不同濃度氧暴露對早產兒外周血單個核細胞SIRT1通路的影響
2017-04-14 03:06:32劉興鈴董文斌李清平雷小平張蓮玉盧佑英趙帥
山東醫藥 2017年14期
關鍵詞:氧化應激
劉興鈴,董文斌,李清平,雷小平,張蓮玉,盧佑英,趙帥
(西南醫科大學附屬醫院,四川瀘州646000)
胎齡<37周出生的活產嬰兒稱為早產兒或未成熟兒。由于早產兒肺臟發育常不成熟,多需要進行呼吸支持,而隨著機械通氣和吸氧治療(以下簡稱氧療)時間的延長,可導致早產兒視網膜病甚至早產兒支氣管肺發育不良(BPD)的發生[1]。活性氧簇(ROS)是細胞線粒體呼吸過程中產生的一個導致氧化損傷和老化的破壞性產物,是氧化應激反應過程中的重要指標。本課題組前期研究證實[2],高濃度的氧暴露可誘導早產兒外周血單個核細胞(PBMCs)中產生大量的ROS,促進氧化應激反應的發生,但其機制尚不明確。沉默信息調節因子2相關酶1(SIRT1)是一種應激反應性分子,作為底物參與SUMO化反應,發揮其去乙酰化的保護作用,避免氧化應激等損傷[3]。SUMO特異性蛋白酶家族(SENPs)參與SUMO蛋白共價結合(被切斷)到靶蛋白賴氨酸殘基上的動態可逆過程,目前研究較多的是SENP1, SIRT1又可以被SENP1去SUMO化,抑制SIRT1的去乙酰化活性[3]。p53是SIRT1的非組蛋白底物,乙酰化p53的促凋亡作用可以被SIRT1的去乙酰化作用緩解,達到抑制凋亡的作用[4]。目前,對SIRT1信號通路在高氧對低體質量早產兒氧化應激損傷中的作用尚不清楚。2015年6月~2016年7月,我們對80例低體質量早產兒采用不同濃度氧療,觀察PBMCs內SIRT1通路相關蛋白的表達變化,探討氧暴露對早產兒氧化應激損傷的影響及其機制,為BPD的防治提供理論依據。
1 資料與方法
1.1 試劑及儀器 人淋巴細胞分離液(灝洋生物制品科技有限責任公司);MitoSOXTMRed(Invitrogen公司);DAPI(北京索萊寶公司);抗熒光淬滅封片劑(碧云天公司);兔抗人SIRT1抗體(Abcam公司);二抗羊抗兔IgG、羊抗小鼠IgG(Abcam公司);5×SDS上樣緩沖液(上海生工);蛋白裂解液(美國碧云天公司);內參一抗(Abcam公司);牛血清白蛋白(Amersham Pharmacia Biotech公司,美國);PVDF膜(Bio-Rad公司,美國)等。低溫高速離心機GS-15R(Beckman公司);超凈工作臺(Thermo公司);倒置相差顯微鏡(Olympus公司);TCSSP5激光共聚焦顯微鏡(Leica公司);全光譜分光光度儀(Eppendorf公司);GDS8000凝膠掃描系統(UVP公司)。
1.2 臨床資料 選取2015年6月~2016年7月生后即入住我院新生兒科的低體質量早產兒80例,男44例、女36例,胎齡(29.82±0.79)周,入院時間為生后(4.51±2.21)h,出生體質量(1 152±117)g。納入標準:胎齡<32周;出生體質量<1 500 g[11];入院時診斷為新生兒呼吸窘迫綜合征且需要立即氧療;出生24 h內入院且存活至生后住院時間28 d以上。排除標準:合并復雜型先天性心臟病及其他先天畸形。按氧暴露水平進行前瞻性隊列分組,根據吸氧濃度分為對照組、低濃度吸氧組、中濃度吸氧組、高濃度吸氧組各20例。四組患兒性別、胎齡、出生體質量差異無統計學意義,具有可比性。本研究經我院倫理委員會批準,征得家屬同意并簽署知情同意書。
1.3 氧療方法 四組均于出生后即開始進行氧療,采用低氧無創CPAP輔助通氣或有創呼吸機機械通氣(氣管插管)。對照組吸氧濃度FiO2=21%或以FiO2>21%吸氧,但總吸氧時間<24 h;低濃度吸氧組FiO2>21%,吸氧時間>24 h,或因病情需要FiO2>30%的吸氧時間<24 h;中濃度吸氧組FiO2為30%~<40%,吸氧時間>24 h,或因病情需要FiO2>40%的吸氧時間<24 h;高濃度吸氧組為FiO2≥40%且吸氧時間>24 h。
1.4 實驗方法
1.4.1 PBMCs分離 分別于生后0、7、14、28 d時,均經橈動脈采血2 mL,置于EDTA抗凝管中,采用Ficoll密度梯度離心法分離PBMCs。
1.4.2 PBMCs中ROS水平檢測 采用激光共聚焦顯微鏡技術。將分離出的PBMCs調整細胞數為2×106/mL。取200 μL細胞懸液小心均勻涂于黏附載玻片上待干;滴加40 g/L多聚甲醛固定20 min,按說明書配制Mito SOX探針,滴加800 μL均勻覆蓋于細胞層上,37 ℃濕盒孵育30 min。取10 μg/mL DAPI 80 μL藍色熒光染料染細胞核5 min,滴加20 μL抗熒光淬滅劑封片。采用激光共聚焦顯微鏡檢測紅色熒光(激發波長510 nm、發射波長580 nm),采用Image Pro Plus6.0圖像分析軟件對圖像進行分析處理,測定平均熒光強度,從而反映ROS水平。
1.4.3 PBMCs中SIRT1定位檢測 采用激光共聚焦顯微鏡技術。細胞涂片和固定同上,滴加80 μL 0.2%TritonX-100室溫下穿透細胞15 min,滴加正常山羊血清工作液,滴加抗體稀釋液稀釋的兔抗人SIRT1抗體(1∶80), 4 ℃孵育過夜。避光滴加FITC記山羊抗小兔IgG(1∶800),37 ℃孵育1 h。DAPI染核,抗熒光淬滅劑封片,用激光共聚焦顯微鏡檢測紅色熒光(激發波長488 nm),反映SIRT1定位情況[12]。DAPI染細胞核標記藍色熒光,兩種熒光合成為紫色熒光。運用Image Pro Plus 中的line profile測定細胞內紫色熒光強度分布,出現熒光強度向靠近胞膜側增高而在胞核中降低的細胞即為發生SIRT1轉位的陽性細胞。取8個視野,以陽性細胞數占細胞總數的構成比計算轉位率。
1.4.4 PBMCs內胞核SIRT1蛋白表達檢測 采用Western blot法。懸浮細胞離心,棄上清留下細胞沉淀,PMSF的細胞質蛋白抽提試劑A,把細胞沉淀完全懸浮并分散開,冰浴;加入細胞質蛋白抽提試劑B 10 μL,得到細胞質蛋白。添加PMSF細胞核蛋白抽提試劑,震蕩、離心后得到細胞核蛋白。SDS-PAGE電泳、封閉,取出聚偏二氟乙烯膜,顯影后晾干。采用UVP凝膠圖像處理系統Lakworks4.6軟件分析目的條帶的灰度值,結果以胞核SIRT1/GAPDH表示。
1.4.5 PBMCs內SENP1、乙酰化p53蛋白表達檢測 采用Western blot法。淋巴細胞分離,收集細胞離心,加入細胞裂解液(RIPA),充分振蕩,離心吸取上清,Bradford方法蛋白定量后加入等體積2×上樣緩沖液,煮沸5 min,短暫離心后-20 ℃保存。取50 μg蛋白樣品行8% SDSPAGE電泳。用濕轉法將膠上蛋白轉移至NC膜上,加入鼠源一抗,4 ℃過夜后,加入HRP標記的兔抗鼠二抗。緩沖液沖洗后,進入暗室,顯影、晾干后,采用UVP凝膠圖像處理系統Lakworks4.6軟件分析目的條帶的灰度值,結果以SIRT1、乙酰化p53與GAPDH的比值表示蛋白表達量。
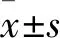
2 結果
2.1 各組不同時間PBMCs中ROS水平比較 低、中、高濃度吸氧組各時點PBMCs中ROS水平均高于對照組,中、高濃度吸氧組各時點ROS水平均高于低濃度吸氧組,高濃度吸氧組各時點ROS水平均高于中濃度吸氧組(P均<0.05);在各時點,PBMCs中ROS水平隨氧濃度增高而增高(F= 889.085,P<0.01)。生后7、14、28 d時各吸氧組PBMCs中ROS水平均高于生后0 d,生后14、28 d時各吸氧組PBMCs中ROS水平均高于生后7 d,生后28 d各吸氧組PBMCs中ROS水平均高于生后14 d(P均<0.05);各吸氧組PBMCs中ROS水平隨氧療時間增加而增高(F=368.535,P<0.01)。吸氧濃度和時點有交互效應(F=177.832,P<0.01),隨著時間的變化,不同氧濃度組各時點PBMCs中ROS的生成量變化趨勢不同。隨著吸入氧濃度的程度增加、生后時間的延長,ROS水平增加。見表1。
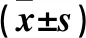
表1 各組不同時間PBMCs中ROS水平比較
注:與對照組比較,*P<0.05;與低濃度吸氧組比較,△P<0.05;與中濃度吸氧組比較,#P<0.05;與同組生后0 d比較,aP<0.05;與同組生后7 d比較,bP<0.05;與同組生后14 d比較,cP<0.05。
2.2 各組PBMCs中SIRT1定位 低、中、高濃度吸氧組各時點PBMCs中SIRT1轉位率均高于對照組,中、高濃度吸氧組各時點SIRT1轉位率均高于低濃度吸氧組,高濃度吸氧組各時點SIRT1轉位率均高于中濃度吸氧組(P均<0.05)。生后7 d、生后14 d、生后28 d各吸氧組PBMCs中SIRT1轉位率均高于生后0 d,生后14 d、生后28 d各吸氧組PBMCs中SIRT1轉位率均高于生后7 d,生后28 d各吸氧組PBMCs中SIRT1轉位率均高于生后14 d(P均<0.05)。吸氧濃度和時點有交互效應(F=347.299,P<0.01),隨著吸入氧濃度的濃度增加及生后時間的延長,SIRT1轉位率增加。見表2。
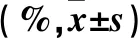
表2 各組PBMCs中SIRT1轉位率比較
注:與對照組比較,*P<0.05;與低濃度吸氧組比較,△P<0.05;與中濃度吸氧組比較,#P<0.05;與同組生后0 d比較,aP<0.05;與同組生后7 d比較,bP<0.05;與同組生后14 d比較,cP<0.05。
2.3 各組PBMCs中胞核SIRT1蛋白表達比較 低、中、高濃度吸氧組除生后0 d外,其余時點上PBMCs中胞核SIRT1蛋白表達均低于對照組(P均<0.05),中、高濃度吸氧組除生后0 d外,其余時點PBMCs中胞核SIRT1蛋白表達均低于低濃度吸氧組(P均<0.05),高濃度吸氧組生后7 d、生后14 d、生后28 d的PBMCs中胞核SIRT1蛋白表達均低于中濃度吸氧組(P均<0.05)。與生后0 d比較,生后7 d各吸氧組的PBMCs中胞核SIRT1蛋白表達均無明顯改變(P均>0.05),生后14、28 d各吸氧組PBMCs中胞核SIRT1蛋白表達均低于生后7 d(P均<0.05),生后28 d的各吸氧組PBMCs中胞核SIRT1蛋白表達均低于生后14 d(P均<0.05)。吸氧濃度和時點有交互效應(F=3.117,P<0.05),隨著吸入氧濃度的濃度增加及生后時間的延長,SIRT1轉位率增加。見表3。
表3 各組PBMCs中胞核SIRT1蛋白表達比較
注:與對照組比較,*P<0.05;與低濃度吸氧組比較,△P<0.05;與中濃度吸氧組比較,#P<0.05;與同組生后0 d比較,aP<0.05;與同組生后7 d比較,bP<0.05;與同組生后14 d比較,cP<0.05。
2.4 各組PBMCs中SENP1蛋白表達比較 中、高濃度吸氧組除生后0 d外,其余時點PBMCs中SENP1蛋白表達均高于低濃度吸氧組(P均<0.05),高濃度吸氧組生后7、14、28 d的PBMCs中SENP1蛋白表達均高于中濃度吸氧組(P均<0.05)。中、高濃度吸氧組生后14、28 d的PBMCs中SENP1蛋白表達均高于生后7 d(P均<0.05),中、高濃度吸氧組生后28 d的PBMCs中SENP1蛋白表達均高于生后14 d(P均<0.05)。吸氧濃度和時點有交互效應(F=7.400,P<0.05),隨著吸入氧濃度的增加及生后時間的延長,SENP1蛋白表達增加。見表4。
表4 各組PBMCs中SENP1蛋白表達比較
注:與對照組比較,*P<0.05;與低濃度吸氧組比較,△P<0.05;與中濃度吸氧組比較,#P<0.05;與同組生后0 d比較,aP<0.05;與同組生后7 d比較,bP<0.05;與同組生后14 d比較,cP<0.05。
2.5 各組PBMCs中乙酰化p53蛋白表達比較 中、高濃度吸氧組各時點PBMCs中乙酰化p53蛋白表達均高于低濃度吸氧組(P均<0.05),高濃度吸氧組各時點PBMCs中乙酰化p53蛋白表達均高于中濃度吸氧組(P均<0.05)。生后7、14、28 d的中、高濃度吸氧組的PBMCs中乙酰化p53蛋白表達均高于生后0 d(P均<0.05),生后14、28 d的中、高濃度吸氧組PBMCs中乙酰化p53蛋白表達均高于生后7 d(P均<0.05),生后28 d的中、高濃度吸氧組PBMCs中乙酰化p53蛋白表達均高于生后14 d(P均<0.05)。吸氧濃度和時點有交互效應(F=8.6037,P<0.05),隨著吸入氧濃度的濃度增加及生后時間的延長,乙酰化p53蛋白增加。見表5。
表5 各組PBMCs中乙酰化p53蛋白表達比較
注:與對照組比較,*P<0.05;與低濃度吸氧組比較,△P<0.05;與中濃度吸氧組比較,#P<0.05;與同組生后0 d比較,aP<0.05;與同組生后7 d比較,bP<0.05;與同組生后14 d比較,cP<0.05。
3 討論
近年來,隨著產前糖皮質激素的應用、生后呼吸支持技術的發展、新生兒重癥監護病房的建立等,極早早產兒、極低體質量兒存活率明顯提高[5]。但由于早產低體質量兒肺處于發育不成熟的階段,肺表面活性物質分泌不足、肺順應性低,故生后早期就有氧療指征,常需行機械通氣及氧療,置于長時間用氧環境中[6]。但氧療在提高存活率的同時,由于早產兒各系統發育不成熟,高濃度氧療可導致氧損傷或氧中毒,其最常見的并發癥是肺損傷。發生氧中毒時,可直接損傷肺泡上皮細胞、毛細血管內皮細胞,使肺泡毛細血管通透性增加,最終可誘發早產兒BPD的產生[7]。目前,對長時間及高濃度氧療導致的氧損傷機制尚不明確。
研究發現,氧化應激反應在高氧肺損傷中起著重要作用,氧化應激時線粒體的ROS生成增加,當累積超過正常生理水平時,會損傷線粒體內膜及DNA,影響線粒體的生物合成,破壞氧化呼吸鏈NAD+與NADH的相互轉換[8]。研究證明,當高氧暴露后,由于這類早產兒肺發育極不成熟,缺乏抵抗氧化應激的能力,高氧誘導的大量ROS不能被機體抗氧化應激酶所清除,造成氧化應激損傷,甚至這些ROS會上調促凋亡蛋白的表達,激活Casepse凋亡途徑,導致高氧肺損傷[9]。本研究發現,吸氧濃度和時點有交互效應,說明隨著時間的變化,不同氧濃度組各時點PBMCs中ROS的生成量變化趨勢不同,故PBMCs中ROS水平隨氧濃度的增高而增高,各吸氧組PBMCs中ROS水平隨氧療時間的增加而增高。
SIRT1是一種依賴酰胺腺嘌呤二核苷酸NAD+的組蛋白/非組蛋白去乙酰化酶,正常生理情況下,SIRT1可維持足夠的NAD+,對活性氧簇的產生具有抑制作用。SIRT1在哺乳動物細胞質和細胞核中均有表達,尤以細胞核為主,細胞核較細胞質明顯多,并且核內SIRT1亞細胞定位受Ⅱ類去乙酰化酶核-漿穿梭中機制調節,與細胞增殖、分化、衰老、凋亡和代謝密切相關。Yang等[10]研究發現,人類SIRT1蛋白在生理調節下定位于細胞核。有研究顯示[11],在細胞核中SIRT1表達是抗凋亡作用,而在細胞質內是促凋亡作用,所以核內SIRT1在機體損傷后升高顯得更加有保護意義。隨著ROS升高,會使生物膜脂質氧化,膜的正常結構發生破壞后,膜的通透性和流動性發生改變,導致SIRT1發生核質穿梭,細胞核中SIRT1減少,其在胞核中發揮的去乙酰化作用減弱,對細胞的保護性作用減弱,并且轉移到細胞質的SIRT1是促凋亡因子。本課題組前期研究發現,高氧可誘導ROS產生增加,使SIRT1發生核質穿梭[2]。本研究發現,在免疫熒光檢測中,SIRT1的轉位率,在吸氧濃度和時點有交互效應,提示隨著吸氧濃度的增加、生后時間延長SIRT1發生核-質穿梭的細胞數也增加,會出現去乙酰化作用減弱,與SIRT1結合的組蛋白,發生乙酰化反應,會啟動參與細胞損傷的過程。并且也發現,SIRT1蛋白在細胞核的表達在在吸氧濃度和時點有交互效應,說明隨著吸氧濃度增加、生后時間的延長, ROS增加,細胞氧化應激程度加重,脂質氧化后通透性增加,各吸氧組各時點PBMCs中SIRT1蛋白發生轉位后,在細胞核表達減少,而細胞質中表達逐漸增多,細胞損傷進行性加重。
SENPs在哺乳動物細胞中有6個成員,目前研究較多的是SENP1。SIRT1是SUMO的底物蛋白,能被SUMO化修飾,形成SIRT1/SUMO蛋白復合物,參與應對抗凋亡等保護作用[12]。研究證實[3,13],當細胞受到氧化應激等刺激時,SENP1不僅參與SUMO化可逆過程,還參與催熟SUMO-1前體,參與SUMO化反應,一方面,SUMO化修飾SIRT1是SUMO-1,SENP1參與該動態反應的過程,另一方面,SIRT1又可以被SENP1去SUMO化,因此,SIRT1的去乙酰化活性會受到抑制。本研究結果顯示,吸氧濃度和時點有交互效應,說明隨著生后時間的變化,不同吸氧濃度組各時點PBMCs中SENP1蛋白表達不同。隨著吸入氧濃度的濃度增加及生后時間的延長,SENP1蛋白表達增加,提示隨著SENP1的升高,發生去SUMO化和SUMO化反應的失衡,SIRT1/SUMO蛋白復合物解體,下調SIRT1的去乙酰化作用,SIRT1也將發生核質穿梭。
SIRT1可以通過去乙酰化作用于靶蛋白如p53、FOXO家族等,發揮其抗氧化應激、抗炎、抗凋亡等保護細胞的作用[3]。研究表明,SIRT1多作用于p53的第373和382賴氨酸位點,SIRT1可能使p53在第373和382賴氨酸位點去乙酰化,進而通過抑制p53的轉錄活性,從而阻止氧化應激誘導的細胞凋亡。乙酰化p53的促凋亡作用可以被SIRT1的去乙酰化作用緩解,達到抑制凋亡的作用[10]。本研究結果發現,與對照組相比,在低濃度吸氧組生后7 d時,細胞乙酰化p53的表達并無差異,這可能是機體p53蛋白保護作用并未失衡;此外,吸氧濃度和時點有交互效應,說明隨著生后時間的變化,不同吸氧濃度組各時點PBMCs中乙酰化p53蛋白表達不同;隨著吸入氧濃度的濃度增加及生后時間的延長,乙酰化p53蛋白增加,表明氧化應激過度時,SENP1改變,胞核SIRT1發生轉位后,SIRT1去乙酰化作用受到抑制,相應地,乙酰化p53升高。
綜上所述,隨著氧暴露的程度增加及生后時間的延長,ROS表達超過機體清除能力,發生氧化應激,一方面導致脂質膜通透性發生改變,另一方面使SENP1參與的SUMO化動態可逆過程受到抑制,SIRT1核質穿梭更加明顯,胞核SIRT1去乙酰化降低,損傷因子乙酰化p53表達增加,最終導致氧化應激損傷。此外,SUMO化SIRT1的改變是否受到氧暴露的時間影響,機制尚不清楚,是否還受到其他位點蛋白的參與調控等均待進一步研究證明。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
