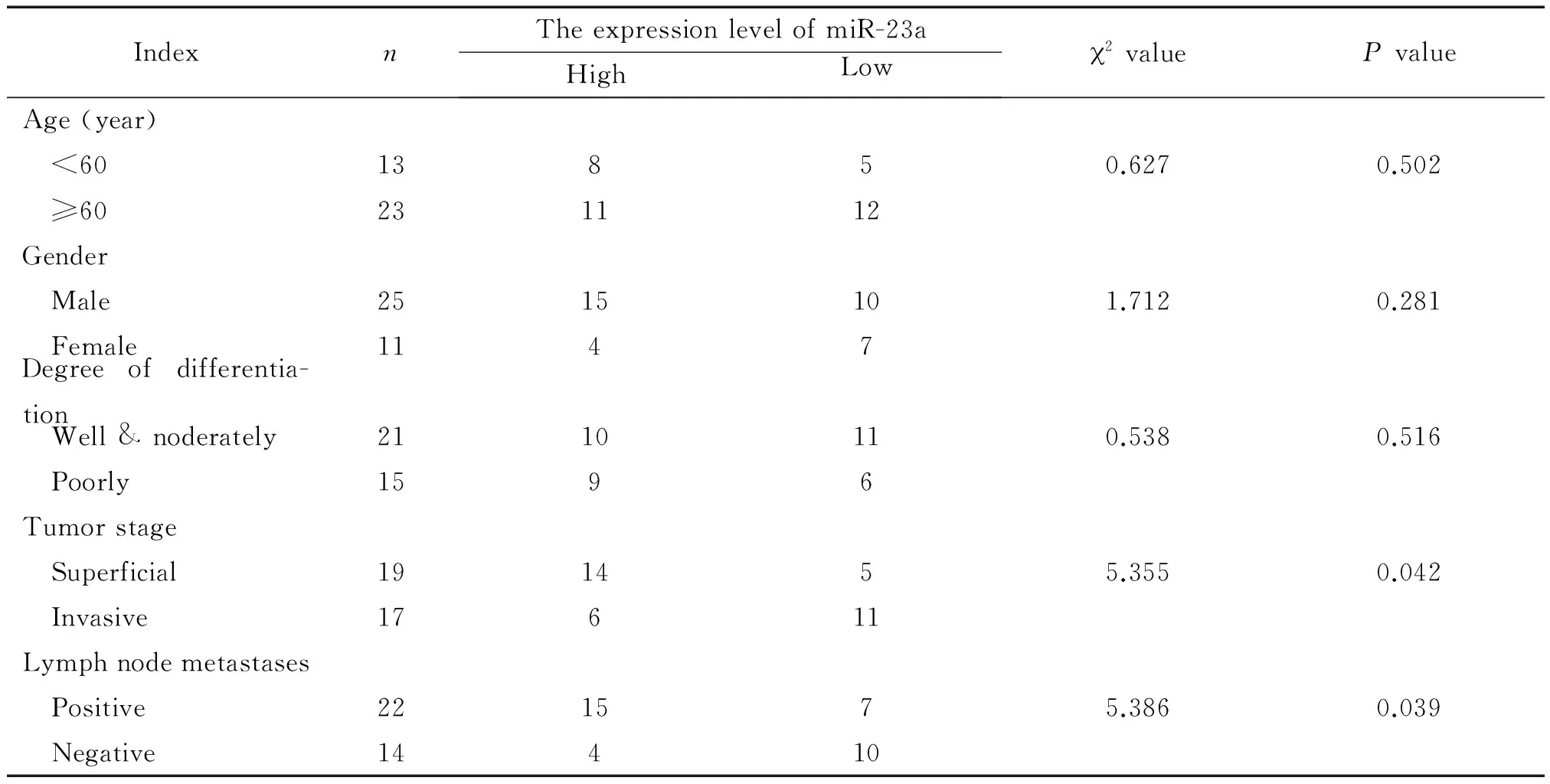
表1 miR-23a與36例直腸癌患者臨床病理因素的關系
2 miR-23a對體外直腸癌細胞活力、凋亡和侵襲的影響
RT-qPCR實驗檢測結果表明,miR-23a在人正常結腸上皮細胞系NCM460中的表達量較低,而在人直腸癌細胞系Caco-2 和SW480中表達明顯升高,差異有統計學意義(P<0.01);同時與Caco-2細胞相比,SW480細胞中miR-23a表達量明顯升高(P<0.01),見圖2A。因此,后續實驗選擇SW480細胞作為實驗對象。
將miR-23a inhibitor或inhibitor NC轉染進入SW480細胞48 h后,RT-qPCR結果表明miR-23a的相對表達量分別較對照組上升(15.25±2.21)%和下降(75.53±11.38)%,轉染miR-23a inhibitor可顯著下調miR-23a在SW480細胞中的表達,差異有統計學意義(P<0.01),而轉染inhibitor NC對miR-23a的相對表達量無明顯影響,見圖2B。CCK-8實驗結果表明,轉染inhibitor NC后48 h,細胞的吸光度與對照組相比較差異無統計學顯著性;而轉染miR-23a inhibitor后SW480細胞的吸光度較inhibitor NC組降低,差異有統計學顯著性(P<0.01),見圖2C。流式細胞術檢測細胞凋亡的結果表明,SW480細胞經轉染miR-23a inhibitor或inhibitor NC 48 h后細胞早期凋亡率分別為(14.5±2.1)%和(3.8±0.5)%,與對照組細胞[(2.4±0.4)%]相比較,差異有統計學意義(P<0.01),見圖2D。細胞侵襲實驗表明,SW480細胞經轉染miR-23a inhibitor后可顯著抑制細胞侵襲行為,SW480細胞經轉染miR-23a inhibitor或inhibitor NC 48 h后侵襲出Transwell小室的細胞數分別為14.6±2.5和85.8±11.3,與對照組細胞(94.6±12.7)相比較,差異有統計學顯著性(P<0.01),見圖2E。
3ESRP1是miR-23a的直接靶基因
通過miRNA預測軟件TargetScan進行分析可知,miR-23a與ESRP1的3’UTR存在結合位點,推測ESRP1可能是miR-23a的靶向調控基因(圖3A)。為了進一步驗證miR-23a與ESRP1 3’UTR是否能直接結合,我們構建wt-pGL3-ESRP1-3’UTR或mut-pGL3-ESRP1-3’UTR質粒,再分別與miR-23a inhibitor或inhibitor NC共轉染至HEK293細胞和SW480細胞中。結果表明野生型ESRP1 3’UTR質粒和miR-23a inhibitor共轉染后的螢光素酶活性較野生型ESRP1 3’UTR質粒和inhibitor NC共轉染后的螢光素酶活性明顯升高(P<0.01)。而突變型ESRP1 3’UTR質粒和miR-23a共轉染組與突變型ESRP1 3’UTR質粒和inhibitor NC共轉染組的螢光素酶活性相比較,差異無統計學顯著性,從而表明ESRP1是miR-23a的直接靶向基因(圖3B)。另外,Western blot實驗結果顯示,miR-23a inhibitor轉染至SW480細胞后可顯著上調ESRP1蛋白的表達,而轉染inhibitor NC至SW480細胞后對ESRP1蛋白的表達無顯著影響(圖3C)。
通過轉染ESRP1 mimic至SW480細胞中,我們觀察到ESRP1 mimic可明顯上調ESRP1蛋白在SW480細胞中的表達,見圖4;同時CCK-8實驗結果表明,mimic NC組細胞的吸光度與對照組相比較差異無統計學顯著性,而ESRP1 mimic組細胞的吸光度較mimic NC組低(56.21±8.23)%,差異有統計學意義(P<0.01),見圖3D;流式細胞術檢測結果表明,與mimic NC組相比較,轉染ESRP1 mimic可明顯增加SW480細胞的凋亡率,差異有統計學意義(P<0.01),見圖3E。
4 ESRP1負向調控XIAP在SW480細胞中的表達
如圖4所示,Western blot檢測結果表明,轉染mimic NC對SW480細胞中caspase-3、Smac和XIAP蛋白的表達無明顯影響,但轉染ESRP1 mimic可明顯上調caspase-3和Smac蛋白在SW480細胞中的表達,同時抑制XIAP蛋白的表達,與mimic NC組相比較,差異均有統計學顯著性(P<0.05)。
討 論
miRNAs在惡性腫瘤的起源和發展過程中扮演著重要角色。miRNAs的表達水平在不同組織來源的腫瘤組織或腫瘤細胞系中有較大區別。miRNAs在腫瘤中發揮癌基因還是抑癌基因的作用取決于其下游靶向 mRNA的表達。經過近幾十年來對miRNAs與惡性腫瘤關系的研究,當前應用miRNAs用于直腸癌的治療引起胃腸外科研究人員的關注。通過基因芯片定量對結直腸癌組織中數百種miRNAs表達譜進行分析,結果表明有24種miRNAs在結直腸癌組織中表達水平異常[7]。沉默miR-6734可延緩結直腸癌細胞增殖,降低其惡性侵襲性行為,并誘導細胞凋亡[8]。另外有報道稱miR-200家族可通過下調Notch通路從而在調控結直腸癌增殖和凋亡中發揮了重要作用[9]。近期研究發現miR-23a在結腸癌組織中表達較正常結腸組織中表達明顯上調[10],但其作用機制尚不明確。
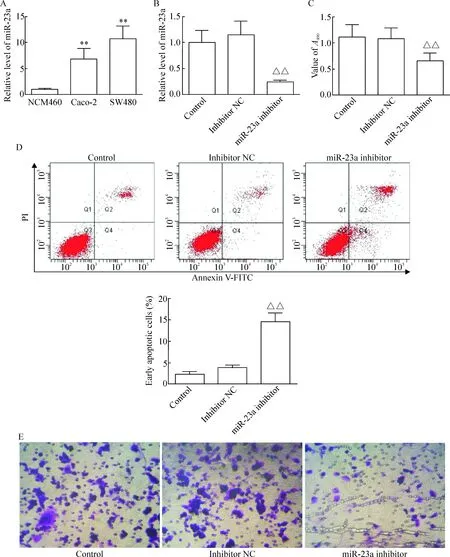
Figure 2.Effects of miR-23a on the viability, apoptosis and invasion of human rectal cancer cells. A: the expression of miR-23a was significantly higher in the malignant Caco-2 cells and SW480 cells than that in the normal human colonic epithelial cells by RT-qPCR; B: decreased miR-23a level in the SW480 cells was detected by RT-qPCR; C: the rectal cancer SW480 cells were transfected with miR-23a inhibitor or inhibitor NC for 48 h, and the cell viability was measured by CCK-8 assay; D: the rectal cancer cells were processed for Annexin V/PI staining and analyzed with flow cytometry after transfected with miR-23a inhibitor or inhibitor NC for 48 h; E: the cell invasion was evaluated by Matrigel counting assay. NC: negative control. Mean±SD.n=3.**P<0.01vsNCM460;△△P<0.01vsinhibitor NC.
圖2 miR-23a對直腸癌細胞活力、凋亡和侵襲的影響
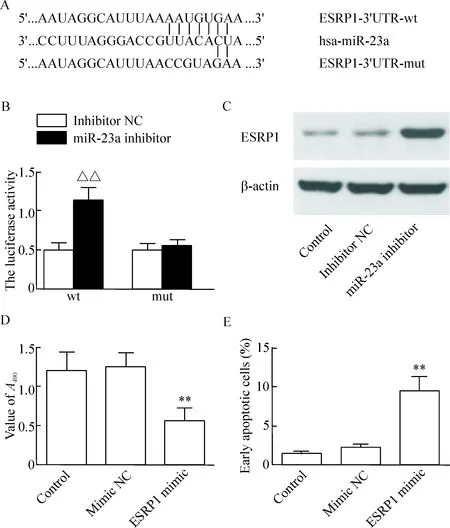
Figure 3.Verification of the relationship between ESRP1 and miR-23a. A: the miR-23a binding site in the 3’-UTR of ESRP1 mRNA; B: the relative luciferase activity was measured in the SW480 cells after co-transfection of ESRP1-3’UTR-wt or ESRP1-3’UTR-mut with miR-23a inhibitor or inhibitor NC; C: the protein level of ESRP1 in the SW480 cells transfected with miR-23a inhibitor or inhibitor NC was examined by Western blot; D, E: the SW480 cells were transfected with ESRP1 mimic to increase the expression of ESRP1, and the viability and apoptosis of the SW480 were analyzed by CCK-8 assay and flow cytometry, respectively. Mean±SD.n=3.△△P<0.01vsinhibitor NC;**P<0.01vsmimic NC.
圖3 ESRP1 與 miR-23a 關系驗證
本研究中,我們檢測了36例直腸癌組織和癌旁組織中miR-23a的表達情況,并分析其與直腸癌患者臨床病理特征的關系。 結果顯示,miR-23a在直腸癌組織中的表達水平明顯高于癌旁組織,提示miR-23a在直腸癌發生的早期階段可能充當著癌基因的作用;同時miR-23a在有淋巴結轉移直腸癌組織中的表達水平較無淋巴結轉移直腸癌組織明顯升高,提示miR-23a在直腸癌的轉移階段又起到了癌基因的作用,從而促進直腸癌的淋巴結轉移;此外,miR-23a的表達水平隨著患者TNM分期升高而逐漸升高,表明miR-23a可能與直腸癌的惡性進展一致;同時miR-23a在體外直腸癌細胞株中的表達也高于正常結腸上皮細胞。這種miR-23a在良、惡性組織及細胞中的表達差異表明miR-23a與直腸癌細胞的發生和發展可能存在一定的關聯,與文獻報道相仿[7]。
為進一步研究miR-23a在直腸癌惡性生物學行為中的作用,我們通過miR-23a inhibitor下調SW480細胞中miR-23a的表達,發現下調miR-23a不僅對體外SW480細胞增殖有明顯抑制作用,而且可促進細胞凋亡。另外體外侵襲實驗證實下調SW480細胞中miR-23a的表達可顯著抑制SW480細胞體外侵襲行為,該實驗結果與miR-23a和臨床病理學特征相關的結果相印證,從而進一步表明miR-23a與直腸癌惡性生物學行為密切相關。既往研究表明,miR-23a在調控宮頸癌、前列腺癌等多種惡性腫瘤增殖和凋亡中發揮了關鍵調控作用[4-6],但其精確的作用機制目前尚不十分明確。本研究通過生物信息學分析miR-23a為ESRP1潛在的上游調控因子,同時通過螢光素酶報告基因,首次在SW480細胞中驗證了ESRP1是miR-23a的直接靶向基因。ESRP1作為一種定位于細胞核內的蛋白,可通過直接結合特異性mRNA序列,調節細胞增殖、凋亡和侵襲等相關基因的表達,從而在調節惡性腫瘤的多種惡性生物學行為中發揮了重要作用[11]。Yae等[12]發現ESRP1在結腸上皮細胞中的異常表達可上調E-cadherin的表達,增強細胞的黏附性,從而抑制結腸上皮細胞的成瘤性。同時ESRP1在哺乳動物上皮細胞系發生上皮-間充質轉化過程中表達下調,而異位表達的ESRP1則可逆轉上皮細胞向間充質表型轉變[13]。本研究發現ESRP1在直腸癌組織中的表達較正常組織明顯減少,而ESRP1和miR-23a在直腸癌組織中的表達呈明顯負相關,提示ESRP1可能與直腸癌的發生和發展存在一定聯系。同時,下調miR-23a可增加其下游靶向基因ESRP1在直腸癌SW480細胞中的表達,有效抑制細胞活力并誘導細胞凋亡,但miR-23a是否通過靶向調控ESRP1而在調控直腸癌細胞活力和凋亡過程中發揮了一定作用尚未可知。為此,我們通過基因轉染技術上調SW480細胞中內源性ESRP1的表達,觀察到提升ESRP1在直腸癌細胞中的表達可有效抑制細胞增殖并誘導細胞凋亡,表明miR-23a可靶向調控直腸癌細胞中ESRP1的表達,從而對直腸癌細胞生長和凋亡產生一定的作用。
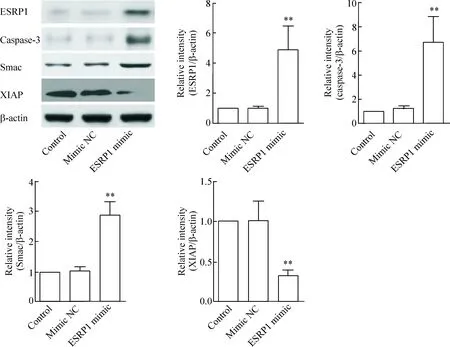
Figure 4.Western blot analysis of ESRP1, caspase-3, Smac and XIAP protein expression after transfection with ESRP1 mimic or mi-mic NC in the SW480 cellsinvitro. Mean±SD.n=3.**P<0.01vsmimic NC.
圖4 Western blot檢測轉染ESRP1 mimic或mimic NC對SW480細胞中ESRP1、caspase-3、Smac和XIAP蛋白表達的影響
細胞凋亡抑制蛋白(inhibitor of apoptosis protein,IAP)家族是caspase家族的內源性抑制因子,利用該家族所含有的高度保守的桿狀病毒IAP序列與caspase結合而發揮凋亡抑制作用,從而顯著抑制細胞凋亡[14]。XIAP作為IAP家族中抑制凋亡作用最強的成員,其可通過與caspase-3的結合而使其喪失活性,從而阻止細胞凋亡發生。而多種基因可負性調控XIAP抑制caspase-3活性作用過程,促凋亡蛋白Smac就是其中非常重要的一員,Smac可與凋亡抑制因子XIAP發生特異性結合,解除XIAP對caspase-3的抑制作用,從而引起活化的caspase的釋放,在調節細胞凋亡活動中發揮著重要的調控作用[15]。既往研究表明,XIAP是miR-23a的靶向調控基因之一[16],而ESRP1作為本次研究中發現的新的miR-23a靶向調控基因。ESRP1和XIAP均在調控腫瘤細胞生長和凋亡中發揮了一定作用,兩者之間是否存在一定聯系?在本研究中,上調ESRP1在體外直腸癌細胞中的表達可明顯抑制XIAP的表達,同時誘導caspase-3和Smac的表達,從而表明ESRPl可能通過調控凋亡抑制因子XIAP和促凋亡因子caspase-3和Smac的表達對體外直腸癌細胞的生長和凋亡產生一定的影響。
綜上所述,本研究表明miR-23a可負向調控ESRP1的表達從而影響體外直腸癌細胞增殖和凋亡,但miR-23a是否對體內直腸癌同樣具有調節增殖和凋亡能力尚未可知,同時雖然證實ESRPl與XIAP在體外直腸癌細胞中存在一定聯系,但ESRP1與XIAP之間是直接調控還是間接調控仍需進一步實驗明確。但本次研究為臨床直腸癌基因治療提供一個新的作用靶點,在揭示直腸癌的發病機制及治療上具有一定積極意義。
[1] 廖紫薇, 鄧紅霞, 張國平, 等. 致癌性微小RNA-106a對正常胃黏膜上皮細胞和胃癌細胞生長的影響[J]. 中國病理生理雜志, 2011, 27(10):1885-1889.
[2] 肖勝軍, 曾思恩, 方 宇, 等. LMP1對鼻咽癌細胞系CNE1癌基因微小RNA表達譜的影響[J]. 中國病理生理雜志, 2010, 26(4): 705-708.
[3] 蔣 振, 郭俊明, 肖丙秀. 特異性微小RNA抑制劑對胃癌細胞增殖的影響[J]. 中國病理生理雜志, 2009, 25(9):1726-1730.
[4] Yang Z, Wang XL, Bai R, et al. miR-23a promotes IKKα expression but suppresses ST7L expression to contribute to the malignancy of epithelial ovarian cancer cells[J]. Br J Cancer, 2016, 115(6):731-740.
[5] Aghaee-Bakhtiari SH, Arefian E, Naderi M, et al. MAPK and JAK/STAT pathways targeted by miR-23a and miR-23b in prostate cancer: computational andinvitroapproaches [J]. Tumour Biol, 2015, 36(6):4203-4212.
[6] Wen YC, Lee WJ, Tan P, et al. By inhibiting snail signaling and miR-23a-3p, osthole suppresses the EMT-mediated metastatic ability in prostate cancer [J]. Oncotarget, 2015, 6(25):21120-21136.
[7] Tanaka R, Tomosugi M, Sakai T, et al. MEK inhibitor suppresses expression of the miR-17-92 cluster with G1-phase arrest in HT-29 humancolon cancer cells and MIA PaCa-2 pancreatic cancer cells[J]. Anticancer Res, 2016, 36(9):4537-4543.
[8] Kang MR, Park KH, Yang JO, et al. miR-6734 up-regulates p21 gene expression and induces cell cycle arrest and apoptosis in colon cancer cells[J]. PLoS One, 2016, 11(8):e0160961.
[9] Suliman MA, Zhang Z, Na H, et al. Niclosamide inhibits colon cancer progression through downregulation of the Notch pathway and upregulation of the tumor suppressor miR-200 family[J]. Int J Mol Med, 2016, 38(3):776-784.
[10] 唐海林, 鄧 敏, 廖前進, 等. miR-23a與轉移抑制因子1在結腸癌中的表達及其臨床意義[J]. 中華病理學雜志, 2012, 41(1):28-32.
[11] Kikuchi M, Yamashita K, Waraya M, et al. Epigenetic regulation of ZEB1-RAB25/ESRP1 axis plays a critical role in phenylbutyrate treatment-resistant breast cancer [J]. Oncotarget, 2016, 7(2):1741-1753.
[12] Yae T, Tsuchihashi K, Ishimoto T, et al. Alternative splicing of CD44 mRNA by ESRP1 enhances lung colonization of metastatic cancer cell[J]. Nat Commun, 2012, 3:883.
[13] Ishii H, Saitoh M, Sakamoto K, et al. Epithelial splicing regulatory proteins 1 (ESRP1) and 2 (ESRP2) suppress cancer cell motility via different mechanisms[J]. J Biol Chem, 2014, 289(40):27386-27399.
[14] Zhou S, Huang Q, Zheng S, et al. miR-27a regulates the sensitivity of breast cancer cells to cisplatin treatment via BAK-SMAC/DIABLO-XIAP axis[J]. Tumour Biol, 2016, 37(5):6837-6845.
[15] Welsh K, Milutinovic S, Ardecky RJ, et al. Characterization of potent SMAC mimetics that sensitize cancer cells to TNF family-induced apoptosis[J]. PLoS One, 2016, 11(9):e0161952.
[16] Siegel C, Li J, Liu F, et al. miR-23a regulation of X-linked inhibitor of apoptosis (XIAP) contributes to sex differences in the response to cerebral ischemia[J]. Proc Natl Acad Sci U S A, 2011, 108(28):11662-11667.
(責任編輯: 林白霜, 羅 森)
miR-23a regulates cell growth and apoptosis of rectal cancer via targetingESRP1 gene
LAO Ling-juan1, SONG Xin-jiang1, XU Jia2
(1DepartmentofAnorectalSurgery,ShaoxingSecondHospital,Shaoxing312000,China;2DepartmentofGastrointestinalSurgery,TheFirstPeople’sHospitalofYueyang,Yueyang414000,China.E-mail:hosrmlan@163.com)
AIM: To investigate the expression of miR-23a and epithelial splicing regulatory protein 1(ESRP1) in rectal cancer tissues and cell lines as well as their effects on rectal cancer cell viability and apoptosis. METHODS: The relative levels of miR-23a in the rectal cancer tissues and cultured cells were assessed by RT-qPCR. The positive expression of ESRP1 in the rectal cancer tissues and non-cancer tissues was detected by immunohistochemical staining. The sequences of miR-23a inhibitor and inhibitor negative control (NC) were synthesized, and transfected into the SW480 cells. The cell viability was measured by CCK-8 assay. The apoptotic rate was analyzed by flow cytometry. The cell invasion was evaluated by Matrigel counting assay. The expression of ESRP1 was determined by Western blot. The wild-type pGL3-ESRP1-3’UTR (wt-pGL3-ESRP1-3’UTR) or mutant pGL3- ESRP1-3’UTR (mut-pGL3-ESRP1-3’UTR) plasmid and miR-23a inhibitor or inhibitor NC were co-transfected into the HEK293 and SW480 cells. The dual luciferase activity was detected according to Promega dual luciferase reporter gene assay kit instructions. The cell viability and apoptosis were evaluated by CCK-8 assay and flow cytometry analysis, respectively, after the SW480 cells were transfected with ESRP1 mimic or mimic NC. The expression of ESRP1, caspase-3, Smac and X-linked inhibitor of apoptosis protein (XIAP) in the SW480 cells was detected by Western blot.RESULTS: The expression of miR-23a was significantly up-re-gulated in the rectal cancer tissues and cell lines, while the positive expression of ESRP1 was significantly decreased in the rectal cancer specimens. The miR-23a expression was also closely related to lymphnode metastasis and TNM stages of rectal cancer patients. ESRP1 was inversely correlated with miR-23a in the rectal cancer tissues. After transfection with miR-23a inhibitor in human rectal cancer SW480 cells, the down-regulation of miR-23a induced significant inhibition of cell viability as compared with the cells transfected with inhibitor NC (P<0.01). Furthermore, the apoptotic rate induced by the miR-23a inhibitor transfection was markedly higher than that of control (P<0.01). Luciferase assay showed thatESRP1 was a direct target gene of miR-23a. The cell viability and apoptosis were inhibited and promoted, respectively, after transfection with ESRP1 mimic in the SW480 cells. Promoted expression of ESRP1 significantly up-regulated the levels of caspase-3 and Smac as well as down-regulated the expression of XIAP in the SW480 cells. CONCLUSION: The expression of miR-23a is significantly associated with the growth and apoptosis of human rectal cancer cells by targetingESRP1. miR-23a may be a potential therapeutic target for the treatment of rectal cancer in the future.
Rectal cancer; miR-23a; Epithelial splicing regulatory protein 1; Apoptosis
1000- 4718(2017)05- 0817- 09
2016- 11- 15
2017- 01- 23
岳陽市2014年第三批科技資助項目
R363; R735.3+7
A
10.3969/j.issn.1000- 4718.2017.05.009
雜志網址: http://www.cjpp.net
△通訊作者 Tel: 0730-8256328; E-mail: hosrmlan@163.com
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
