3個玉米自交系對鹽脅迫的生理響應及耐鹽性評價
2017-05-11 08:50:02王婧澤高樹仁孫麗芳胡凱鳳
干旱地區農業研究 2017年2期
關鍵詞:分析
王婧澤,高樹仁,孫麗芳,王 霞,王 晶,胡凱鳳,鄧 杰
(黑龍江八一農墾大學農學院, 黑龍江 大慶 163319)
3個玉米自交系對鹽脅迫的生理響應及耐鹽性評價
王婧澤,高樹仁,孫麗芳,王 霞,王 晶,胡凱鳳,鄧 杰
(黑龍江八一農墾大學農學院, 黑龍江 大慶 163319)
以自交系M-2導入系、Hr30導入系和Mo17導入系為試驗材料,研究不同濃度NaCl(0、100 mmol·L-1、150 mmol·L-1、200 mmol·L-1、250 mmol·L-1)對3個玉米幼苗的超氧化物歧化酶(SOD)、過氧化物酶(POD)、可溶性蛋白等生理指標的變化規律的影響,并采用主成份分析與隸屬函數相結合的方法對其耐鹽性進行綜合評價,旨在為玉米耐鹽性篩選及抗鹽新品種的選育奠定基礎。結果表明,3個自交系的株高、生物量、葉綠素含量、凈光合速率(Pn)以及可溶性蛋白質的含量大體上隨著鹽濃度的增加而呈下降的趨勢;根長、相對電導率、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性和根冠比大體上呈先上升后下降的趨勢。利用主成份分析歸納出株高、凈光合速率(Pn)、丙二醛(MDA)等9個與耐鹽密切相關的生理指標,運用模糊數學隸屬函數分析得出,3個玉米自交系的耐鹽性強弱依次為M-2導入系>Hr30導入系>Mo17導入系。
玉米;鹽;隸屬函數
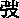
1 材料與方法
1.1 供試材料與設計
本課題組前期對野生堿蓬總DNA通過花粉管導入玉米自交系并穩定遺傳的20個后代進行耐鹽性篩選試驗,選擇其中3個耐鹽性較強的玉米自交系M-2導入系、Hr30導入系和Mo17導入系作為試驗材料。
1.2 試驗設計
試驗于植物生長室內進行,培養條件為:晝夜溫度為25℃/18℃,白天光照12 h,光強為1 000 μmol·m-2·s-1,相對濕度為60%~80%。挑選大小一致,無破損無病斑的玉米種子,用10%的次氯酸鈉表面消毒后,于24℃的培養箱內催芽,挑選長勢發芽一致的種子播于裝有等量蛭石的塑料盆內(直徑20 cm、高22 cm),每盆澆灌等量的1/2 Hoagland營養液。于三葉一心時分別以5個不同濃度梯度(0、100 mmol·L-1、150 mmol·L-1、200 mmol·L-1、250 mmol·L-1)的NaCl溶液進行鹽脅迫處理,處理7天后,選取相同功能的幼嫩葉片進行樣品采集。
1.3 測定項目及方法
1.3.1 生長指標的測定 鹽脅迫處理一周后,每個處理隨機選取5株幼苗,測定其株高、根長。將幼苗的地上部與地下部分離,置于 105℃下殺青30 min,80℃烘干至恒重,稱干重即生物量,并計算根冠比(根冠比=地上部干重/地下部干重)。
1.3.2 生理指標的測定 用便攜式Li-6400XTR光合儀(美國Li-COR公司)測定幼苗三葉一心時期生長點下第2片完全展開功能葉的凈光合速率(Pn),每葉片重復記錄3組數據,結果取其平均值。測定時葉室CO2濃度為400 μL·L-1,光強約為800 mmol·m-2·s-1,溫度為(25±2)℃。葉綠素含量的測定方法參照Wellburn[12]的方法測定。相對電導率的測定參照Lutts等[13]的方法測定。MDA含量的測定采用李合生等[14]的方法。SOD和POD活性的測定分別參照Giannopolitis C N等[15]和Rao M V等[16]的方法測定。可溶性蛋白含量的測定參照郝再彬等[17]的方法測定。
⑨Feng S.,Heerink N.,“Are farm households’land renting and migration decisions inter-related in rural China?”NJAS - Wageningen Journal of Life Sciences,4,2008.
1.4 數據分析
用Excel 2003進行數據整理,SPSS 19.0軟件進行單因素方差分析和主成分分析,采用Duncan檢驗法進行多重比較及差異顯著性分析,圖表中的數據均為3次重復的平均值。選取鹽脅迫一周后的主成分分析中得分系數較高的9個生理指標的測定值,采用模糊數學中的隸屬函數法進行耐鹽性綜合評價,計算方法如下。
隸屬函數值計算公式:
R(Xi)=(Xi-Xmin)/(Xmax-Xmin)
(1)
反隸屬函數值計算公式:
R(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)
(2)
2 結果與分析
2.1 鹽脅迫對玉米幼苗生長的影響
鹽脅迫對玉米幼苗的生長變化有直接的影響。如圖1(A)、1(B)所示,鹽脅迫對3個玉米自交系玉米幼苗的株高和生物量有明顯的抑制作用,當鹽濃度達到250mmol·L-1時,與對照相比,M-2導入系的株高和生物量分別下降14.6%和13.7%,Hr30導入系下降20.2%和33.2%,Mo17導入系下降21.78%和36.9%,M-2導入系下降幅度最小。如圖1(C)、1(D)所示,鹽脅迫對3個玉米自交系玉米幼苗的根長和根冠比有一定的促進作用,隨著鹽濃度的增加,3個玉米自交系的根長和根冠比呈先上升后下降的趨勢,Mo17導入系的根長和根冠比在鹽濃度為150mmol·L-1時達到最大值,分別比對照高67.8%和120.9%,隨著鹽濃度的繼續增加逐漸下降;M-2導入系和Hr30導入系在鹽濃度達到250mmol·L-1時達到最大值,與對照相比,M-2導入系的根長和根冠比增加了75.1%和248.5%,Hr30導入系的根長和根冠比增加了75.0%和183.2%,之后逐漸降低。
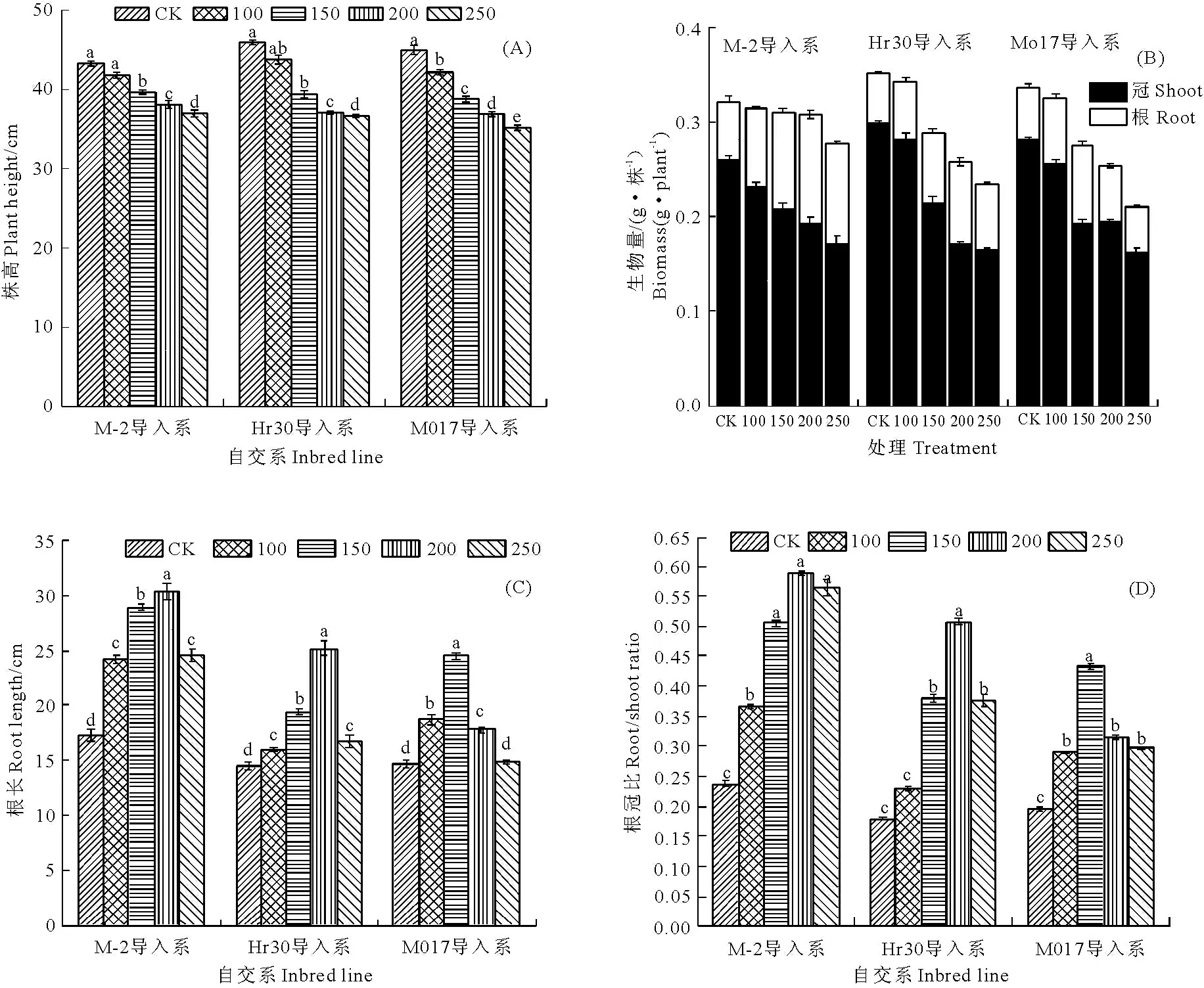
圖1 鹽脅迫下各導入系株高、生物量、根長和根冠比的變化
Fig.1Plantheight,totalbiomass,rootlengthandroot/shootratioofdifferentinbredlinesundersaltstress
2.2 鹽脅迫對玉米幼苗葉片葉綠素含量和凈光合速率的影響
鹽脅迫下,3個玉米幼苗葉片的葉綠素含量和凈光合速率顯著下降。如圖2(A)、2(B)所示,當鹽濃度增加到250mmol·L-1時,與對照相比,M-2導入系的葉綠素含量和凈光合速率分別比對照下降了46.7%和42.7%,Hr30導入系分別比對照下降了49.5%和64.8%,Mo17導入系分別比對照下降了51.2%和68.6%,M-2導入系的下降幅度較小。
2.3 鹽脅迫對玉米幼苗葉片電導率、MDA、可溶性蛋白及抗氧化酶活性的影響
結果如表1所示,3個玉米葉片的MDA含量和相對電導率的變化均隨著鹽濃度的增加呈先上升后降低的趨勢,當鹽濃度達到200mmol·L-1時3個自交系均達到最大值,與對照差異顯著,M-2導入系的MDA含量和相對電導率分別比對照增加115.3%和81.7%,Hr30導入系分別比對照增加了144.9%和90.4%,Mo17導入系分別比對照增加了156%和120.5%,M-2導入系的增幅較小。
3個玉米葉片的可溶性蛋白含量的變化大體上隨鹽濃度的增加而呈逐漸下降的趨勢。當鹽濃度達到100mmol·L-1時,Hr30導入系葉片可溶性蛋白的含量與對照相比略有升高,比同濃度下M-2導入系和Mo17導入系分別高7.04%和52.3%,隨著鹽濃度的增加,可溶性蛋白的含量顯著下降。當鹽濃度升高到250mmol·L-1時,M-2導入系的可溶性蛋白含量與對照相比下降了40%,但分別比Hr30導入系和Mo17導入系高35.8%和81.6%。
圖2 鹽脅迫下各導入系葉綠素含量和凈光合速率的變化
Fig.2Chlorophyllcontent(a+b)andnetphotosyntheticrateindifferentinbredlinesundersaltstress

表1 鹽脅迫對玉米幼苗葉片電導率、MDA、可溶性蛋白及抗氧化酶活性的影響
注:同列不同字母表示達到顯著性差異(P<0.05)。
Note: Different small letters in same columns indicate significant difference(P<0.05).
POD、SOD活性均隨著鹽濃度的增加呈先上升后下降的趨勢,當鹽濃度升高到150 mmol·L-1時,Mo17導入系的POD和SOD活性達到最大值,分別比對照增加了207.9%和183.2%,隨著鹽濃度的繼續增加,活性下降顯著。當鹽濃度升高到200 mmol·L-1時,M-2導入系和Hr30導入系的POD和SOD活性達到最大值,POD的活性分別比對照增加了173%和228.8%,分別比同濃度下的Mo17高54.5%和47.1%;SOD的活性分別比對照增加了103%和199%,分別比同濃度下的Mo17高54.5%和39.4%。
2.4 耐鹽指標的相關性分析和方差分析
對鹽脅迫下3個玉米自交系的11個生理指標進行了相關性分析(表2),大部分指標間存在著相關性,部分指標間的相關性達到了顯著或極顯著水平。為避免指標間的信息重疊,采用主成分分析,轉換成較少的綜合指標,能夠較好地反映玉米的耐鹽性。
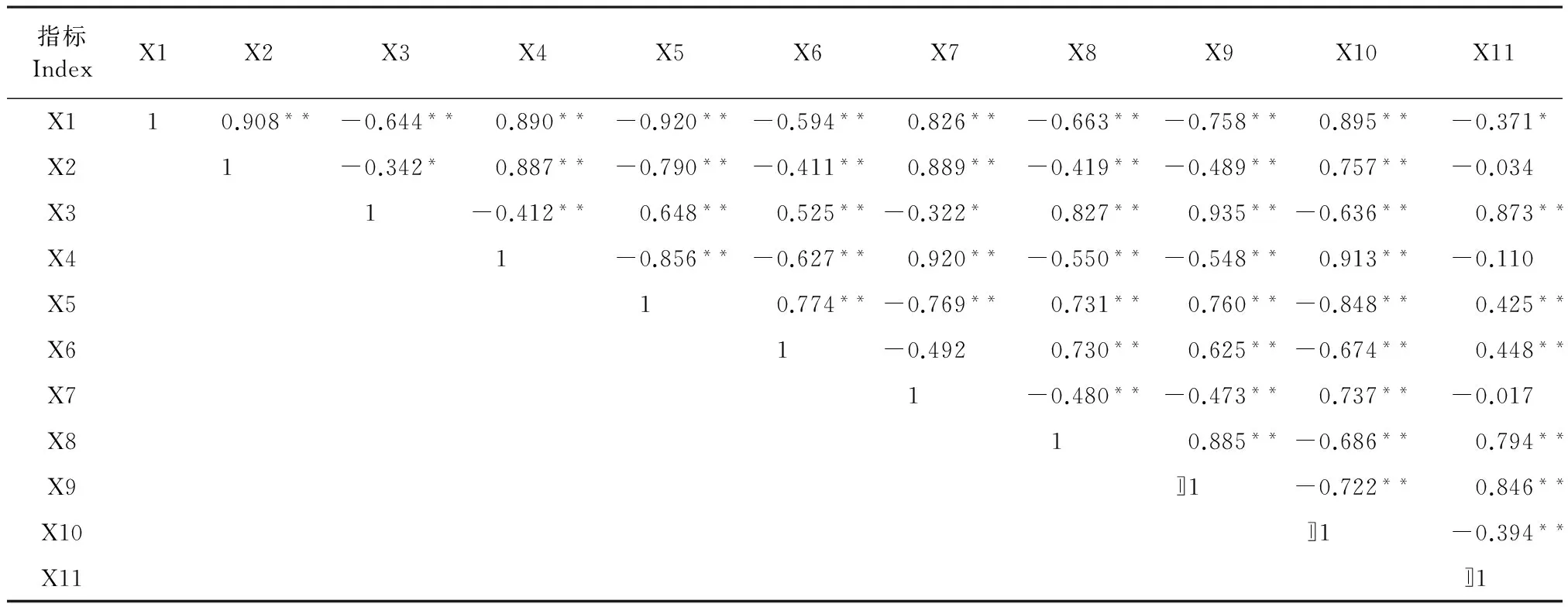
表2 玉米不同耐鹽性指標的相關關系
注:*和**分別表示在0.05和0.01的顯著水平。X1:株高;X2:生物量;X3:根冠比;X4:凈光合速率;X5:丙二醛;X6:相對電導率X7:可溶性蛋白;X8:過氧化物酶;X9:超氧化物歧化酶;X10:葉綠素;X11:根長。
Note: * and ** indicate significance at 0.05 and 0.01 probability levels, respectively. X1: plant height; X2: biomass; X3: shoot/root ratio; X4: Pn; X5: MDA; X6: relative conductivity; X7: soluble protein content; X8: POD; X9: SOD; X10: chl (a+b); X11: Plant root.
方差分析結果表明(表3),11項指標在3個玉米自交系試驗材料和不同鹽濃度間呈顯著或極顯著差異。3個試驗材料間存在顯著差異才可進行主成分分析,因此本試驗的11項耐鹽指標可以作主成分分析。
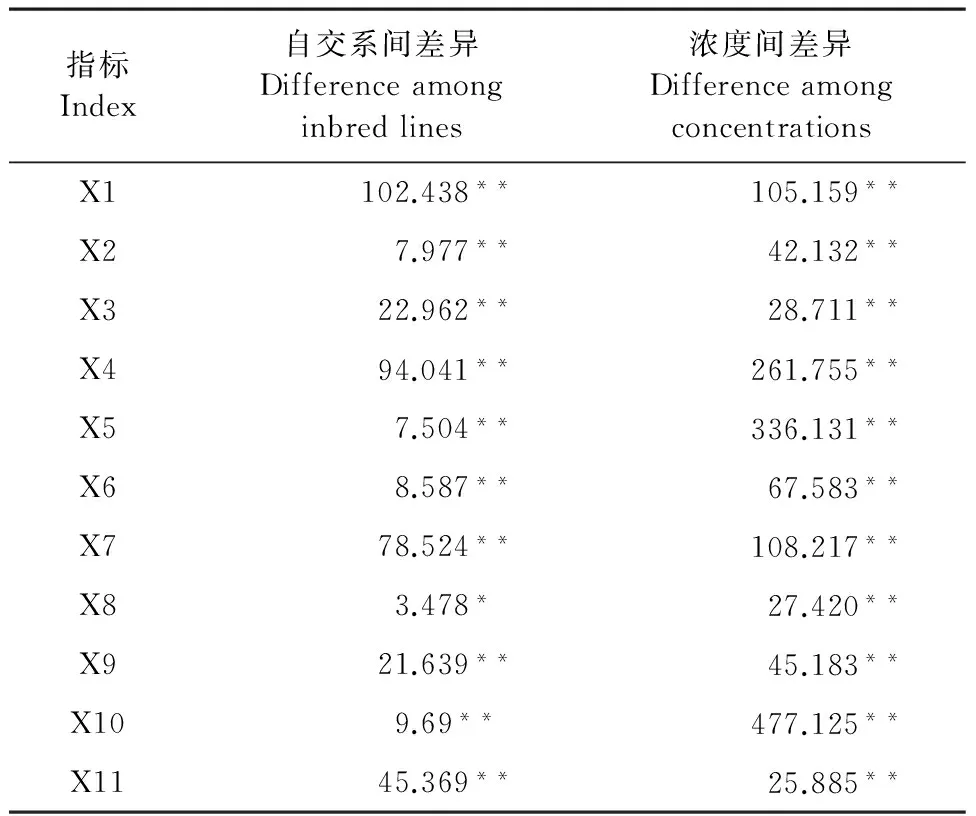
表3 3種玉米自交系耐鹽性狀的11個指標 方差分析及F值檢驗
注:F檢驗,*表示差異顯著(P<0.05);**表示差異極顯著(P<0.01)。
Note:Ftest; * and ** indicate significant difference at 0.05 and 0.01 level, respectively.
2.5 耐鹽指標間主成分分析
對3個玉米自交系不同濃度NaCl脅迫的11個指標進行主成分分析(表4)。前三個主成分的累積貢獻率達到93.959%,表明前3個主成分已經對3個玉米自交系的耐鹽指標作出了充分的概括。由表4、5可知,第一主成分的特征值為7.570,貢獻率為68.815%,其中株高、MDA、Chl(a+b)、Pn、 SOD和POD有絕對值較大的得分系數,除株高、Pn、和Chl(a+b)負向標外,其余均為正向標,第一主成分主要反映地上部生長和生理狀況;第二主成分的特征值為2.185,貢獻率為19.864%,其中根冠比、根長和生物量有絕對值較大的得分系數,均為正向坐標,根長擁有絕對值最大的得分系數,第二主成分與地下部生物量因子密切相關;第三主成分的特征值為0.581,貢獻率為5.280%,其中電導率有絕對值較大的得分系數,第三主成分主要反映與膜透性相關的因子。
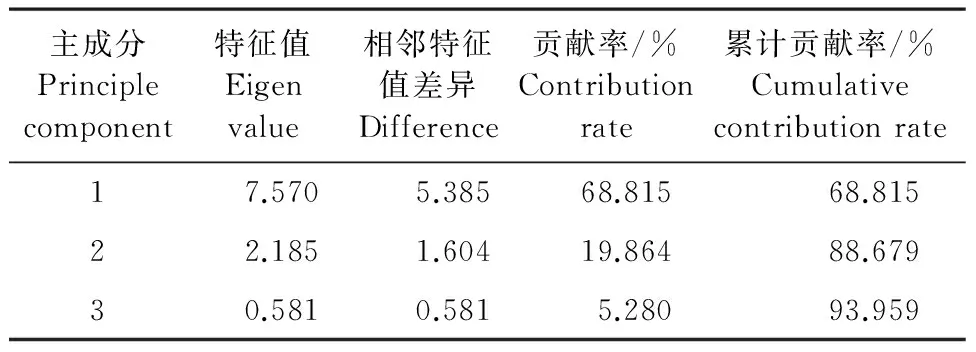
表4 耐鹽指標篩選主成分分析
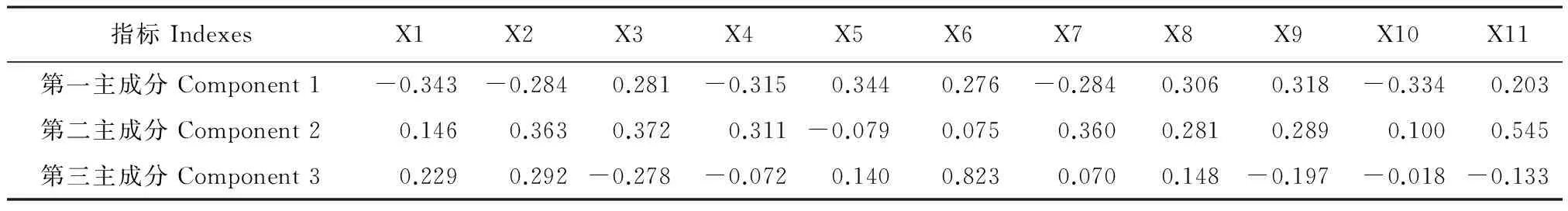
表5 主成分因子得分系數
2.6 隸屬函數分析
主成分分析中各特征向量表示各指標相對于綜合指標貢獻的大小,特征向量的大小是植物耐鹽性評價指標選擇的重要依據。第一主成分和第二主成分累積貢獻率為84.349%,表明前2個主成分反映了3個玉米自交系耐鹽能力的84.349%的信息。第一、二主成分中特征向量較大的有株高、MDA、POD、SOD、Chl(a+b)、Pn、根冠比、根長和生物量9個指標,然后對篩選出的9個指標進行隸屬函數分析,求平均值并評價3個玉米自交系的耐鹽性。根據隸屬函數平均值的大小對3個玉米自交系的耐鹽能力進行排序(表6),3個玉米自交系玉米耐鹽能力由強到弱的順序為:M-2導入系、Hr30導入系、Mo17導入系。
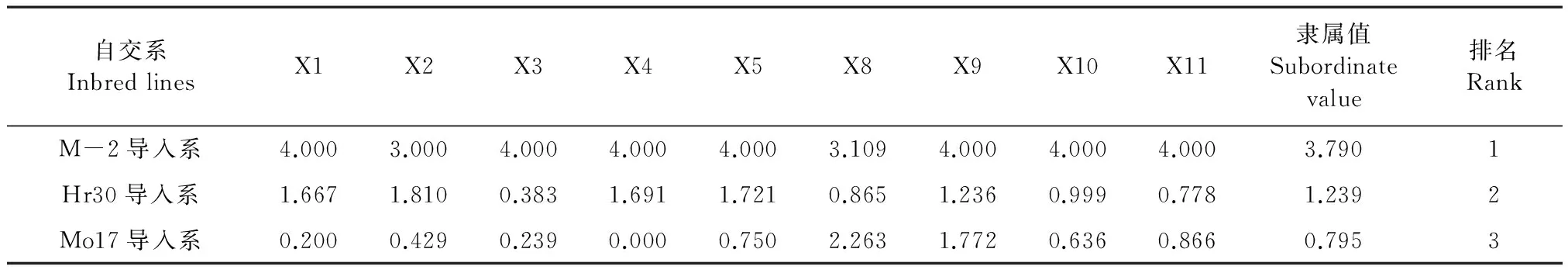
表6 鹽脅迫條件下玉米隸屬函數值
3 結論與討論
植物形態的變化是反映鹽害最直觀的指標[18],合適的篩選壓是植物抗逆篩選的基礎。本試驗中除根冠比的變化率(相對變化率=脅迫后的測定值/對照)在4個鹽脅迫下3個自交系間差異均不顯著外,在250 mmol·L-1鹽脅迫下,株高、生物量和根長的變化率在3個玉米自交系間的差異顯著性最高,所以本試驗設計的鹽脅迫梯度中,250 mmol·L-1可以作為玉米苗期耐鹽性篩選的最適濃度。由于土壤中過度鹽分的存在,導致土壤的滲透勢過高,會損傷葉片中具有蒸騰作用的細胞,降低植物對水分吸收的能力,抑制了植物的正常生長發育[19]。本研究中,三個玉米自交系的株高和根冠比大體上隨著鹽濃度的增加而降低,相比較根系受鹽脅迫影響較小,在低鹽濃度下反而會促進根系的生長發育,說明玉米的地上部對鹽脅迫敏感,比地下部更容易受到鹽脅迫的傷害。有研究表明,鹽脅迫通常會破壞植物細胞膜透性和完整性,引起脂膜過氧化和膜透性增大[20-21]。本研究中3個玉米自交系的MDA和相對電導率均隨著鹽脅迫濃度的增加呈先升高后降低的趨勢,M-2的變化幅度較小,說明M-2的膜質過氧化程度和細胞膜受損傷程度均小于其他兩個自交系。此外,鹽脅迫會使植物的葉綠體色素系統受損,導致葉綠素酶活性增強,迫使葉綠素不斷降解,進而抑制植物的光合作用[22]。本試驗的3個玉米自交系的葉綠素含量和凈光合速率均隨著鹽濃度的增加而下降,這與鄭世英[23]等研究結果一致。鹽脅迫導致植物體內產生過量的活性氧,活性氧大量增加還會損壞蛋白質的分子結構,使蛋白質合成受阻[24],本研究中三個玉米自交系的可溶性蛋白質大體上隨著鹽濃度的增加而逐漸降低。同時,植物會產生大量抗氧化酶來清除多余的活性氧,以此來形成植物體內活性氧的生成和清除之間的動態平衡,減少鹽脅迫對植物造成的氧化損傷[25-26]。付艷[27]等研究表明,隨著鹽濃度的增加,玉米幼苗的POD和SOD活性均呈先升后降的趨勢,本研究得到相似結果,這可能與活性氧的大量產生超出了抗氧化酶系統的清除能力有關。本試驗中的3個自交系的兩種酶活性達到最大值的鹽處理濃度不同,因此不同品種對活性氧的清除能力也存在一定差異。
通常植物耐鹽的機理相對復雜,采用多種生理指標主成分分析和隸屬函數法相結合的方法可以充分挖掘各指標間更深層的關系,避免了單一生理指標評價的片面性。戴海芳[28]等通過主成分分析將鹽脅迫下棉花幼苗葉片的11項生理指標轉換成6個彼此獨立的指標,通過隸屬函數分析得到不同幼苗的耐鹽性的綜合評價值。劉春榮等[29]同樣結合主成分分析和隸屬函數法對玉米苗期的生長指標進行耐鹽性評價。本試驗運用主成分分析歸納出株高、Pn、MDA等9個關鍵篩選指標,因此鹽脅迫下株高、生物量、根冠比和根長可以作為苗期玉米耐鹽性鑒定及篩選的形態指標,4個鹽脅迫下除根冠比的變化率(相對變化率=脅迫后的測定值/對照)在3個自交系間的差異顯著性均不明顯外,250 mmol·L-1的鹽脅迫下株高、生物量和根長的變化率在3個玉米自交系間的差異顯著性均高于其余三個濃度,所以本試驗設計的鹽濃度梯度中,250 mmol·L-1可以作為耐鹽性評價的最適濃度。根據篩選的指標應用隸屬函數法對3個玉米自交系進行綜合評價,求出每個自交系9項生理指標的隸屬平均值,平均值越大,耐鹽性就越強。模糊數學隸屬函數分析得出,3個玉米自交系玉米耐鹽能力由強到弱的順序為:M-2導入系、Hr30導入系、Mo17導入系。
[1] Schwabe K A, Kan I, Knapp K C. Drainwater management for salinity mitigation in irrigated agriculture[J]. American Journal of Agricultural Economics, 2006,88(1):133-149.
[2] International Grains Council. Grain Market: report of IGC Grains conference[R]. London: IGC, 2012.
[3] Heng L K, Hsiao T, Evett S, et al. Validating the FAO aqua crop model for irrigated and water deficient field maize[J]. Agronomy Journal, 2009,101(3):488-498.
[4] Bao Y, Zhao R, Li F, et al. Simultaneous expression of spinacia oleracea chloroplast choline monooxygenase (CMO) and betaine aldehyde dehydrogenase (BADH) genes contribute to dwarfism in transgenic lolium perenne[J]. Plant Molecular Biology Reporter, 2011,29(2):379-388.
[5] Ahmad P, Jaleel C A, Salem M A, et al. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress[J]. Critical Reviews in Biotechnology, 2010,30(3):161-175.
[6] Rubio M C, Pilar B S, Clemente M R, et al. Effects of salt stress on the expression of antioxidant genes and proteins in the model legume Lotus japonicus[J]. New Phytologist, 2009,181(4):851-859.
[7] Ashraf M, Akram N A. Improving salinity tolerance of plants through conventional breeding and genetic engineering: An analytical comparison. Biotechnol Adv[J]. Biotechnology Advances, 2009,27(6):744-752.
[8] Kachout S S, Hamza K J, Bouraoui N K, et al. Salt-Induced changes in antioxidative enzyme activities in shoot tissues of two atriplex varieties[J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2013,41(1):115-121.
[10] 付長方,張海艷.鹽脅迫對玉米種子萌發、幼苗葉綠素含量和滲透勢的影響[J].山東農業科學,2015,5(5):27-30.
[11] 彭云玲,李偉麗,王坤澤,等.NaCl脅迫對玉米耐鹽系與鹽敏感系萌發和幼苗生長的影響[J].草業學報,2012,(4):62-71.
[12] Wellburn A R. The spectra determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution[J]. Journal of Plant Physiology, 1994,144(3):307-313.
[13] Lutts S, Kinet J M, Bouharmont J. NaCl-induced senescence in leaves of rice(Oryza sativaL.) cultivars differing in salinity resistance[J]. Annals of Botany, 1996,78(3):389-398.
[14] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000:258-260.
[15] Giannopolitis C N, Ries S K. Superoxide dismutases: II. purification and quantitative relationship with water-soluble protein in seedlings[J]. Plant Physiology, 1977,59(2):481-484.
[16] Rao M V, Paliyath G, Ormrod D P, et al. Influence of salicylic acid on H2O2production, oxidative stress, and H2O2-metabolizing enzymes. Salicylic acid-mediated oxidative damage requires H2O2[J]. Plant Physiology, 1997,115(1):137-149.
[17] 郝再彬,蒼 晶,徐 仲.植物生理實驗[M].哈爾濱:哈爾濱工業大學出版社,2006.
[18] 王景艷,劉兆普,劉 玲,等.NaCl脅迫對長春花幼苗離子分布和光合作用的影響[J].生態學雜志,2008,(10):1680-1684.
[19] Munns R. Genes and salt tolerance: Bringing them together[J]. New Phytologist, 2005,(167):645-663.
[20] Singh P K, Singh R, Singh S. Cinnamic acid induced changes in reactive oxygen species scavenging enzymes and protein profile in maize (ZeamaysL.) plants grown under salt stress[J]. Physiology & Molecular Biology of Plants, 2013,19(1):53-59.
[21] 趙小強,彭云玲,李健英,等.16份玉米自交系的耐鹽性評價[J].干旱地區農業研究,2014,32(5):40-45.
[22] 杜 菲,陳 新,楊春華,等.NaCl脅迫對不同柳枝稷材料種子萌發與幼苗生長的影響[J].草地學報,2011,(6):1018-1024.
[23] 鄭世英,商學芳,王麗燕,等.鹽脅迫對不同基因型玉米生理特性和產量的影響[J].干旱地區農業研究,2010,28(2):109-112.
[24] 張福鎖.環境脅迫與植物育種[M].北京:農業出版社,1993.
[25] Shahid M A, Balal R M, Pervez M A, et al. Differential response of pea (PisumsativumL.) genotypes to salt stress in relation to the growth, physiological attributes antioxidant activity and organic solutes[J]. Australian Journal of Crop Science, 2012,6(6):828-838.
[26] 魯向暉,穆興民,VinayNangia,等.DSSAT模型對豫西冬小麥保護性耕作效應模擬效果驗證[J].干旱地區農業研究,2010,28(3):64-70.
[27] 付 艷,高樹仁,楊克軍,等.鹽脅迫對玉米耐鹽系與鹽敏感系苗期幾個生理生化指標的影響[J].植物生理學報,2011,(5):459-462.
[28] 戴海芳,武 輝,阿曼古麗,等.不同基因型棉花苗期耐鹽性分析及其鑒定指標篩選[J].中國農業科學,2014,(7):1290-1300.
[29] 劉春榮,張國新,王秀萍.主成分分析及隸屬函數法綜合評價玉米苗期耐鹽性[J].安徽農業科學,2015,(28):13-14.
Physiological responses and tolerance of three maize inbred lines to salt stress
WANG Jing-ze, GAO Shu-ren, SUN Li-fang, WANG Xia, WANG Jing, HU Kai-feng, DENG Jie
(CollegeofAgronomy,HeilongjiangBayiAgriculturalUniversity,Daqing,Heilongjiang163319,China)
Using three maize inbred lines (including M-2 introgression line, Hr30 introgression line, and Mo17 introgression line) as test materials, an experiment was conducted to investigate the effects of different concentrations of NaCl(0 mmol·L-1, 100 mmol·L-1, 150 mmol·L-1, 200 mmol·L-1, and 250 mmol·L-1)on physiological parameters such as superoxide dismutase (SOD) activity, peroxidase (POD) activity and malondialdehyde (MDA) content in maize seedlings. Furthermore, the salt tolerance of these three inbred lines was comprehensively evaluated by the methods of principal components analysis and fuzzy subordinate function, so as to lay foundation of identifying salt tolerance and selecting salt-tolerant cultivars of maize. The results showed that, with the increase of salt concentration, the plant height, biomass, chlorophyll content, net photosynthetic rate (Pn) and soluble protein content in all three inbred lines tended to decrease, while the root length, relative conductivity, MDA content, SOD activity, POD activity and root/shoot ratio were increased at first and decreased subsequently. According to the results of principal components analysis, 9 parameters (including plant height,Pn, MDA content, SOD activity and so on) were closely related to salt tolerance. Through the analysis of fuzzy subordinate function, the salt-resistant capability of tested inbred lines was ranked as: M-2 introgression line>Hr30 introgression line>Mo17 introgression line.
maize; salt; subordinate function
1000-7601(2017)02-0089-07
10.7606/j.issn.1000-7601.2017.02.15
2015-12-01基金項目:黑龍江八一農墾大學研究生創新項目(YJSCX2015-Y05);農業部公益性項目(201303007)

高樹仁,教授,主要從事玉米遺傳育種的研究工作。 E-mail:gaoshr107@126.com。
S332.1
A
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
