枯草芽孢桿菌精氨酸脫羧酶基因speA的表達(dá)與蛋白純化
2017-04-07 12:53:32曹寧王亞娟鐘杰佟碩秋吳擁軍
中國(guó)釀造 2017年3期
曹寧,王亞娟,鐘杰,佟碩秋,吳擁軍*
(貴州大學(xué)生命科學(xué)學(xué)院,貴州貴陽(yáng)550025)
枯草芽孢桿菌精氨酸脫羧酶基因speA的表達(dá)與蛋白純化
曹寧,王亞娟,鐘杰,佟碩秋,吳擁軍*
(貴州大學(xué)生命科學(xué)學(xué)院,貴州貴陽(yáng)550025)
根據(jù)枯草芽孢桿菌(Bacillus subtilis)BJ3-2的精氨酸脫羧酶(ADC)的編碼基因speA序列設(shè)計(jì)特異性酶切引物,克隆基因speA序列。測(cè)序結(jié)果顯示,基因speA全長(zhǎng)為1 473 bp,編碼490個(gè)氨基酸,分子質(zhì)量為58 ku。基因speA克隆至原核表達(dá)載體,獲得重組菌pET28a-speA/BL21,十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(SDS-PAGE)結(jié)果顯示,1.0 mmol/L的異丙基β-D-硫代半乳糖苷(IPTG)28℃誘導(dǎo)4 h,上清液和菌體均能表達(dá)出ADC蛋白,上清液經(jīng)純化、透析、冷凍干燥可獲得純度97%的ADC酶,酶聯(lián)免疫吸附檢測(cè)(ELISA)ADC酶活為16 780 U/mg。為speA基因的表達(dá)、純化及酶學(xué)性質(zhì)研究奠定了理論基礎(chǔ)。
枯草芽孢桿菌;精氨酸脫羧酶;表達(dá);純化
生物胺(biogenic amine,BA)是一類具有生物活性、低分子質(zhì)量、含氮有機(jī)化合物的總稱。根據(jù)組成成分可分為單胺(酪胺、組胺、尸胺、苯乙胺、色胺等)和多胺(腐胺、精胺和亞精胺)兩大類[1]。生物胺可作為生物體合成荷爾蒙、核苷酸、蛋白質(zhì)的前體物質(zhì),具有一定生理功能。但是過(guò)量攝入,則會(huì)引起諸如頭痛、惡心、心悸、血壓變化、呼吸紊亂等過(guò)敏反應(yīng),嚴(yán)重的還會(huì)危及生命[2]。
生物胺的產(chǎn)生需具備游離的氨基酸和氨基酸脫羧酶,通過(guò)氨基酸脫羧酶催化使氨基酸脫羧而產(chǎn)生[3]。目前研究發(fā)現(xiàn),氨基酸脫羧酶主要存在于芽孢桿菌屬、乳酸菌屬、埃希氏桿菌屬、變性菌屬、假單胞菌屬、沙門(mén)氏菌屬、志賀氏菌屬及一些檸檬酸細(xì)菌屬細(xì)菌,其中乳酸桿菌的催化脫羧作用最為突出[4]。
貴州地區(qū)水豆豉屬于典型的多菌混合細(xì)菌型自然發(fā)酵豆豉,主要參與發(fā)酵的微生物有芽孢桿菌屬、枝芽孢桿菌屬、產(chǎn)堿菌屬、腸球菌屬、叢毛單胞菌屬、乳酸菌屬和微球菌屬等,在這些微生物的發(fā)酵作用下形成風(fēng)味獨(dú)特的豆豉[5]。實(shí)驗(yàn)室前期研究發(fā)現(xiàn),細(xì)菌型豆豉能夠有效分離出8種生物胺,分別是色胺、β-苯乙胺、腐胺、尸胺、組胺、酪胺、亞精胺和精胺。其中前發(fā)酵階段亞精胺含量最高,精胺次之,后發(fā)酵階段酪胺含量最高,且顯著高于其他生物胺2倍以上[6]。在枯草芽孢桿菌中生物胺合成只有一條途徑,起始于精氨酸,在精氨酸脫羧酶、胍基丁胺酶、亞精胺合酶和精胺合酶共同作用合成亞精胺或精胺[7-8]。精氨酸脫羧酶(arginine decarboxylase,ADC)作為胺合成的關(guān)鍵酶[9-10]對(duì)精胺和亞精胺的形成起著重要的作用。所以目前認(rèn)為控制食品中過(guò)量生物胺最有前景的方法是通過(guò)控制精氨酸脫羧酶的活性來(lái)控制生物胺的形成[7]。
枯草芽孢桿菌(Bacillus subtilis)是中國(guó)農(nóng)業(yè)部批準(zhǔn)使用的一種重要工業(yè)酶制劑的生產(chǎn)菌,現(xiàn)已大量用于食品發(fā)酵行業(yè)[11]。本實(shí)驗(yàn)室從水豆豉樣品中分離獲得了B.subtilis BJ3-2菌株[12]。本研究對(duì)B.subtilisBJ3-2菌株中獲得的精氨酸脫羧酶(arginine decarboxylase ADC)基因speA序列進(jìn)行聚合酶鏈反應(yīng)(polymerase chain reaction,PCR)擴(kuò)增,通過(guò)
1 材料與方法
1.1 材料與試劑
1.1.1 菌種和質(zhì)粒
枯草芽孢桿菌(Bacillus subtilis)BJ3-2、大腸桿菌(Escherichiacoli)DH5α:本實(shí)驗(yàn)室保藏菌株;表達(dá)載體pET28a、宿主菌大腸桿菌(Escherichia coli)BL21(DE3):德國(guó)默克(中國(guó))公司。
1.1.2 培養(yǎng)基及試劑
LB培養(yǎng)基:配制方法參考文獻(xiàn)[13]。
T4 DNA連接酶、溶菌酶、基因組DNA提取試劑盒:美國(guó)Promega公司;脫氧核糖核苷三磷酸(deoxy-ribonucleotide triphosphate,dNTP)、DL2000 DNA Marker、rTaqDNA聚合酶、蛋白質(zhì)Marker、異丙基-β-D-硫代半乳糖苷(isopropyl-β-D-thiogalactopyranoside,IPTG)、限制性核酸內(nèi)切酶、DNA Ligation Kit Ver.2.1連接試劑盒:TaKaRa(大連)公司;氨芐青霉素(ampicillin,Amp)、卡拉霉素(kanamycin,Kan):北京索萊寶科技有限公司;E.Z.N.A.TMGelExtraction Kit、E.Z.N.A.TM Plasmid Mini KitⅠ:美國(guó)Omega公司;考馬斯亮藍(lán)R-250、精氨酸脫羧酶(ADC)酶聯(lián)免疫分析試劑盒:上海生工生物工程有限公司;His.Bind Purification Kit試劑盒:美國(guó)Novagen公司;咪唑(純度99%):上海展云化工有限公司。
1.2 儀器與設(shè)備
TGL-161高速臺(tái)式離心機(jī)、MyCycler PCR儀、Gel Doc XR凝膠成像系統(tǒng):美國(guó)Bio-Rad公司;Biosafer-10D真空冷凍干燥機(jī):美國(guó)Thermo公司。SW-CJ-1FD標(biāo)準(zhǔn)型凈化工作臺(tái):蘇州凈化設(shè)備有限公司;GMSX-280高壓滅菌鍋:英國(guó)阿斯太歐(Astell)公司;SHZ-82A氣浴恒溫振蕩器:江蘇榮華儀器制造有限公司;DYCZ-25D小型垂直電泳槽(SDA-PAGE電泳槽)、DYY-4型穩(wěn)壓穩(wěn)流電泳儀:北京市六一儀器廠。
1.3 實(shí)驗(yàn)方法
1.3.1 基因組DNA的提取
挑選B.subtilisBJ3-2的單菌落,接種到5 mL LB液體培養(yǎng)基中37℃、180 r/min過(guò)夜培養(yǎng),按照Promega公司的革蘭氏陽(yáng)性細(xì)菌中基因組提取試劑盒說(shuō)明書(shū)提取B.subtilis BJ3-2全基因組。
1.3.2 PCR擴(kuò)增精氨酸脫羧酶基因
根據(jù)枯草芽孢桿菌(Bacillus subtilis)BJ3-2的speA基因序列,設(shè)計(jì)合成含BamHI引物PF:CGCGGATCCATGTCTCA ACATGAAACACCC和含XhoI引物PR:CCGCTCGAGTT GAATTGCTTTTTGTTCTTTG。采用PCR擴(kuò)增引物對(duì)目的基因進(jìn)行擴(kuò)增,PCR反應(yīng)體系:10μmol/μLPF和PR各0.4 μL、10×PCRbuffer2.0μL、dNTPMixture1.6μL、模板DNA1μL、rTaqDNA聚合酶0.1μL、ddH2O14.5μL,總體積20μL。PCR反應(yīng)條件:94℃預(yù)變性5 min,94℃變性40 s,55℃復(fù)性45 s,72℃延伸90 s,30個(gè)循環(huán),72℃延伸10 min。反應(yīng)結(jié)束后用0.7%瓊脂糖凝膠電泳檢測(cè)。
1.3.3 重組質(zhì)粒pGEM-T-speA的構(gòu)建及測(cè)序
PCR產(chǎn)物采用膠回收試劑盒進(jìn)行純化,純化后的PCR產(chǎn)物與pGEM-T連接,連接的過(guò)程及產(chǎn)物的轉(zhuǎn)化過(guò)程按試劑盒的操作說(shuō)明書(shū)進(jìn)行。將連接好的10 μL產(chǎn)物按照常規(guī)方法轉(zhuǎn)化至E.coliDH5α感受態(tài)細(xì)胞中。挑選陽(yáng)性重組子,提取質(zhì)粒后進(jìn)行質(zhì)粒PCR鑒定。將陽(yáng)性菌株提取質(zhì)粒后送至Life Technologies公司進(jìn)行測(cè)序[14]。序列結(jié)果通過(guò)網(wǎng)站http://blast.ncbi.nlm.nih.gov/Blast.cgi的BLAST軟件對(duì)基因序列進(jìn)行在線的比對(duì)和分析,鑒定所克隆的基因是否為目的基因。將測(cè)序鑒定正確的重組質(zhì)粒命名為pGEM-T-speA。
1.3.4 重組質(zhì)粒pET28a-speA的構(gòu)建
將質(zhì)粒pGEM-T-speA和表達(dá)載體pET28a分別用BamH I(37℃)和XhoI(37℃)雙酶切,對(duì)目的片段膠回收并純化,用T4 DNA連接酶將雙酶切產(chǎn)物和載體pET28a連接。按照試劑盒DNA Ligation Kit Ver.2.1中的說(shuō)明書(shū)的使用方法進(jìn)行連接。將連接產(chǎn)物轉(zhuǎn)化至感受態(tài)細(xì)胞E.coli BL21(DE3)中,劃線接種到含有Kan的LB固體平板上,37℃恒溫培養(yǎng)箱中過(guò)夜培養(yǎng),同時(shí)接種到含有Kan(50 μg/mL)的LB液體培養(yǎng)基中,37℃、180 r/min振蕩培養(yǎng)過(guò)夜。將陽(yáng)性菌株提取質(zhì)粒DNA后進(jìn)行酶切鑒定。
1.3.5 重組蛋白精氨酸脫羧酶的表達(dá)
將重組菌pET28a-speA/BL21和pET28a/BL21分別接種于5 mL含Kan(50 μg/mL)的LB液體培養(yǎng)基中,37℃、180 r/min培養(yǎng)至OD600nm=0.4~0.6,用終濃度為1 mmol/L的IPTG,28℃、180 r/min誘導(dǎo)培養(yǎng)4 h。取1 mL菌液4 000 r/min離心收集菌體,加入500μL的pH7.4磷酸鹽緩沖液(phosphatebuffered saline,PBS)溶液重懸后進(jìn)行超聲波破碎(破碎條件:冰浴、200 MHz、超聲3 s,間隔2 s)30 min,10 000 r/min離心10 min,收集上清液即為重組蛋白精氨酸脫羧酶。分別取20 μL上清液和超聲沉淀上清液進(jìn)行十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamidegelelectrophoresis,SDS-PAGE)檢測(cè)(分離膠濃度為15%濃縮膠濃度為5%),考馬斯亮藍(lán)R-250染色、脫色[11],凝膠成像系統(tǒng)拍照并進(jìn)行灰度掃描分析。
1.3.6 重組蛋白的純化及濃度測(cè)定
參照His.Bind Purification Kit試劑盒說(shuō)明書(shū)對(duì)重組蛋白精氨酸脫羧酶進(jìn)行純化,純化條件:依次采用濃度為10 mmol/L、50 mmol/L、200 mmol/L、500 mmol/L的咪唑溶液進(jìn)行洗脫,收集洗脫液,參考文獻(xiàn)[15]的方法進(jìn)行透析除鹽,將透析過(guò)后的蛋白質(zhì)樣品裝入無(wú)菌離心管內(nèi),立即置于液氮中2 min,將其快速?gòu)氐變鐾福D(zhuǎn)入大氣壓為110 kPa的冷凍干燥機(jī),4℃預(yù)冷15~30 min,放入樣品,啟動(dòng)真空泵,37℃干燥24~72 h,取出樣品即為純化后的重組蛋白。采用考馬斯亮藍(lán)法測(cè)定蛋白濃度[15]。
1.3.7 重組蛋白酶活的測(cè)定
參照ADC酶聯(lián)免疫分析試劑盒說(shuō)明書(shū)對(duì)重組蛋白精氨酸脫羧酶活性進(jìn)行測(cè)定。以精氨酸脫羧酶標(biāo)準(zhǔn)試劑的酶活力作為橫坐標(biāo),吸光度值OD450nm作為縱坐標(biāo),繪制標(biāo)準(zhǔn)曲線。將樣品所測(cè)得的OD450nm代入酶活力標(biāo)準(zhǔn)曲線直線回歸方程式,算出樣品酶活力后乘以稀釋倍數(shù)所得值即為樣品實(shí)際酶活力大小。
2 結(jié)果與分析
2.1 B.subtilisBJ3-2基因組DNA的提取及PCR擴(kuò)增
以菌株B.subtilisBJ3-2基因組DNA為模板,PCR擴(kuò)增speA基因,經(jīng)0.7%的瓊脂糖凝膠電泳,結(jié)果見(jiàn)圖1。由圖1可知,泳道1~4出現(xiàn)約1條帶,其中約1 400 bp處擴(kuò)增帶與預(yù)期的speA大小相符。

圖1 speA基因PCR擴(kuò)增電泳圖Fig.1 Electrophoretogram of genespeAby PCR amplification
2.2 重組質(zhì)粒PCR的鑒定
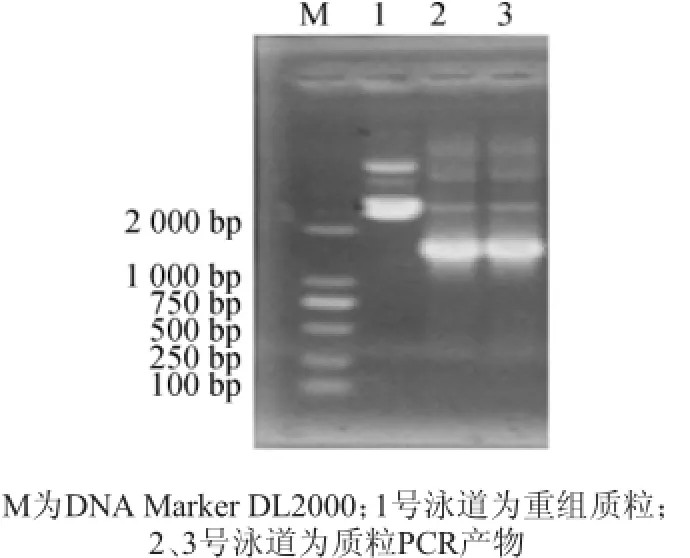
圖2 重組質(zhì)粒PCR擴(kuò)增電泳圖Fig.2Electrophoretogramof recombinant plasmid by PCR amplification
目的DNA片段進(jìn)行膠回收,連接T載體后轉(zhuǎn)化大腸桿菌(Escherichia coli)DH5α。經(jīng)過(guò)菌落PCR鑒定為陽(yáng)性的重組菌落接種到LB培養(yǎng)基中過(guò)夜培養(yǎng),提取質(zhì)粒進(jìn)行質(zhì)粒PCR鑒定,結(jié)果(圖2)可知,在預(yù)期的1 400 bp處出現(xiàn)特異性條帶,說(shuō)明目的基因已經(jīng)正確克隆至pGEM-T載體,重組載體命名為pGEM-speA。
2.3 目的基因的序列分析
測(cè)序結(jié)果表明,B.subtilisBJ3-2精氨酸脫羧酶基因speA大小為1 473 bp,編碼490個(gè)氨基酸,計(jì)算其蛋白質(zhì)分子質(zhì)量為53.53 ku,等電點(diǎn)為5.29,酸性氨基酸19.6%,堿性氨基酸12.2%。與GenBank中登錄號(hào)為KJ561348序列完全一致,說(shuō)明所克隆的基因正確。
2.4 重組表達(dá)載體pET28a-speA構(gòu)建及鑒定
以BamH I和XhoI分別雙酶切陽(yáng)性質(zhì)粒pGEM-T-speA與pET28a載體,經(jīng)轉(zhuǎn)化、抗性篩后進(jìn)行菌落PCR驗(yàn)證,結(jié)果見(jiàn)圖3。取1號(hào)菌過(guò)夜培養(yǎng)后提取質(zhì)粒進(jìn)行雙酶切驗(yàn)證,結(jié)果見(jiàn)圖4。

圖3 pET28a-speA重組質(zhì)粒PCR擴(kuò)增電泳圖Fig.3 Electrophoretogram of recombinant plasmid pET28a-speA by PCR amplification
由圖3可知,挑取得3個(gè)菌落均擴(kuò)增出均出現(xiàn)1條帶,其中約1400bp處擴(kuò)增帶與預(yù)期的speA大小相符,說(shuō)明目的基因已正確克隆至pET28a載體中,重組質(zhì)粒命名為pET28a-speA。
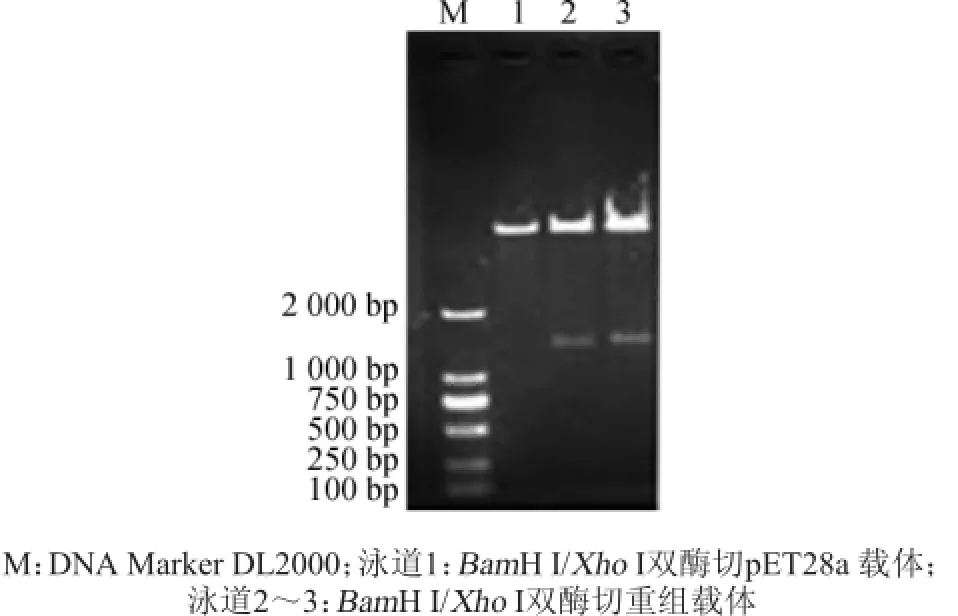
圖4 pET28a-speA重組質(zhì)粒酶切鑒定Fig.4 Enzyme digestion identification of recombinant plasmid pET28a-speA
由圖4可知,酶切產(chǎn)物片段大小與克隆的基因大小相符。泳道2、3均酶切出兩個(gè)條帶,其中較小的條帶出現(xiàn)的位置大約在1 400 bp處,與目的基因(speA)序列大小一致。其中泳道3目的片段稍微偏大,可能由于瓊脂糖凝膠制備原因所致。為確定讀碼框是否正確,取2號(hào)泳道菌進(jìn)行測(cè)序,結(jié)果顯示序列長(zhǎng)度為1473bp,同時(shí)在pET28a載體中的讀碼框也正確,這表明克隆的精氨酸脫羧酶基因與表達(dá)載體pET28a連接成功,即預(yù)期獲得重組表達(dá)載體pET28a-speA重組子。
2.5 目的蛋白誘導(dǎo)表達(dá)
將pET28a-speA/BL21(DE3)重組菌接種于含Kan(50μg/mL)的LB液體培養(yǎng)基,28℃搖瓶大量培養(yǎng),1.0mmol/L IPTG誘導(dǎo)培養(yǎng)后離心,分別取上清、沉淀及未誘導(dǎo)的細(xì)胞裂解液進(jìn)行SDS-PAGE電泳檢測(cè),結(jié)果見(jiàn)圖5。由圖5可知,未添加誘導(dǎo)物的菌液蛋白基本不表達(dá),泳道2、3和4號(hào)均出現(xiàn)顯著的蛋白條帶,說(shuō)明ADC蛋白在上清、菌體內(nèi)都能表達(dá)。通過(guò)軟件Quantity One分析得出目的蛋白的分子大小約為58 ku,與推測(cè)的蛋白質(zhì)分子質(zhì)量大小相當(dāng)。誘導(dǎo)表達(dá)的ADC蛋白菌體為包涵體形式表達(dá),上清為可溶性表達(dá)且表達(dá)量更高,因此選取可溶性部分進(jìn)行后續(xù)實(shí)驗(yàn)。

圖5 蛋白小量誘導(dǎo)表達(dá)SDS-PAGE電泳圖Fig.5 Electrophoretogram of protein inducible expression by SDS-PAGE analysis
2.6 蛋白的純化
對(duì)pET28a-speA/BL21(DE3)重組菌進(jìn)行500 mL大量表達(dá),取上清利用His純化試劑盒進(jìn)行過(guò)柱純化,純化后大部分雜蛋白被除去,結(jié)果如圖6所示。由圖6可知,純化結(jié)果表明,10~500 mmol/L咪唑洗脫均能獲得目的蛋白,其中200 mmol/L咪唑可洗脫獲得較高濃度的蛋白帶。選擇該濃度進(jìn)行剩余蛋白的洗脫,合并所有洗脫液進(jìn)行后續(xù)進(jìn)一步純化處理。
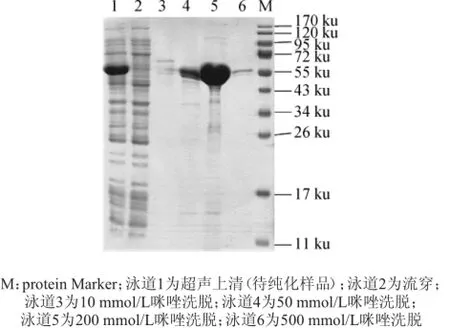
圖6 純化蛋白的SDS-PAGE電泳圖Fig.6 Electrophoretogram of purified protein by SDS-PAGE analysis
2.7 蛋白含量的測(cè)定
采用方法測(cè)定純化后重組蛋白含量,以牛血清蛋白(bovine serum albumin,BSA)質(zhì)量濃度(X)為橫坐標(biāo),吸光度值OD595nm(Y)為縱坐標(biāo),繪制牛血清蛋白標(biāo)準(zhǔn)曲線,根據(jù)標(biāo)準(zhǔn)曲線得出回歸方程為:Y=3.515 4X+0.002,相關(guān)系數(shù)為R2=0.998 4,按照回歸方程計(jì)算ADC蛋白質(zhì)含量為0.97 mg/mL,即純度為97%。
2.8 ADC酶活的檢測(cè)
選用酶聯(lián)免疫分析試劑盒對(duì)重組蛋白ADC酶活進(jìn)行檢測(cè),以精氨酸脫羧酶標(biāo)準(zhǔn)試劑的酶活(X)為橫坐標(biāo),吸光度值OD450nm(Y)為縱坐標(biāo),繪制ADC酶標(biāo)準(zhǔn)曲線。根據(jù)標(biāo)準(zhǔn)曲線得出回歸方程為:Y=0.008 1X+0.036 4,R2=0.980 2,根據(jù)回歸方程計(jì)算ADC酶活為16 780 U/mg。
3 結(jié)論
枯草芽孢桿菌作為豆豉發(fā)酵過(guò)程中的主要菌種,本研究成功克隆獲得了B.subtilisBJ3-2精氨酸脫羧酶基因speA。以pET-28a作為表達(dá)載體,經(jīng)IPGT誘導(dǎo)表達(dá),并經(jīng)15% SDS-PAGE電泳檢測(cè)分析,在200mmol/L咪唑處可洗脫獲得較高濃度蛋白,采用考馬斯亮藍(lán)法測(cè)其濃度為0.972mg/mL,即純度為97%。應(yīng)用ELISA法檢測(cè)精氨酸脫羧酶酶活為16 780 U/mg。通過(guò)對(duì)主效微生物枯草芽孢桿菌產(chǎn)生的主要生物胺相關(guān)酶—ADC進(jìn)行表達(dá)純化以及酶活性研究,可以進(jìn)一步探索酶活性與生物胺的關(guān)聯(lián)性,以ADC活性作為指標(biāo)可以反映出發(fā)酵豆豉的品質(zhì);同時(shí)通過(guò)對(duì)酶活性影響因素的研究,進(jìn)而達(dá)到控制發(fā)酵過(guò)程,從而降低生物胺提高發(fā)酵食品安全性的目的。
[1]王穎,邱璠,韓北忠,等.食品中的生物胺及其檢測(cè)方法[J].中國(guó)釀造,2011,30(10):1-5.
[2]馮婷婷,方芳,楊娟.食品生物制造過(guò)程中生物胺的形成與消除[J].食品科學(xué),2013,34(19):360-366.
[3]王光強(qiáng),俞劍燊,胡健,等.食品中生物胺的研究進(jìn)展[J].食品科學(xué),2016,37(1):269-278.
[4]RIVAS B,MARCOBAL A,CARRASCOSA A V,et al.PCR detection of foodborne bacteria producing the biogenic amines histamine,tyramine, putrescine,and cadaverine[J].J Food Prot,2006,69(10):2509-2514.
[5]唐雪.細(xì)菌型水豆豉發(fā)酵菌群的動(dòng)態(tài)研究[D].貴陽(yáng):貴州大學(xué),2015. [6]韓忠安,羅信旭,楊春艷,等.豆豉中生物胺含量的動(dòng)態(tài)變化研究[J].中國(guó)釀造,2016,35(5):60-64.
[7]劉艷敏,盧彪,沈璽龍,等.枯草芽孢桿菌BJ3-2精氨酸脫羧酶基因speA的克隆與序列分析[J].中國(guó)釀造,2014,33(5):39-43.
[8]馬瑛,劉靜.植物體中多胺代謝及其功能研究進(jìn)展[J].陜西理工學(xué)院學(xué)報(bào):自然科學(xué)版,2010,26(2):47-54.
[9]TIBURCIO A F,MASDéU M A,DUMORTIER F M.Polyamine metabolism and osmotic stress I.Relation to protoplast viability[J]. Plant Physiol,1986,82(2):369-374.
[10]汪天,孟艷瓊,谷風(fēng),等.多胺代謝及其與農(nóng)產(chǎn)品貯藏保鮮關(guān)系的研究[C].合肥:安徽食品安全博士論壇,2005.
[11]孟望霓,吳擁軍,李耀中,等.應(yīng)用原生質(zhì)體融合技術(shù)篩選豆豉芽孢桿菌[J].中國(guó)釀造,2011,30(8):57-60.
[12]賈東旭,吳擁軍,李耀中,等.細(xì)菌型豆豉發(fā)酵芽孢桿菌的篩選與鑒定[J].食品科學(xué),2009,30(5):217-221.
[13]SAMBROOKJ,RUSSELLDW.Molecularcloning:a laboratorymanual [M].New York:Scholastic Corporation Press,2001:106-152.
[14]唐雪,蔡傳斌,羅信旭,等.枯草芽孢桿菌BJ3-2賴氨酸脫羧酶基因yaaO的克隆與序列分析[J].中國(guó)釀造,2014,33(12):116-120.
[15]李慎濤,張富喜,陳振文,等.精編蛋白質(zhì)科學(xué)實(shí)驗(yàn)指南[M].北京:科學(xué)出版社,2007:4-12.
[16]李志軍,吳永寧,薛長(zhǎng)湖.生物胺與食品安全[J].食品與發(fā)酵工業(yè),2004,30(10):84-91.
[17]蘇國(guó)興,劉友良.高等植物體內(nèi)的多胺分解代謝及其主要產(chǎn)物的生理作用[J].植物學(xué)通報(bào),2005,22(4):408-418.
[18]何慶華,吳永寧,印遇龍.食品中生物胺研究進(jìn)展[J].中國(guó)食品衛(wèi)生雜志,2007,19(5):451-454.
Expression and purification of arginine decarboxylase genespeAofBacillus subtilis
CAO Ning,WANG Yajuan,ZHONG Jie,TONG Shuoqiu,WU Yongjun*
(College of Life Science,Guizhou University,Guiyang 550025,China)
Specific enzyme primers were designed according to the arginine decarboxylase(ADC)gene sequence ofBacillus subtilisBJ3-2,the gene speAwas cloned and obtained.The results of sequence analysis indicated that the full-length of genespeAwas 1 473 bp,which could encode 490 amino acids with deduced molecular mass of 58 ku.The genespeAwas cloned into prokaryotic expression vector to obtain recombinant strain pET28a-speA/BL21.The results of SDS-PAGE showed that the target protein was induced with 1.0 mmol/LIPTG at 28℃for 4 h,and the ADC protein could be expressed in supernatant fluid and bacteria.After purification,dialysis and freeze-drying,the ADC with 97%purity was obtained in supernatant fluid.The ADC activity was 16 780 U/mg by ELISA.The study laid a theoretical foundation for expression,purification and enzymatic properties of genespeA.
Bacillus subtilis;arginine decarboxylase;expression;purification
Q936
0254-5071(2017)03-0090-05
10.11882/j.issn.0254-5071.2017.03.019
2016-12-20
國(guó)家自然科學(xué)基金項(xiàng)目(31260394/C200207)
曹寧(1989-),女,碩士研究生,研究方向?yàn)樯锘瘜W(xué)與分子生物學(xué)。
*通訊作者:吳擁軍(1971-),男,教授,博士,研究方向?yàn)槭称飞锛夹g(shù)。大腸桿菌(Escherichia coli)BL21(DE3)進(jìn)行表達(dá),獲得精氨酸脫羧酶,并對(duì)其進(jìn)行純化和酶活力的檢測(cè)對(duì)其ADC基因的研究會(huì)有助于進(jìn)一步了解多胺的合成途徑,可用于后期酶學(xué)性質(zhì)的研究,以期為實(shí)際生產(chǎn)中抑制ADC活性、間接控制腐胺、亞精胺和精胺等相關(guān)生物胺的合成提供理論依據(jù)。
