首次自褐家鼠糞便標(biāo)本中檢出香港海鷗菌
2017-04-26 08:51:22劉優(yōu)招陳煒聰劉志華韓宇宇
中國(guó)人獸共患病學(xué)報(bào) 2017年4期
關(guān)鍵詞:耐藥
劉優(yōu)招,陳煒聰,劉志華,王 麗,葛 婧,韓宇宇,陳 清,胡 靜
?
首次自褐家鼠糞便標(biāo)本中檢出香港海鷗菌
劉優(yōu)招1,陳煒聰2,劉志華3,王 麗1,葛 婧1,韓宇宇2,陳 清1,胡 靜1
目的 了解褐家鼠糞便標(biāo)本中是否攜帶香港海鷗菌,并分析其耐藥特征及分子分型。方法 2015年6月-2016年5月,利用籠捕法于廣州市南方醫(yī)院及其周邊居民區(qū)捕獲褐家鼠,其糞便樣本經(jīng)增菌后接種改良頭孢哌酮MacConkey瓊脂(CMA)平板培養(yǎng)分離可疑菌株,經(jīng)生化試驗(yàn)及16S rRNA測(cè)序確認(rèn)。采用K-B法進(jìn)行藥物敏感性測(cè)定,并運(yùn)用多位點(diǎn)序列分型進(jìn)行分子分型分析。結(jié)果 共捕獲褐家鼠191只,并在2份褐家鼠糞便標(biāo)本中檢出香港海鷗菌,檢出率為1.05%。兩株菌16S rRNA的測(cè)序結(jié)果與香港海鷗菌標(biāo)準(zhǔn)株HKU1的一致性達(dá)100%。藥敏試驗(yàn)顯示兩株分離株均對(duì)頭孢菌素類抗生素及利福平耐藥。多位點(diǎn)序列分型結(jié)果顯示兩株菌分別為2個(gè)新的ST型:ST-163和ST-164。結(jié)論 褐家鼠糞便中存在香港海鷗菌污染。褐家鼠與人類生活關(guān)系密切,可能為人類感染的的另一潛在來源。
香港海鷗菌;褐家鼠;16S rRNA測(cè)序;多位點(diǎn)序列分型
香港海鷗菌(Laribacterhongkongensis,Lh)是一種可能致人類嚴(yán)重腹瀉的新發(fā)現(xiàn)病原菌。該菌2001年首次在香港肝硬化病人的胸腔膿血中分離到并被鑒定為變形菌門(Proteobaxteria)、β-變形菌綱(β-subclass Proteobacteria)、奈瑟菌科(Neisseriaceae)的一個(gè)新屬[1]。
香港海鷗菌作為一個(gè)新發(fā)現(xiàn)的病原菌,其可能的源頭、存在哪些宿主、造成人類感染的方式,不同來源菌株間的遺傳結(jié)構(gòu)和耐藥性等問題都亟待解決。到目前為止,香港海鷗菌除造成人類感染外,主要在淡水魚和虎紋蛙等淡水生物的腸道標(biāo)本中檢出[2-4],被認(rèn)為是該菌的主要宿主,進(jìn)食淡水魚類和異地旅游被證實(shí)是感染該菌的危險(xiǎn)因素[5]。但另有研究顯示,有些感染香港海鷗菌的病人在感染近期均無進(jìn)食魚類史和異地旅游史,其感染來源和途徑尚不清楚[6-7],是否存在其他宿主和危險(xiǎn)因素仍有待進(jìn)一步研究。
褐家鼠(Rattusnorvegicus)廣泛分布于人類的生活環(huán)境中,與人密切接觸,是許多病原菌的儲(chǔ)存宿主,如鉤端螺旋體[8],鼠疫耶爾森菌[9],傷寒立克次體[10],沙門菌[11]和空腸彎曲菌[12]。為了解褐家鼠糞便中是否攜帶香港海鷗菌,于2015年6月-2016年5月在廣州市某綜合三甲醫(yī)院周圍及附近居民區(qū)捕獲褐家鼠,采集褐家鼠糞便標(biāo)本進(jìn)行香港海鷗菌的分離培養(yǎng)和鑒定,并對(duì)陽(yáng)性菌進(jìn)行藥物敏感性測(cè)定和多位點(diǎn)序列分型(multi-loci sequence typing,MLST)分析。
1 材料與方法
1.1 樣本采集 2015年6月-2016年5月,按照《中華人民共和國(guó)出入境檢驗(yàn)檢疫行業(yè)標(biāo)準(zhǔn)》(SN/T 2788—2011)相關(guān)標(biāo)準(zhǔn)在廣東省廣州市南方醫(yī)院周圍及附近居民區(qū)利用籠捕法捕鼠。鼠籠布置地點(diǎn)主要為倉(cāng)庫(kù)、廚房、下水道、垃圾箱、建筑物墻邊等,以“晚放晨收”形式每月捕鼠一次,連續(xù)捕鼠一年。捕獲的老鼠立即運(yùn)送至實(shí)驗(yàn)室,按照《實(shí)驗(yàn)動(dòng)物細(xì)菌學(xué)檢測(cè)標(biāo)本采集》(GB/T—14926.42—2001)要求解剖并收集糞便樣本。
1.2 主要試劑 營(yíng)養(yǎng)肉湯、營(yíng)養(yǎng)瓊脂、麥康凱瓊脂(CMA)、MH藥敏實(shí)驗(yàn)瓊脂、三糖鐵瓊脂、脲酶及精氨酸雙水解酶生化管均購(gòu)自北京路橋技術(shù)股份有限公司;頭孢哌酮、氧化酶試紙、觸酶及K-B法藥物敏感紙片購(gòu)自英國(guó)Oxoid公司;細(xì)菌基因組DNA提取試劑盒購(gòu)自北京天根生化科技有限公司;引物合成及測(cè)序由深圳華大基因公司完成。
1.3 試驗(yàn)方法
1.3.1 香港海鷗菌的分離培養(yǎng) 將糞便樣本置于營(yíng)養(yǎng)肉湯中,37 ℃增菌18~24 h。將增菌肉湯劃線接種于改良頭孢哌酮MacConkey瓊脂(CMA),頭孢哌酮濃度為16 μg/mL。CMA平皿置于37 ℃培養(yǎng)箱培養(yǎng)48 h。挑取香港海鷗菌可疑菌落(直徑在1.0~1.5 mm之間,淺灰色、半透明圓形、中間略凸起、光滑)接種于營(yíng)養(yǎng)瓊脂平板分純培養(yǎng),置37 ℃培養(yǎng)18~24 h后進(jìn)行生化試驗(yàn)。對(duì)氧化酶、觸酶、脲酶、及精氨酸雙水解酶陽(yáng)性,三糖鐵陰性的菌株再進(jìn)行16S rRNA測(cè)序確認(rèn)。HKU1和HLHK9 菌株(均由香港大學(xué)微生物學(xué)系袁國(guó)勇教授惠贈(zèng))為陽(yáng)性對(duì)照菌株,大腸埃希菌ATCC25922(南方醫(yī)科大學(xué)流行病學(xué)實(shí)驗(yàn)室保存)為陰性對(duì)照菌株。
1.3.2 16S rRNA擴(kuò)增及測(cè)序 細(xì)菌基因組DNA提取試劑盒提取初篩菌株的DNA,置于-20 ℃保存?zhèn)溆谩CR擴(kuò)增16S rRNA的特異性片段,片段大小550 bp。目的片段陽(yáng)性的進(jìn)一步擴(kuò)增16S rRNA全序,擴(kuò)增產(chǎn)物送華大基因公司測(cè)序后利用在線Blast (http://www.ncbi.nlm.nih.gov/BLAST)比對(duì)確認(rèn)。具體引物設(shè)計(jì)及PCR反應(yīng)條件參照文獻(xiàn)[13]。
1.3.3 藥敏實(shí)驗(yàn) 采用紙片擴(kuò)散法(K-B法),按照美國(guó)臨床和實(shí)驗(yàn)室標(biāo)準(zhǔn)協(xié)會(huì)(Clinical and Laboratory Standards Institute, CLSI)標(biāo)準(zhǔn)判讀。抗生素紙片包括:頭孢噻吩(30 μg)、頭孢唑林(30 μg)、頭孢呋辛(30 μg)、頭孢哌酮(75 μg)、頭孢他啶(30 μg)頭孢吡肟(30 μg);氨芐西林(10 μg)、哌拉西林(100 μg);亞胺培南(10 μg)氨曲南(30 μg);慶大霉素(10 μg)、阿米卡星(30 μg)、鏈霉素(10 μg);紅霉素(15 μg);利福平(5 μg);環(huán)丙沙星(5 μg);四環(huán)素(30 μg);氯霉素(30 μg);復(fù)方新諾明(甲氧芐啶1.25 μg+磺胺甲惡唑23.75 μg)。大腸桿菌ATCC25922和金黃色葡萄球菌ATCC25923作為質(zhì)控菌。
1.3.4 MLST分型 根據(jù)香港大學(xué)MLST分型方法[14],選取rho、acnB、ftsH、trpE、ilvC、thiC、eno等7個(gè)管家基因作為MLST分型位點(diǎn)。引物設(shè)計(jì)及PCR反應(yīng)體系及條件同參考文獻(xiàn)[14]。取5 μL PCR擴(kuò)增產(chǎn)物于1%瓊脂糖凝膠經(jīng)100 V電泳25 min, 凝膠成像儀觀察電泳條帶。PCR陽(yáng)性產(chǎn)物送至華大基因進(jìn)行雙向測(cè)序,Chrome軟件查看峰圖并評(píng)估測(cè)序結(jié)果。用DNAman軟件拼接正反序列,得到每個(gè)管家基因的完整序列并將菌株的7個(gè)管家基因序列送至香港大學(xué)與現(xiàn)存的MLST數(shù)據(jù)庫(kù)進(jìn)行比對(duì)分析,得到等位基因號(hào)及ST號(hào),運(yùn)用eBURST、START2等分析軟件進(jìn)行譜系分析和聚類分析。
2 結(jié) 果
2.1 褐家鼠糞便中香港海鷗菌的檢出情況 2015年6月—2016年5月,連續(xù)捕鼠12個(gè)月,共捕獲191只褐家鼠,其中156只成年(118只雄性,38只雌性),35只未成年。在191份糞便標(biāo)本中檢出2份標(biāo)本香港海鷗菌陽(yáng)性,檢出率為1.05%。兩只褐家鼠均為未成年鼠,鼠籠放置地點(diǎn)均在居民區(qū)附近的垃圾箱旁邊,捕獲時(shí)間分別為2015年7月和2015年11月。兩株香港海鷗菌分別命名為R251和R398。
2.2 16S rRNA基因序列分析 PCR擴(kuò)增結(jié)果顯示,不包括引物結(jié)合區(qū),菌株R251和R398的16S rRNA 基因序列在1 400 bp左右,兩者之間無差異。Blast比對(duì)結(jié)果顯示:R251和R398與香港海鷗菌標(biāo)準(zhǔn)株HKU1(GenBank 登錄號(hào):AF389085)相應(yīng)的序列一致性達(dá)100%,與已全基因組測(cè)序菌株HLHK9(GenBank登錄號(hào):NR_074669)相應(yīng)的序列存在2個(gè)堿基差異,一致性為99%。
2.3 藥敏結(jié)果 R251和R398對(duì)10個(gè)種類常用抗生素中的19中抗菌藥表現(xiàn)出相似的耐藥性,均對(duì)頭孢呋辛、頭孢吡肟、哌拉西林、亞胺培南、氨曲南、慶大霉素、阿米卡星、鏈霉素、紅霉素、環(huán)丙沙星、四環(huán)素、氯霉素及復(fù)方新諾明敏感;對(duì)頭孢噻吩、頭孢唑啉、頭孢哌酮及利福平表現(xiàn)為耐藥。此外,R251對(duì)頭孢他啶及氨芐西林敏感,但R398表現(xiàn)為耐藥。
2.4 MLST分型 菌株R251的7個(gè)管家基因(rho、acnB、ftsH、trpE、ilvC、thiC、eno)的等位基因號(hào)分別為:14、18、19、18、13、18、20。菌株R398的則為:42、18、19、49、35、23、20。由7個(gè)等位基因號(hào)組成等位基因譜,分別得到相應(yīng)的ST型:ST-163和ST-164。如圖1所示,eBURST軟件對(duì)香港海鷗菌的等位基因號(hào)及ST型號(hào)進(jìn)行分析,將ST型分成12組克隆復(fù)合體。ST-163和ST-164分散存在,不與其他ST型形成克隆復(fù)合體。START2的聚類結(jié)果(圖2)也顯示:菌株R251和R398不與其他ST聚在一起,但兩株鼠源分離株與人源分離株的親緣關(guān)系較近。
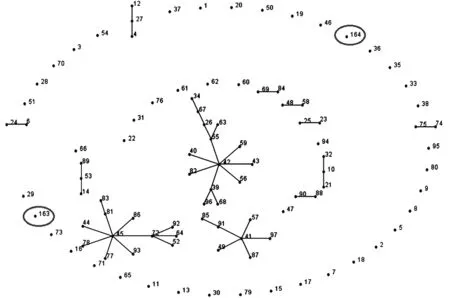
每個(gè)數(shù)字代表一個(gè)ST型,連接2個(gè)ST型之間的直線表示兩個(gè)ST型的7對(duì)等位基因中至少有6對(duì)是相同的。Each number represents a MLST sequence type (ST) and each line connects STs that are identical in at least six out of the seven housekeeping genes.圖1 eBURST 分析香港海鷗菌ST型分布圖Fig.1 eBURST analyses of L. hongkongensis strains
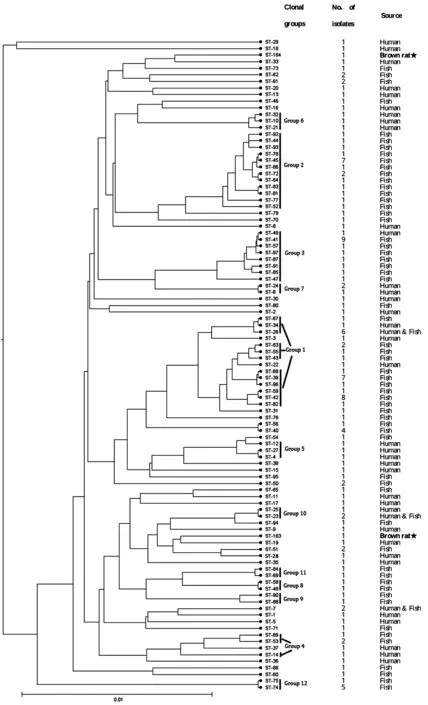
圖2 香港海鷗菌的聚類分析Fig.2 Cluster analysis for L.hongkongensis strains
3 討 論
香港海鷗菌由香港學(xué)者Yuen等[1]于2001年從一名肝硬化病人的胸腔膿血分離,后研究表明此菌可致人類腹瀉且食用淡水魚為其危險(xiǎn)因素之一[5]。此外,香港海鷗菌還可能是造成侵入性感染的致病菌,如菌血癥[15-16]和腹膜炎[17]。自從香港海鷗菌發(fā)現(xiàn)至今,除病人以外,僅在淡水魚[2, 4, 18]、可食用蛙[2-4]、水庫(kù)水[19]等淡水生物或環(huán)境中檢測(cè)出該菌,但在海產(chǎn)品、豬、牛、雞、鴨、鵝等糞便或肛拭子標(biāo)本中均未檢測(cè)到該菌的存在[18]。本研究首次發(fā)現(xiàn)作為哺乳動(dòng)物的褐家鼠也能攜帶香港海鷗菌,該結(jié)果提示褐家鼠可能是除人和淡水生物以外的另一個(gè)香港海鷗菌的潛在宿主。
褐家鼠為雜食性動(dòng)物,食譜廣而雜。在居民區(qū)室內(nèi),喜吃肉類、蔬菜等;在野外,常以動(dòng)物性食物為主要食料,捕食小魚、蝦、蛙類等。其棲息場(chǎng)所主要在廚房、倉(cāng)庫(kù)、下水道、垃圾堆等雜亂無章處[20]。褐家鼠的生活習(xí)性為感染香港海鷗菌提供可能,比如攝食被香港海鷗菌污染的魚肉類,或捕食攜帶此菌的小魚、蛙類等。
我國(guó)內(nèi)地報(bào)道,某些感染香港海鷗菌的腹瀉患者發(fā)病前2周并無進(jìn)食魚類史和外出旅游史[6-7],提示除食用淡水魚和異地旅行等危險(xiǎn)因素外,可能還存在感染該菌的其他來源和途徑。本次研究從褐家鼠糞便中分離出香港海鷗菌,提示其可能成為人類感染此菌的的另一潛在來源。香港海鷗菌可隨褐家鼠糞便排出體外,污染食物、飲水及周圍環(huán)境,人類可能通過接觸污染的食物、飲水而感染此菌。除此之外,褐家鼠還可以通過抓傷、咬傷直接接觸人類,傳播多種疾病。雖然本研究中,褐家鼠糞便中香港海鷗菌的檢出率(1.05%)低于淡水魚(16.3%)及可食用蛙(59.5%)的檢出率[2],但褐家鼠活動(dòng)范圍大,又多活動(dòng)于居民區(qū),與居民接觸機(jī)會(huì)多,給人類感染帶來的潛在危險(xiǎn)值得關(guān)注。
兩株香港海鷗菌的鼠分離株R251和R398的藥敏實(shí)驗(yàn)結(jié)果顯示,其對(duì)利福平及一、三代頭孢菌素類藥物均耐藥,對(duì)二、四代頭孢菌類抗生素、其他β-內(nèi)酰胺類和氨基糖苷類抗生素均敏感,與其他宿主來源的分離株基本一致[2, 18]。但是,與其他宿主來源的香港海鷗菌的耐藥譜相比,也存在一定的差異性。鼠來源株(R251和R398)對(duì)紅霉素類、四環(huán)素類、喹諾酮類及磺胺類均表現(xiàn)為敏感,而魚來源株對(duì)紅霉素有一定水平的耐藥,部分人來源株對(duì)紅霉素類和四環(huán)素類耐藥,而蛙來源株對(duì)這4類抗生素均表現(xiàn)為一定水平的耐藥。
目前,已有學(xué)者運(yùn)用脈沖場(chǎng)凝膠電泳(pulsed-field gel electrophoresis, PFGE)技術(shù)[18]及MLST方法[14]進(jìn)行香港海鷗菌的感染溯源研究,對(duì)魚來源株和人來源株分型,發(fā)現(xiàn)大部分人來源株聚在一起,提示一些菌株毒力可能更強(qiáng),但未進(jìn)行驗(yàn)證。本研究的MLST聚類分析提示鼠來源株與人來源株有更近的親緣關(guān)系,但兩者之間的聯(lián)系有待進(jìn)一步研究。到目前為止,香港海鷗菌的致病機(jī)理、菌株間的毒力是否存在差異性仍不明確。動(dòng)物模型是研究腹瀉發(fā)病機(jī)制的重要手段,褐家鼠能攜帶香港海鷗菌提示從褐家鼠分化而來的SD大鼠可能是一個(gè)可以用來嘗試建立此菌感染的動(dòng)物模型。
本研究首次報(bào)道了野生型褐家鼠糞便中存在香港海鷗菌。這是除人群病例以外,第一次在哺乳類動(dòng)物中分離到該菌,擴(kuò)大了對(duì)香港海鷗菌的宿主研究范圍,該菌的傳播途徑、流行范圍的認(rèn)識(shí)。此外,褐家鼠在香港海鷗菌的傳播過程中扮演著何種角色,其可能造成人類感染的方式等問題也值得進(jìn)一步研究。
[1] Yuen KY, Woo PC, Teng JL, et al.Laribacterhongkongensisgen. nov. sp. nov., a novel gram-negative bacterium isolated from a cirrhotic patient with bacteremia and empyema[J]. J Clin Microbiol, 2001, 39(12):4227-4232. DOI:10.1128/JCM.39.12.4227-4232.2001
[2] Feng JL, Hu J, Lin JY, et al. The prevalence, antimicrobial resistance and PFGE profiles ofLaribacterhongkongensisin retail freshwater fish and edible frogs of southern China[J]. Food Microbiol, 2012, 32(1):118-123. DOI:10.1016/j.fm.2012.04.018
[3] Lau SK, Lee LC, Fan RY, et al. Isolation ofLaribacterhongkongensis, a novel bacterium associated with gastroenteritis, from Chinese tiger frog[J]. Int J Food Microbiol, 2009, 129(1):78-82. DOI:10.1016/j.ijfoodmicro.2008.10.021
[4] Feng M, Liu YZ, He YQ, et al. Detection and drug resistance characteristic ofLaribacterhongkongensisin freshwater products in a district of Shenzhen City[J]. Chin J Dis Ctrl Prev, 2016, 20(8):764-767. DOI:10.16462 /j.cnki.zhjbkz.2016.08.003 (in Chinese)
馮梅,劉優(yōu)招,何雅青,等. 深圳市某區(qū)淡水產(chǎn)品中香港海鷗菌的檢測(cè)及耐藥特征分析[J]. 中華疾病控制雜志,2016, 20(8):764-767. DOI:10 16462 /j.cnki.zhjbkz.2016. 08.003
[5] Woo PC, Lau SK, Teng JL, et al. Association ofLaribacterhongkongensisin community-acquired gastroenteritis with travel and eating fish:a multicentre case-control study[J]. Lancet, 2004, 363(9425):1941-1947. DOI:10. 1016/S0140-6736(04)16407-6
[6] Zhu JF, Zheng NC, Liao RY, et al.Laribacterhongkongensisisolated from a patient with conmmunity-acquired gastroenteritis in Guangdong province[J]. Chin J Public Health, 2011, 27(4):403-405. (in Chinese)
朱江峰,鄭南才,廖如燕,等. 廣東省香港海鷗菌感染性腹瀉1例分析[J]. 中國(guó)公共衛(wèi)生,2011, 27(4):403-405.
[7] Ni XP, Ren SH, Sun JR, et al.Laribacterhongkongensisisolated from a patient with community-acquired gastroenteritis in Hangzhou City[J]. J Clin Microbiol, 2007, 45(1):255-256. DOI:10.1128/JCM.01400-06
[8] Evangelista KV, Coburn J.Leptospiraasan emerging pathogen:a review of its biology, pathogenesis and host immune responses[J]. Future Microbiol, 2010, 5(9):1413-1425. DOI:10.2217/fmb.10.102
[9] Chanteau S, Ratsifasoamanana L, Rasoamanana B, et al. Plague, a reemerging disease in Madagascar[J]. Emerg Infect Dis, 1998, 4(1):101-104. DOI:10.3201/eid0401.980114
[10] Billeter SA, Gundi VA, Rood MP, et al. Molecular detection and identification ofBartonellaspecies inXenopsyllacheopisfleas (Siphonaptera:Pulicidae) collected fromRattusnorvegicusrats in Los Angeles, California[J]. Appl Environ Microbiol, 2011, 77(21):7850-7852. DOI:10.1128/AEM.06012-11
[11] Yokoyama E, Maruyama S, Kabeya H, et al. Prevalence and genetic properties ofSalmonellaentericaserovar typhimurium definitive phage type 104 isolated fromRattusnorvegicusandRattusrattushouse rats in Yokohama City, Japan[J]. Appl Environ Microbiol, 2007, 73(8):2624-2630. DOI:10.1128/AEM.02465-06
[12] Nkogwe C, Raletobana J, Stewart-Johnson A, et al. Frequency of detection ofEscherichiacoli,Salmonellaspp., andCampylobacterspp. in the faeces of wild rats(Rattusspp.) in Trinidad and Tobago[J]. Vet Med Int, 2011, 2011:686923. DOI:10.4061/2011/686923
[13] Hu J, Zhu JF, Zheng NC, et al. Detection ofLaribacterhongkongensisin freshwater products in Jiangmen, Guangdong[J]. 2013, 28(7):532-535. (in Chinese)
胡靜,朱江峰,鄭南才,等. 廣東省江門市淡水產(chǎn)品中香港海鷗菌的檢測(cè)[J]. 疾病監(jiān)測(cè), 2013, 28(7):532-535.
[14] Woo PC, Teng JL, Tsang AK, et al. Development of a multi-locus sequence typing scheme forLaribacterhongkongensis, a novel bacterium associated with freshwater fish-borne gastroenteritis and traveler’s diarrhea[J]. BMC Microbiol, 2009, 9:21. DOI:10.1186/1471-2180-9-21
[15] Tse CW, Curreem SO, Cheung I, et al. A novel MLST sequence type discovered in the first fatal case ofLaribacterhongkongensisbacteremia clusters with the sequence types of other human isolates[J]. Emerg Microbes Infect, 2014, 3(6):e41. DOI:10.1038/emi.2014.39
[16] Kim DS, Wi YM, Choi JY, et al. Bacteremia caused byLaribacterhongkongensismisidentified as Acinetobacter lwoffii:report of the first case in Korea[J]. 2011, 26:679-681. DOI:10.3346/jkms.2011.26.5.679
[17] Woo PC, Poon RW, Foo CH, et al. First report ofLaribacterhongkongensisperitonitis in continuous ambulatory peritoneal dialysis[J]. Perit Dial Int, 2016, 36(1):105-107. DOI:10.3747/pdi.2014.00270
[18] Teng JL, Woo PC, Ma SS, et al. Ecoepidemiology ofLaribacterhongkongensis, a novel bacterium associated with gastroenteritis[J]. J Clin Microbiol, 2005, 43(2):919-922. DOI:10.1128/JCM.43.2.919-922.2005
[19] Lau SK, Woo PC, Fan RY, et al. Isolation ofLaribacterhongkongensis, a novel bacterium associated with gastroenteritis, from drinking water reservoirs in Hong Kong[J]. J Appl Microbiol, 2007, 103(3):507-515. DOI:10.1111/j.1365-2672.2006.03263.x
[20] Gao G, Wang SW. Host animals of plague and its prevention in China[M]. Lanzhou:Technology of Gansu Publishing Press, 2012:109-110. (in Chinese)
高共,王升文. 中國(guó)鼠疫宿主動(dòng)物及其防制[M]. 蘭州:甘肅科學(xué)技術(shù)出版社, 2012:109-110.
Hu Jing, Email:hjalzh@smu.edu.cn
First isolation ofLaribacterhongkongensisfrom stool samples ofRattusnorvegicus
LIU You-zhao1, CHEN Wei-cong2, LIU Zhi-hua3, WANG Li1, GE Jing1, HAN Yu-yu2,CHEN Qing1, HU Jing1
(1.DepartmentofEpidemiology,SchoolofPublicHealth,SouthernMedicalUniversity,GuangdongProvincialKeyLaboratoryofTropicalDiseaseResearch,Guangzhou510515,China; 2.SchoolofTraditionalChineseMedicine,SouthernMedicalUniversity,Guangzhou510515,China; 3.DepartmentofInfectiousDiseases,NanfangHospital,SouthernMedicalUniversity,Guangzhou510515,China)
In order to investigate whetherLaribacterhongkongensiscould be detected in stool samples ofRattusnorvegicusin the wild,Rattusnorvegicuswere trapped alive in an urban community of Guangzhou, China over a period of one year from June 2015 to May 2016, and their stool samples were examined for the presence ofL.hongkongensisstrains. Isolates were identified based on phenotypic characteristics and 16S rRNA sequence analysis, and were examined for their susceptibility to 19 antimicrobial agents. Further typing of the isolates was performed using multi-loci sequence typing (MLST) analysis. A total of 191R.norvegicuswere trapped alive.L.hongkongensiswas identified and successfully isolated from two samples, representing a prevalence of 1.05%. Although the two isolates possessed similar phenotypic characteristics and have no base difference of 16S rRNA gene, they constituted two new distinct sequence types (STs), ST-163 and ST-164. This is the first report thatL.hongkongensiscan be detected in the intestinal tract ofR.norvegicus. Results suggest thatR.norvegicuscould serve as carriers ofL.hongkongensisand therefore could be another potential source of infection.
Laribacterhongkongensis;Rattusnorvegicus; 16S rRNA sequence; sequence type
10.3969/j.issn.1002-2694.2017.04.010
國(guó)家自然科學(xué)基金(No.81373052 & 81373051)資助
胡 靜, Email:hjalzh@smu.edu.cn
1.南方醫(yī)科大學(xué)公共衛(wèi)生學(xué)院流行病學(xué)系,廣州 510515; 2.南方醫(yī)科大學(xué)中醫(yī)藥學(xué)院中西醫(yī)臨床,廣州 510515; 3.南方醫(yī)科大學(xué)南方醫(yī)院感染內(nèi)科,廣州 510515
Supported by the National Science Foundation of China (Nos. 81373052 & 81373051)
R378
A
1002-2694(2017)04-0343-06
2016-09-30 編輯:梁小潔
猜你喜歡
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:40
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年3期)2021-07-21 09:04:02
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
云南醫(yī)藥(2019年3期)2019-07-25 07:25:10
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2016年1期)2016-11-12 13:19:40
國(guó)外醫(yī)藥(抗生素分冊(cè))(2016年6期)2016-07-10 11:34:45
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年14期)2016-01-15 02:58:37
中國(guó)當(dāng)代醫(yī)藥(2015年17期)2015-03-01 02:03:58
