內(nèi)側(cè)杏仁核中催產(chǎn)素對(duì)布氏田鼠社會(huì)行為的影響
2017-04-08 07:18:52張楠閆立新高杰孫秀萍宋銘晶
四川動(dòng)物 2017年2期
關(guān)鍵詞:實(shí)驗(yàn)
張楠, 閆立新, 高杰, 孫秀萍, 宋銘晶
(中國(guó)醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所,北京協(xié)和醫(yī)學(xué)院比較醫(yī)學(xué)中心,衛(wèi)生部人類(lèi)疾病比較醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室,北京100021)
內(nèi)側(cè)杏仁核中催產(chǎn)素對(duì)布氏田鼠社會(huì)行為的影響
張楠, 閆立新, 高杰, 孫秀萍, 宋銘晶*
(中國(guó)醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所,北京協(xié)和醫(yī)學(xué)院比較醫(yī)學(xué)中心,衛(wèi)生部人類(lèi)疾病比較醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室,北京100021)
目的 探究?jī)?nèi)側(cè)杏仁核(MeA)腦區(qū)中催產(chǎn)素(OT)對(duì)布氏田鼠Lasiopodomysbrandtii社會(huì)行為的影響。方法 在雄性布氏田鼠MeA區(qū)注射催產(chǎn)素受體拮抗劑后,采用社會(huì)識(shí)別實(shí)驗(yàn)、中央競(jìng)技場(chǎng)實(shí)驗(yàn)確定MeA區(qū)催產(chǎn)素受體的變化對(duì)其社會(huì)行為的影響。結(jié)果 社會(huì)識(shí)別實(shí)驗(yàn)發(fā)現(xiàn)在MeA區(qū)注射人工腦脊液的布氏田鼠(對(duì)照組)對(duì)陌生鼠的探索時(shí)間顯著長(zhǎng)于探索熟悉鼠(0.001
布氏田鼠;催產(chǎn)素;杏仁核;社會(huì)記憶;親社會(huì)性
無(wú)論是人還是動(dòng)物都有一部分個(gè)體或物種表現(xiàn)出較多的社會(huì)友善行為,一部分表現(xiàn)出較多的反社會(huì)行為。嚙齒動(dòng)物既有以家族為單位群居的,表現(xiàn)出較多社會(huì)互助行為,如社會(huì)性強(qiáng)的橙腹田鼠Microtusochrogaster和布氏田鼠Lasiopodomysbrandtii等(Olazabal & Young,2006;Wangetal.,2011),也有獨(dú)居的,在繁殖期和非繁殖期都有很強(qiáng)攻擊性的山地田鼠Microtusmontanus和大倉(cāng)鼠Tscherskiatriton(Liuetal.,2001;Panetal.,2010)。Pan等(2016)研究發(fā)現(xiàn)群居的橙腹田鼠在遇到種群內(nèi)其他個(gè)體時(shí)表現(xiàn)出更多的探究行為,即社會(huì)探究行為,而獨(dú)居的山地田鼠表現(xiàn)出對(duì)空籠的探究行為超過(guò)對(duì)其他個(gè)體的探究行為。鑒于不同嚙齒動(dòng)物社會(huì)行為的巨大差異,通過(guò)對(duì)其特定腦區(qū)的基因進(jìn)行調(diào)控,改變其社會(huì)行為,可以研究該動(dòng)物社會(huì)行為的遺傳機(jī)制。
催產(chǎn)素(oxytocin,OT)是哺乳動(dòng)物分泌的一種環(huán)狀九肽分子神經(jīng)激素,以其在分娩和泌乳中起神經(jīng)調(diào)節(jié)作用而聞名,研究發(fā)現(xiàn)其在調(diào)節(jié)社會(huì)認(rèn)知行為方面的影響顯著,其中包括焦慮、學(xué)習(xí)記憶、社會(huì)認(rèn)知、配偶選擇等行為(孟雅婧等,2014;Shen,2015)。OT及其受體在動(dòng)物腦區(qū)內(nèi)廣泛表達(dá),不過(guò)目前的研究顯示其在不同腦區(qū)的表達(dá)會(huì)影響不同的社會(huì)行為,已有研究通過(guò)對(duì)棕色田鼠特定腦區(qū)內(nèi)側(cè)杏仁核(medial amygdala,MeA)和伏隔核區(qū)OT的表達(dá)進(jìn)行調(diào)控,發(fā)現(xiàn)雄性棕色田鼠的育幼行為會(huì)隨著OT的表達(dá)減少而減弱(Caoetal.,2014)。Wang等(2015)對(duì)不同生殖經(jīng)驗(yàn)的雄性棕色田鼠MeA腦區(qū)OT的研究發(fā)現(xiàn)OT及催產(chǎn)素受體(oxytocin receptor,OTR)的表達(dá)與棕色田鼠的生殖經(jīng)驗(yàn)和親子行為呈正相關(guān)。
布氏田鼠社會(huì)行為發(fā)達(dá),是一種研究動(dòng)物親社會(huì)行為較為理想的模型動(dòng)物,已有研究對(duì)群居的布氏田鼠與獨(dú)居的大倉(cāng)鼠腦區(qū)OT的分布進(jìn)行檢測(cè),發(fā)現(xiàn)在MeA等腦區(qū),群居的布氏田鼠OT的分布明顯多于獨(dú)居的大倉(cāng)鼠(Xuetal.,2010),而本實(shí)驗(yàn)室對(duì)布氏田鼠與BALB/c小鼠不同腦區(qū)的OT及其受體的Western Blot檢測(cè)顯示布氏田鼠MeA區(qū)OT分布較多,且有研究表明腦部MeA區(qū)與社會(huì)識(shí)別行為有密切聯(lián)系(Wangetal.,2013)。社會(huì)識(shí)別行為以社會(huì)記憶能力為生理基礎(chǔ),個(gè)體的社會(huì)記憶是個(gè)體間進(jìn)行社會(huì)交往所必需的,是某個(gè)體在同物種內(nèi)能夠區(qū)分熟悉個(gè)體與陌生個(gè)體的能力,是種群內(nèi)所有社會(huì)關(guān)系形成的基礎(chǔ)(Fergusonetal.,2002)。為了明確大腦MeA區(qū)OT表達(dá)對(duì)動(dòng)物社會(huì)行為和社會(huì)性的影響,本研究調(diào)控布氏田鼠MeA區(qū)OT的表達(dá),并進(jìn)行行為學(xué)分析,揭示動(dòng)物社會(huì)行為的遺傳調(diào)控機(jī)制。
1 材料與實(shí)驗(yàn)方法
1.1 材料
1.1.1 實(shí)驗(yàn)動(dòng)物 4月齡雄性布氏田鼠60只,體質(zhì)量45~55 g,均為中國(guó)醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所飼養(yǎng)、繁育并凈化。10只雄性3月齡SPF級(jí)BALB/c小鼠,體質(zhì)量約25 g,購(gòu)自北京維通利華實(shí)驗(yàn)動(dòng)物技術(shù)有限公司,實(shí)驗(yàn)動(dòng)物生產(chǎn)許可證號(hào):SYXK(京)2012-0022。動(dòng)物飼養(yǎng)在獨(dú)立送排風(fēng)凈化動(dòng)物籠。飲用水、墊料和籠具均經(jīng)高壓滅菌處理,飼料經(jīng)60Co照射消毒達(dá)到SPF級(jí)動(dòng)物飼料標(biāo)準(zhǔn),光照時(shí)間14L∶10D。實(shí)驗(yàn)動(dòng)物的使用通過(guò)了中國(guó)醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所的實(shí)驗(yàn)動(dòng)物使用和管理委員會(huì)批準(zhǔn),批準(zhǔn)編號(hào):ILAS-PG-2014-011。
1.1.2 儀器 腦立體定位儀,曠場(chǎng)裝置(長(zhǎng)50 cm×寬50 cm×高30 cm),三箱裝置(長(zhǎng)105 cm×寬35 cm×高35 cm),動(dòng)物行為學(xué)視頻跟蹤系統(tǒng)(Noldus Information Technology,EthoVision XT,The Netherlandtiaokongs)。
1.2 實(shí)驗(yàn)方法
1.2.1 Western Blot檢測(cè) 雄性布氏田鼠(n=6)和雄性BALB/c小鼠(n=6)取腦,扣取MeA區(qū)腦組織,由于單只MeA區(qū)腦組織較少,故將扣取的布氏田鼠和小鼠腦組織分別混合,提取蛋白,Western Blot檢測(cè)其OT與OTR的本底表達(dá)量,對(duì)比布氏田鼠MeA腦區(qū)OT與OTR的表達(dá)量及其與小鼠的差異。
1.2.2 埋管注射 布氏田鼠(n=46)經(jīng)水合氯醛麻醉(5%,0.6 mL/100 g)后固定在腦立體定位儀上,確認(rèn)腦部前囟為零點(diǎn)坐標(biāo)(X,Y,Z),根據(jù)MeA區(qū)三維坐標(biāo)(X:-1.8 mm,Y:±2.3 mm,Z:-6.3 mm)開(kāi)顱鉆打孔,雙側(cè)腦部埋管,導(dǎo)管深入腦下6.3 mm,進(jìn)入MeA腦區(qū)。套管通過(guò)螺絲和牙科水泥固定于頭骨上,不進(jìn)行注射時(shí),用導(dǎo)管帽進(jìn)行封閉。待傷口愈合,手術(shù)10 d后微量進(jìn)樣器給藥,然后進(jìn)行行為學(xué)實(shí)驗(yàn)。注射試劑:實(shí)驗(yàn)組(n=23)雙側(cè)注射催產(chǎn)素受體拮抗劑(oxytocin receptor antagonist,OTRA;Sigma,L-368899),注射劑量為單側(cè)750 ng(Arakawaetal.,2010),注射速度0.1 μL·min-1,總體積0.5 μL。注射后留針2 min,使藥物充分?jǐn)U散,防止倒流;對(duì)照組(n=23)注射相同體積的人工腦脊液(artificial cerebrospinal fluid,ACSF)。
1.2.3 社會(huì)識(shí)別實(shí)驗(yàn) 三箱測(cè)試是在社會(huì)識(shí)別或社會(huì)新奇實(shí)驗(yàn)中用來(lái)評(píng)估個(gè)體社交能力和對(duì)新事物的興趣程度的方法。實(shí)驗(yàn)過(guò)程參考Page等(2009)的實(shí)驗(yàn)步驟并加以修改,測(cè)試區(qū)域由3個(gè)相鄰的箱體組成(長(zhǎng)105 cm×寬35 cm×高35 cm),中間2個(gè)透明塑料分隔器分隔,并通過(guò)開(kāi)放的門(mén)道使三箱連通起來(lái)。實(shí)驗(yàn)對(duì)象為實(shí)驗(yàn)組布氏田鼠(n=12)和對(duì)照組布氏田鼠(n=12)。實(shí)驗(yàn)分為3個(gè)階段:
(1)第一階段(適應(yīng)階段):社會(huì)交互箱兩側(cè)箱體空置,中間的2個(gè)隔板門(mén)關(guān)閉,將實(shí)驗(yàn)鼠放入中間箱體,適應(yīng)10 min。
(2)第二階段:適應(yīng)結(jié)束后,同一時(shí)間段進(jìn)行測(cè)試期。測(cè)試過(guò)程中,左側(cè)箱體放入空籠,右側(cè)箱體的籠中放入同性幼年刺激鼠。將實(shí)驗(yàn)鼠迅速放入中間箱體中,撤掉隔板,立即開(kāi)始測(cè)試,動(dòng)物跟蹤系統(tǒng)自動(dòng)記錄實(shí)驗(yàn)鼠進(jìn)入各箱的時(shí)間、次數(shù)、路程,接觸空鼠籠和刺激鼠的潛伏期、時(shí)間,跟蹤時(shí)間為10 min。10 min后將實(shí)驗(yàn)鼠取出。
(3)第三階段:步驟(2)中所用刺激鼠作為熟悉鼠,同時(shí),在左側(cè)箱體的籠中,放入另外一只陌生刺激鼠,觀察10 min,采集數(shù)據(jù)指標(biāo)同步驟(2)。每只實(shí)驗(yàn)鼠結(jié)束實(shí)驗(yàn)后都需要將實(shí)驗(yàn)鼠的糞便清除干凈,并用75%的酒精再次擦拭,掩蓋老鼠的味道,待酒精揮發(fā)完后再進(jìn)行下一只實(shí)驗(yàn)。
1.2.4 中央競(jìng)技場(chǎng)實(shí)驗(yàn) 中央競(jìng)技場(chǎng)實(shí)驗(yàn)參照李廣宇(2012)的方法,在曠場(chǎng)箱中間設(shè)置一個(gè)能上下抽拉的隔板,將曠場(chǎng)箱分為左右相等的兩部分。實(shí)驗(yàn)于暗光周期1 h后開(kāi)始,行為過(guò)程采用攝像記錄設(shè)備(夜視模式)記錄后進(jìn)行分析,實(shí)驗(yàn)時(shí)將實(shí)驗(yàn)鼠與刺激鼠分別置于中央競(jìng)技場(chǎng)隔板的兩側(cè),并給予其5 min適應(yīng)環(huán)境,每組實(shí)驗(yàn)時(shí)間為20 min,每次實(shí)驗(yàn)完成后清理箱底,用75%酒精擦拭箱底和箱壁,待晾干之后再進(jìn)行下一組實(shí)驗(yàn)。實(shí)驗(yàn)對(duì)象為實(shí)驗(yàn)組布氏田鼠(n=11)和對(duì)照組布氏田鼠(n=11)。
1.3 數(shù)據(jù)分析
行為學(xué)數(shù)據(jù)采用SPSS 20.0進(jìn)行統(tǒng)計(jì)學(xué)處理,用獨(dú)立樣本t檢驗(yàn)分析MeA區(qū)OT對(duì)社會(huì)識(shí)別實(shí)驗(yàn)和中央競(jìng)技場(chǎng)實(shí)驗(yàn)數(shù)據(jù)的影響。P<0.05為差異有統(tǒng)計(jì)學(xué)意義,P<0.01為差異有高度統(tǒng)計(jì)學(xué)意義,數(shù)據(jù)以平均值±標(biāo)準(zhǔn)誤(Mean±SE)表示,得到的結(jié)果用GraphPad Prism 6作圖。
2 實(shí)驗(yàn)結(jié)果
2.1 Western Blot檢測(cè)
Western Blot檢測(cè)顯示在雄性布氏田鼠MeA區(qū)OT的表達(dá)量遠(yuǎn)高于雄性BALB/c小鼠(圖1:A),雄性布氏田鼠MeA區(qū)OTR的表達(dá)量與雄性BALB/c小鼠沒(méi)有明顯差異(圖1:B)。

圖1 Western Blot檢測(cè)OT與OTR的蛋白表達(dá)
A. 雄性布氏田鼠與雄性BALB/c小鼠MeA區(qū)OT蛋白相對(duì)表達(dá)量的對(duì)比, B. 雄性布氏田鼠與雄性BALB/c小鼠MeA區(qū)OTR蛋白相對(duì)表達(dá)量的對(duì)比, 用GAPDH蛋白做對(duì)照, 比較OT或OTR與GAPDH灰度的比值;***P<0.001。
A. comparison of OT protein relative expression in the MeA between the maleLasiopodomysbrandtiiand the male BALB/c mice, B. comparison of OTR protein relative expression in the MeA between the maleL.brandtiiand the male BALB/c mice using GAPDH as control;***P<0.001.
2.2 社會(huì)識(shí)別實(shí)驗(yàn)
由于腦部埋管注射位置有偏差,通過(guò)腦部切片確認(rèn)位置,統(tǒng)計(jì)分析注射正確位置樣本的行為:實(shí)驗(yàn)組,n=8;對(duì)照組,n=7。對(duì)照組和實(shí)驗(yàn)組布氏田鼠對(duì)有刺激鼠的籠探索時(shí)間多于對(duì)空籠的探索時(shí)間,且差異有高度統(tǒng)計(jì)學(xué)意義(0.001
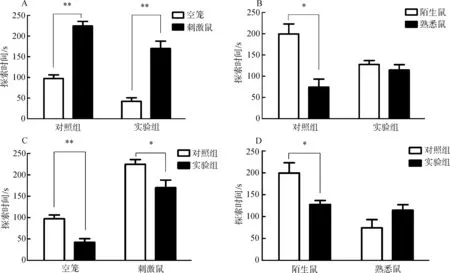
圖2 實(shí)驗(yàn)組和對(duì)照組社會(huì)識(shí)別實(shí)驗(yàn)
Fig. 2 Social recognition experiment of experimental group and control group
A.對(duì)空籠和刺激鼠探索時(shí)間, B. 對(duì)熟悉鼠和陌生鼠探索時(shí)間, C. 對(duì)空籠和刺激鼠探索時(shí)間的組間對(duì)比, D. 對(duì)陌生鼠和熟悉鼠探索時(shí)間的組間對(duì)比;*P<0.05,**P<0.01。
A.the exploration time between empty and stimulus, B. the exploration time between novel and familiar, C. the exploration time of empty and stimulus between the groups, D. comparison the exploration time of novel and familiar between the groups;*P<0.05,**P<0.01.
2.3 中央競(jìng)技場(chǎng)實(shí)驗(yàn)
由于腦部埋管注射位置有偏差,通過(guò)腦部切片確認(rèn)位置,統(tǒng)計(jì)分析注射正確位置樣本的行為:實(shí)驗(yàn)組,n=7;對(duì)照組,n=7。實(shí)驗(yàn)組布氏田鼠的攻擊潛伏期和攻擊時(shí)長(zhǎng)與對(duì)照組的差異均無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)(圖3)。
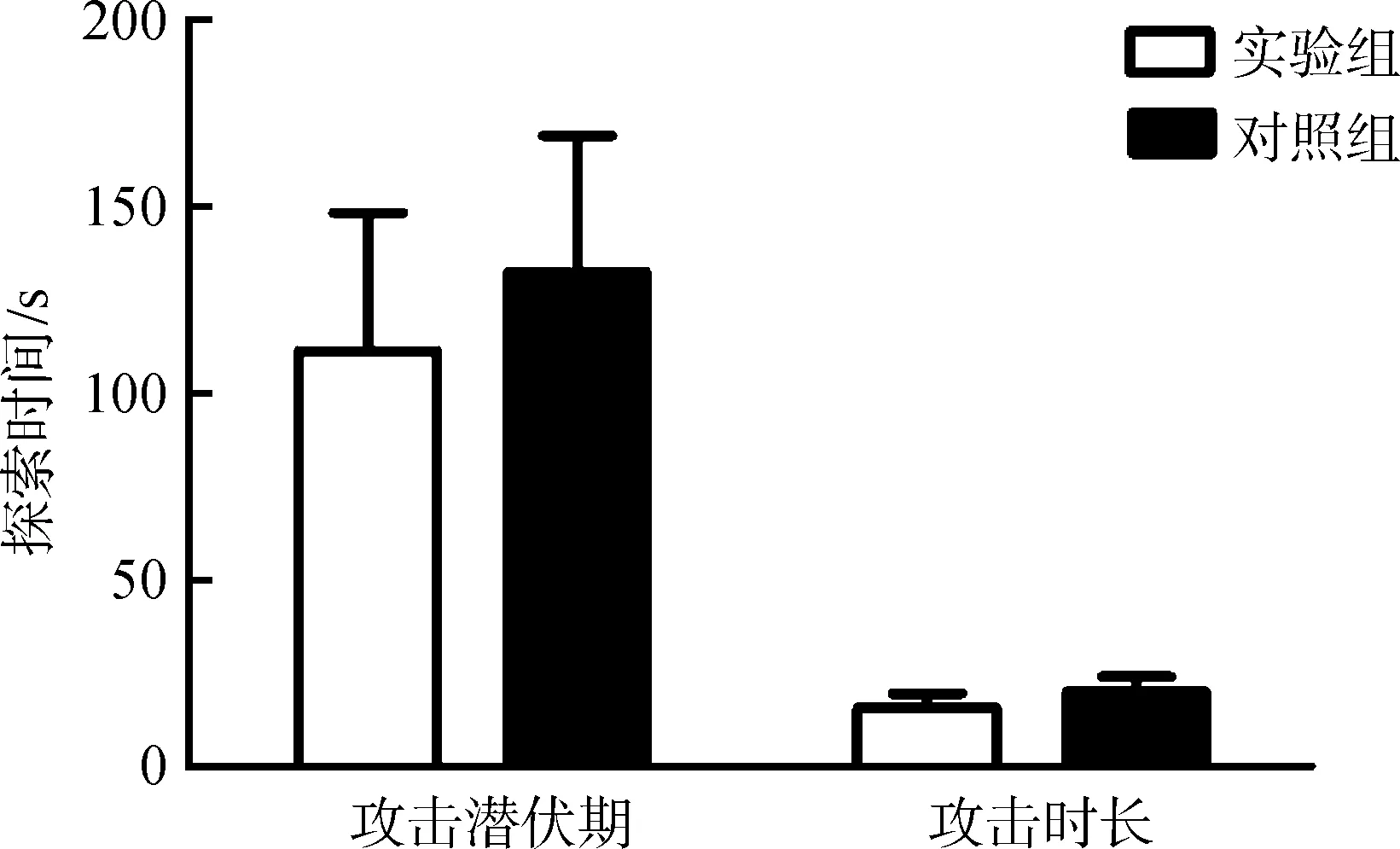
圖3 催產(chǎn)素受體抑制劑對(duì)布氏田鼠攻擊行為的影響
3 討論
識(shí)別一個(gè)個(gè)體并對(duì)此個(gè)體形成一個(gè)社會(huì)記憶的過(guò)程是后期社會(huì)關(guān)系形成的一個(gè)主要步驟。社會(huì)識(shí)別實(shí)驗(yàn)是基于嚙齒類(lèi)動(dòng)物探索新物體(或者個(gè)體)的時(shí)間要多于探索熟悉物體(個(gè)體)的時(shí)間這種現(xiàn)象而成立的(Winslow,2003)。MeA區(qū)注射ACSF的對(duì)照組布氏田鼠對(duì)陌生鼠的探索時(shí)間明顯高于對(duì)熟悉鼠的探索時(shí)間,而注射OTRA的實(shí)驗(yàn)組布氏田鼠對(duì)陌生鼠和熟悉鼠的探索時(shí)間的差異無(wú)統(tǒng)計(jì)學(xué)意義,說(shuō)明MeA區(qū)內(nèi)OTR的抑制能影響布氏田鼠的社會(huì)行為。這種社會(huì)行為的改變可能有2個(gè)原因:一是由于社會(huì)記憶形成的障礙造成的;二是動(dòng)物親社會(huì)性的改變?cè)斐傻摹an等(2016)對(duì)獨(dú)居的山地田鼠和群居的橙腹田鼠分別做社會(huì)識(shí)別實(shí)驗(yàn),發(fā)現(xiàn)獨(dú)居的山地田鼠對(duì)空籠探索的時(shí)間比對(duì)同種的其他個(gè)體更長(zhǎng),而且對(duì)熟悉個(gè)體和陌生個(gè)體的探索時(shí)間沒(méi)有顯著差異,推測(cè)因?yàn)楠?dú)居動(dòng)物的親社會(huì)性水平低,所以它們更關(guān)注物體,而不是同種的陌生或熟悉個(gè)體,缺少探究行為。而野外實(shí)驗(yàn)研究也報(bào)道獨(dú)居鼠在自然狀態(tài)下遭遇時(shí)多回避或攻擊(張健旭等,1999)。本實(shí)驗(yàn)結(jié)果顯示,實(shí)驗(yàn)組和對(duì)照組雄性布氏田鼠在面對(duì)空籠和刺激鼠時(shí),對(duì)刺激鼠的探索時(shí)間都顯著高于對(duì)空籠的探索時(shí)間,初步說(shuō)明MeA區(qū)OT阻斷不影響雄性布氏田鼠的親社會(huì)性。而對(duì)不同組間進(jìn)行比較發(fā)現(xiàn),注射OTRA的實(shí)驗(yàn)組布氏田鼠對(duì)空籠、刺激鼠以及陌生鼠的探索時(shí)間都明顯低于注射ACSF的對(duì)照組布氏田鼠,說(shuō)明OTRA能夠降低布氏田鼠的社會(huì)探索行為。以上實(shí)驗(yàn)都證明在MeA區(qū)注射OTRA能夠減少其社會(huì)探索行為。但是這種社會(huì)行為的改變是由于社會(huì)記憶障礙造成的還是親社會(huì)性的改變?cè)斐傻倪€需要進(jìn)一步確定。因此為了進(jìn)一步確定布氏田鼠的親社會(huì)性有無(wú)改變,我們用中央競(jìng)技場(chǎng)的實(shí)驗(yàn)進(jìn)行驗(yàn)證。結(jié)果證明,實(shí)驗(yàn)組與對(duì)照組相比,其攻擊潛伏期和攻擊時(shí)長(zhǎng)都沒(méi)有明顯差異,表明OT在MeA區(qū)的阻斷對(duì)布氏田鼠的主動(dòng)攻擊行為沒(méi)有影響。綜上所述,OT在MeA的表達(dá)是通過(guò)影響布氏田鼠的社會(huì)記憶,進(jìn)而影響其社會(huì)行為,而不是改變布氏田鼠的親社會(huì)性。
已有研究證明OT在不同腦區(qū)表達(dá)變化與自閉癥、抑郁癥等精神類(lèi)疾病的形成有關(guān)(Penagarikanoetal.,2015)。Shen(2015)對(duì)近年OT的研究進(jìn)行總結(jié),發(fā)現(xiàn)有關(guān)OT對(duì)自閉癥影響的研究中,OT的治愈能力不僅能在動(dòng)物實(shí)驗(yàn)中呈現(xiàn),在臨床治療中也得到了肯定,研究中還發(fā)現(xiàn)OT的劑量效應(yīng)呈“U”型曲線,推測(cè)OT對(duì)精神類(lèi)疾病的作用比較復(fù)雜,有待進(jìn)一步探究。現(xiàn)有有關(guān)OT動(dòng)物實(shí)驗(yàn)與臨床治療的研究表明,OT的作用隨其表達(dá)腦區(qū)的位置和濃度梯度變化均有所改變,所以我們希望找到調(diào)控社會(huì)行為的主要腦區(qū)和OT對(duì)社會(huì)行為的調(diào)控機(jī)制,本實(shí)驗(yàn)室和其他研究都已經(jīng)證明OT與社會(huì)行為有關(guān),但要確定影響社會(huì)行為的主要腦區(qū),還需要對(duì)布氏田鼠的不同腦區(qū)進(jìn)行比較研究和基因調(diào)控才能實(shí)現(xiàn)。
李廣宇. 2012. 配偶選擇對(duì)黑線倉(cāng)鼠腦內(nèi)Dadr2表達(dá)的影響[D]. 山東曲阜: 曲阜師范大學(xué).
孟雅婧, 楊洋, 劉瑩, 等. 2014. 催產(chǎn)素受體配體及其中樞和外周功能研究進(jìn)展[J]. 中國(guó)藥理學(xué)通報(bào), (6): 752-756.
張健旭, 張知彬, 王祖望. 1999. 大倉(cāng)鼠在繁殖期的行為關(guān)系及交配行為[J]. 獸類(lèi)學(xué)報(bào), (2): 52-55.
Arakawa H, Arakawa K, Deak T. 2010. Oxytocin and vasopressin in the medial amygdala differentially modulate approach and avoidance behavior toward illness-related social odor[J]. Neuroscience, 171(4): 1141-1151.
Cao Y, Wu R, Tai F,etal. 2014. Neonatal paternal deprivation impairs social recognition and alters levels of oxytocin and estrogen receptor alpha mRNA expression in the MeA and NAcc, and serum oxytocin in mandarin voles[J]. Hormones & Behavior, 65(1): 57-65.
Ferguson JE, Head BH, Frank FH,etal. 2002. Misoprostol versus low-dose oxytocin for cervical ripening: a prospective, randomized, double-masked trial[J]. American Journal of Obstetrics and Gynecology,187(2): 273-280.
Liu Y, Fowler CD, Wang Z. 2001. Ontogeny of brain-derived neurotrophic factor gene expression in the forebrain of prairie and montane voles[J]. Developmental Brain Research, 127(1): 51-61.
Olazabal DE, Young LJ. 2006. Oxytocin receptors in the nucleus accumbens facilitate “spontaneous” maternal behavior in adult female prairie voles[J]. Neuroscience, 141(2): 559-568.
Page DT, Kuti OJ, Sur M. 2009. Computerized aassessment of social approach behavior in mouse[J]. Frontiers in Behavioral Neuroscience,3: 48.
Pan Y, Liu Y, Lieberwirth C,etal. 2016. Species differences in behavior and cell proliferation/survival in the adult brains of female meadow and prairie voles[J]. Neuroscience, 315: 259-270.
Pan Y, Xu L, Young KA,etal. 2010. Agonistic encounters and brain activation in dominant and subordinate male greater long-tailed hamsters[J]. Hormones and Behavior, 58(3): 478-484.
Penagarikano O, Lazaro MT, Lu XH,etal. 2015. Exogenous and evoked oxytocin restores social behavior in the Cntnap2 mouse model of autism[J]. Science Translational Medicine, 7(271): 271r-278r.
Shen H. 2015. Neuroscience: the hard science of oxytocin[J]. Nature, 522(7557): 410-412.
Wang B, Li Y, Wu R,etal. 2015. Behavioral responses to pups in males with different reproductive experiences are sssociated with changes in central OT, TH and OTR, D1R, D2R mRNA expression in mandarin voles[J]. Hormones & Behavior, 67: 73-82.
Wang D, Li D, Liu M,etal. 2011. Behavioral evaluation of quinestrol as a sterilant in male Brandt’s voles[J]. Physiology & Behavior, 104(5): 1024-1030.
Wang Y, Xu L, Pan Y,etal. 2013. Species differences in the immunoreactive expression of oxytocin, vasopressin, tyrosine hydroxylase and estrogen receptor alpha in the brain of mongolian gerbils (Merionesunguiculatus) and Chinese striped hamsters (Cricetulusbarabensis)[J]. PLoS ONE, 8(6): e65807. DOI:10.1371/journal.pone.0065807.
Winslow JT. 2003. Mouse social recognition and preference[J]. Current Protocols in Neuroscience, 8: 8-16.
Xu L, Pan Y, Young KA,etal. 2010. Oxytocin and vasopressin immunoreactive staining in the brains of Brandt’s voles (Lasiopodomysbrandtii) and greater long-tailed hamsters (Tscherskiatriton)[J]. Neuroscience, 169(3): 1235-1247.
Effect of Oxytocin in the Medial Amygdala on the Social Behavior ofLasiopodomysbrandtii
ZHANG Nan, YAN Lixin, GAO Jie, SUN Xiuping, SONG Mingjing*
(Key Laboratory of Human Diseases Comparative Medicine, Ministry of Health, Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences & Comparative Medicine Center, Peking Union Medical College, Beijing 100021, China)
Objective To explore the influence of oxytocin (OT) in the medial amygdala (MeA) on the social behaviors of Brandt’s voles (Lasiopodomysbrandtii). Methods Oxytocin receptor antagonist (OTRA) was injected into the MeA of male Brants’s voles, and then the social behaviors were tested using social recognition experiment and neutral arena encounter experiment. Results The control group (injected with artificial cerebrospinal fluid) took more time to explore the strangers than the familiars (0.001
Lasiopodomysbrandtii; oxytocin; medial amygdala; social memory; prosocial
2016-10-21 接受日期:2016-12-15
國(guó)家自然科學(xué)基金項(xiàng)目(31301890); 農(nóng)業(yè)蟲(chóng)害鼠害綜合治理研究國(guó)家重點(diǎn)實(shí)驗(yàn)室開(kāi)放研究基金項(xiàng)目(Chinese IPM1615)
張楠(1991—), 男, 碩士研究生, 主要從事實(shí)驗(yàn)動(dòng)物學(xué)研究
*通信作者Corresponding author, E-mail:songmj@cnilas.org
10.11984/j.issn.1000-7083.20160288
Q959.837; Q95-33
A
1000-7083(2017)02-0193-05
猜你喜歡
作文·小學(xué)低年級(jí)(2025年2期)2025-02-13 00:00:00
小雪花·小學(xué)生快樂(lè)作文(2024年11期)2024-12-31 00:00:00
作文·小學(xué)低年級(jí)(2024年2期)2024-04-29 00:00:00
作文·小學(xué)低年級(jí)(2023年3期)2023-04-29 00:00:00
小獼猴智力畫(huà)刊(2022年9期)2022-11-04 02:31:42
小主人報(bào)(2022年4期)2022-08-09 08:52:06
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
