不同濃度NF-κB信號(hào)通路抑制劑SN50對體外培養(yǎng)小鼠成骨細(xì)胞MC3T3-E1自噬體表達(dá)的影響
2017-04-04 16:31:44秦晗徐宏志龔永慶
山東醫(yī)藥 2017年19期
關(guān)鍵詞:信號(hào)實(shí)驗(yàn)
秦晗 ,徐宏志,龔永慶
(徐州醫(yī)科大學(xué)附屬連云港醫(yī)院,江蘇連云港222002)
不同濃度NF-κB信號(hào)通路抑制劑SN50對體外培養(yǎng)小鼠成骨細(xì)胞MC3T3-E1自噬體表達(dá)的影響
秦晗 ,徐宏志,龔永慶
(徐州醫(yī)科大學(xué)附屬連云港醫(yī)院,江蘇連云港222002)
目的 觀察不同濃度NF-κB信號(hào)通路抑制劑SN50培養(yǎng)的小鼠成骨細(xì)胞MC3T3-E1中自噬體的表達(dá)變化。方法 將體外培養(yǎng)的成骨細(xì)胞MC3T3-E1分為實(shí)驗(yàn)1、2、3組和對照組。對照組不加藥,實(shí)驗(yàn)1、2、3組分別加入終濃度為6.25、12.5、25 μg/mL的SN50,繼續(xù)培養(yǎng)24 h,行單丹( 磺)酰戊二胺(MDC)染色,熒光顯微鏡下觀察各組自噬體表達(dá)情況。結(jié)果 對照組成骨細(xì)胞MC3T3-E1未見明顯點(diǎn)狀分布的綠色熒光。實(shí)驗(yàn)1組可見少許點(diǎn)狀綠色熒光。實(shí)驗(yàn)2組點(diǎn)狀綠色熒光數(shù)量較多。實(shí)驗(yàn)3組可見大量的點(diǎn)狀綠色熒光顆粒散布于成骨細(xì)胞的胞質(zhì)和細(xì)胞突起。對照組、實(shí)驗(yàn)1組、實(shí)驗(yàn)2組、實(shí)驗(yàn)3組綠色熒光細(xì)胞比例分別為1.0%±0.2%、12.3%±1.5%、26.8%±2.4%、56.8%±5.5%。綠色熒光細(xì)胞比例對照組<實(shí)驗(yàn)1組<實(shí)驗(yàn)2組<實(shí)驗(yàn)3組,P<0.05。結(jié)論 NF-κB信號(hào)通路抑制劑SN50培養(yǎng)的小鼠成骨細(xì)胞MC3T3-E1中自噬體表達(dá)且呈劑量依賴性升高。
核因子κB;成骨細(xì)胞;自噬;自噬體
牙齒萌出是指牙齒由骨腔內(nèi)移動(dòng)至骨外、進(jìn)而突破牙齦, 直至達(dá)到功能頜的過程[1]。成骨細(xì)胞和破骨細(xì)胞之間的動(dòng)態(tài)平衡是牙齒正常萌出通道形成的基礎(chǔ),核因子κB(NF-κB)信號(hào)通路是調(diào)節(jié)二者平衡的重要通路[2],其通過調(diào)控RANK/RANKL表達(dá),從而調(diào)節(jié)成骨、破骨細(xì)胞分化信號(hào)進(jìn)而影響牙齒萌出通道的形成,在牙齒萌出過程中發(fā)揮著關(guān)鍵作用[3,4]。但是NF-κB調(diào)節(jié)成骨細(xì)胞分化的機(jī)制尚不明確。新近有研究發(fā)現(xiàn)NF-κB通路調(diào)控神經(jīng)細(xì)胞分化的機(jī)制可能和抑制細(xì)胞周期、激活自噬有關(guān)[5],提示我們NF-κB調(diào)節(jié)成骨細(xì)胞分化可能與自噬有關(guān)。本研究觀察了不同濃度NF-κB信號(hào)通路抑制劑SN50培養(yǎng)的成骨細(xì)胞MC3T3-E1中自噬體的表達(dá)變化。現(xiàn)報(bào)告如下。
1 材料與方法
1.1 材料 成骨細(xì)胞MC3T3-E1購于中科院上海
1.2 成骨細(xì)胞分組及SN50應(yīng)用方法 將成骨細(xì)胞MC3T3-E1用?- MEM培養(yǎng)基于37 ℃、5%CO2培養(yǎng)箱中靜置培養(yǎng)。待細(xì)胞生長良好,用0.25%胰蛋白酶消化制成單細(xì)胞懸液,以2×104/cm2的密度接種與已置入消毒蓋玻片的24孔培養(yǎng)板中。隨機(jī)分為實(shí)驗(yàn)1組、實(shí)驗(yàn)2組、實(shí)驗(yàn)3組、對照組,每組6孔。對照組不加藥,實(shí)驗(yàn)1、2、3組分別加入終濃度為6.25、12.5、25 μg/mL的SN50,繼續(xù)培養(yǎng)24 h。
1.3 細(xì)胞自噬體表達(dá)觀察方法 采用單丹( 磺)酰戊二胺(MDC)染色法。將MDC染色液按體積比稀釋10倍。每孔加入100 μL MDC染色液,室溫避光染色15~45 min后,棄去染色液,以300 μL的1×Wash Buffer洗3次,再以100 μL的collection buffer覆蓋細(xì)胞爬片,激光共聚焦顯微鏡下觀察并拍照。激發(fā)濾光片波長355 nm。阻斷濾光片波長512 nm。
高倍鏡(400倍)下計(jì)數(shù)1 000個(gè)細(xì)胞,計(jì)算綠色熒光細(xì)胞占總細(xì)胞數(shù)的百分比。
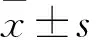
2 結(jié)果
對照組成骨細(xì)胞MC3T3-E1未見明顯點(diǎn)狀分布的綠色熒光。實(shí)驗(yàn)1組可見少許點(diǎn)狀綠色熒光。實(shí)驗(yàn)2組點(diǎn)狀綠色熒光數(shù)量較多。實(shí)驗(yàn)3組可見大量的點(diǎn)狀綠色熒光顆粒散布于成骨細(xì)胞的胞質(zhì)和細(xì)胞突起。對照組、實(shí)驗(yàn)1組、實(shí)驗(yàn)2組、實(shí)驗(yàn)3組綠色熒光細(xì)胞比例分別為1.0%±0.2%、12.3%±1.5%、26.8%±2.4%、56.8%±5.5%。綠色熒光細(xì)胞比例對照組<實(shí)驗(yàn)1組<實(shí)驗(yàn)2組<實(shí)驗(yàn)3組,P<0.05。
3 討論
牙齒的萌出是一個(gè)多因素過程,在這一過程中既有牙槽骨、牙胚的組織學(xué)改變, 又有破骨細(xì)胞、成骨細(xì)胞、牙胚細(xì)胞等細(xì)胞學(xué)的改變, 還有多種細(xì)胞因子的參與,而且這些生物信號(hào)分子是通過細(xì)胞間、細(xì)胞與基質(zhì)間、基質(zhì)間的自分泌或旁分泌途徑來調(diào)控發(fā)育進(jìn)程[6,7]。成骨細(xì)胞和破骨細(xì)胞之間的動(dòng)態(tài)平衡是保證牙齒正常萌出過程的基礎(chǔ),若這一平衡失調(diào),則可能導(dǎo)致牙齒萌出障礙。牙齒萌出障礙是指牙齒萌出期已過而仍在頜骨內(nèi)未能萌出的牙齒,其代表性疾病為顱骨鎖骨發(fā)育不全(CCD)則是多數(shù)牙齒萌出永久性的延遲[8]。我們曾對CCD患者進(jìn)行血清檢測發(fā)現(xiàn),在成骨細(xì)胞的標(biāo)志性酶血清骨特異性堿性磷酸酶(B-ALP)升高的同時(shí),伴有破骨細(xì)胞的標(biāo)志性酶-抗酒石酸酸性磷酸酶-5b(TRCAP-5b)升高[9]。提示成骨細(xì)胞和破骨細(xì)胞不能協(xié)調(diào)分化可能是該類患者牙齒萌出障礙的主要原因。目前關(guān)于牙齒萌出障礙研究的重點(diǎn)主要集中于牙齒萌出過程中破骨細(xì)胞的數(shù)量和分化的相關(guān)分子機(jī)制的變化情況。相反,對于密切相關(guān)的成骨細(xì)胞如何啟動(dòng)分化誘導(dǎo)破骨細(xì)胞的聚集、分化、促進(jìn)牙齒萌出通道的形成,以及在這過程中信號(hào)通路情況及是否伴有自噬的發(fā)生等分子調(diào)控機(jī)制的研究則相對較少。
NF-κB信號(hào)通路是骨量動(dòng)態(tài)平衡調(diào)節(jié)的重要通路,其通過對RANK/RANKL表達(dá)的調(diào)控,調(diào)節(jié)成骨和破骨細(xì)胞的分子信號(hào),在骨基質(zhì)的形成、發(fā)育和改建中發(fā)揮關(guān)鍵調(diào)控作用[10]。因此,同樣也是牙齒萌出通道形成過程中的關(guān)鍵信號(hào)通路。破骨細(xì)胞核因子κB受體活化因子配體(RANKL)主要由免疫細(xì)胞、骨髓間質(zhì)細(xì)胞、成骨細(xì)胞、血管內(nèi)皮細(xì)胞及一些腫瘤細(xì)胞表達(dá),通常以三聚體復(fù)合物的形式存在于細(xì)胞質(zhì)中,可以被許多因素激活,包括炎性細(xì)胞因子、生長因子、免疫受體、細(xì)菌及其產(chǎn)物脂多糖、病毒與病毒蛋白及某些理化因素等[11]。破骨細(xì)胞核因子κB受體活化因子(RANK)是RANKL刺激破骨細(xì)胞分化、生存、融合與活化的唯一靶受體。RANK的活化能使細(xì)胞骨架重構(gòu),改變破骨細(xì)胞的形態(tài),如細(xì)胞的延展性、移動(dòng)性等,使破骨細(xì)胞移動(dòng)并黏附到骨吸收區(qū)域。位于破骨細(xì)胞前體細(xì)胞細(xì)胞膜表面的RNAK與RNAKL結(jié)合后可促進(jìn)破骨細(xì)胞前體細(xì)胞的分化與成熟[12,13]。在前期的研究中,我們圍繞牙齒萌出過程中破骨細(xì)胞和NF-κB信號(hào)通路開展了一系列的基礎(chǔ)和臨床研究。結(jié)果顯示RANKL的陽性信號(hào)廣泛表達(dá)于成牙本質(zhì)細(xì)胞、牙髓成纖維細(xì)胞、牙周韌帶成纖維細(xì)胞、牙齦成纖維細(xì)胞、牙囊細(xì)胞、成骨細(xì)胞和成釉細(xì)胞等多種牙齒、牙周相關(guān)細(xì)胞。體外培養(yǎng)的破骨細(xì)胞加入NF-κB信號(hào)通路阻斷劑后其數(shù)量減少,活性降低,RANKL表達(dá)較對照組明顯減弱[14]。體內(nèi)的動(dòng)物實(shí)驗(yàn)也同樣證明,加入NF-κB信號(hào)通路阻斷劑后,小鼠的牙齒萌出較對照組減慢,牙囊和頜骨周圍的RANKL表達(dá)較對照組明顯減弱[15]。驗(yàn)證了NF-κB信號(hào)通路可能是調(diào)控破骨細(xì)胞活化的重要通路,同時(shí)也證實(shí)破骨細(xì)胞抑制劑可能通過調(diào)節(jié)RANKL的表達(dá)、調(diào)控破骨細(xì)胞的分化,進(jìn)一步說明NF-κB信號(hào)通路在調(diào)節(jié)成骨細(xì)胞和破骨細(xì)胞的動(dòng)態(tài)平衡、促進(jìn)牙齒萌出過程中具有重要作用。
新近有研究發(fā)現(xiàn)NF-κB抑制劑觸發(fā)神經(jīng)干細(xì)胞分化的機(jī)制可能和抑制細(xì)胞周期、激活自噬有關(guān)[5];另有研究發(fā)現(xiàn)自噬基因可以在牙齒發(fā)育過程中的不同部位表達(dá),提示自噬可能參與了牙齒的發(fā)生、發(fā)育[16,17]。然而自噬在牙齒萌出過程中發(fā)揮怎樣的作用,以及自噬與牙齒萌出過程中成骨細(xì)胞、NF-κB信號(hào)通路之間存在一個(gè)怎樣的聯(lián)系?目前研究較少。眾所周知,自噬是廣泛存在于真核細(xì)胞內(nèi)的一種溶酶體依賴性降解途徑,是細(xì)胞進(jìn)行自我保護(hù)的一種重要機(jī)制,在維持細(xì)胞存活、更新、物質(zhì)再利用和內(nèi)環(huán)境穩(wěn)定中起著重要作用[18]。自噬具有雙重作用,當(dāng)自噬適度激活時(shí),可加速胞內(nèi)蛋白質(zhì)降解,產(chǎn)生氨基酸等小分子物質(zhì),抵抗能量不足,防止錯(cuò)誤蛋白堆積,進(jìn)而促進(jìn)細(xì)胞存活;當(dāng)自噬過度激活時(shí),可導(dǎo)致細(xì)胞裂解,引起自噬性細(xì)胞死亡[19,20]。根據(jù)細(xì)胞內(nèi)底物運(yùn)送到溶酶體方式的不同,哺乳動(dòng)物細(xì)胞自噬可分為三種主要方式:巨自噬、微自噬和分子伴侶介導(dǎo)的自噬,通常自噬指的是巨自噬。在巨自噬中,細(xì)胞漿中可溶性蛋白和變性壞死的細(xì)胞器被非溶酶體來源的雙層膜結(jié)構(gòu)所包裹,即自噬泡,并有自噬泡將其攜帶到溶酶體中降解[21,22]。自噬/大自噬在維持細(xì)胞的穩(wěn)定性、發(fā)育、細(xì)胞生存、衰老、免疫、腫瘤和神經(jīng)退行性變中發(fā)揮作用。但是直到目前,自噬參與這些作用的機(jī)制還不十分清楚。許多外界刺激(如饑餓、激素、手術(shù))和細(xì)胞內(nèi)刺激(如錯(cuò)誤折疊蛋白的積聚,一些小細(xì)胞器的損傷)均可誘導(dǎo)自噬發(fā)生。
本研究在體外培養(yǎng)的成骨細(xì)胞內(nèi)加入不同濃度的NF-κB信號(hào)通路阻斷劑SN50,通過MDC熒光染色法和激光共聚焦顯微鏡下觀察點(diǎn)狀分布綠色熒光即自噬體的表達(dá)情況。本研究結(jié)果顯示,未加SN50組成骨細(xì)胞胞質(zhì)中未見明顯點(diǎn)狀分布的綠色熒光。隨著SN50濃度的增加,成骨細(xì)胞中點(diǎn)狀分布的綠色熒光體數(shù)和熒光強(qiáng)度均逐漸增加。在加入25 μg/mL的SN50組可以看到大量的點(diǎn)狀綠色熒光顆粒分布于成骨細(xì)胞的胞質(zhì)和細(xì)胞突起。提示SN50阻斷成骨細(xì)胞NF-κB通路過程中自噬被大量的激活。這也進(jìn)一步說明NF-κB信號(hào)通路和自噬在成骨細(xì)胞分化調(diào)節(jié)中發(fā)揮重要作用。
[1] Oosterkamp BC, Ockeloen CW, Carels CE, et al. Tooth eruption disturbances and syndromes[J]. Ned Tijdschr Tandheelkd, 2014,121(4):233-238.
[2] Zhou C, Liu W, He W, et al. Saikosaponin a inhibits RANKL-induced osteoclastogenesis by suppressing NF-κB and MAPK pathways [J]. Int Immunopharmacol, 2015,25(1):49- 54.
[3] Wu C, Wang W, Tian B, et al. Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL-induced osteoclastogenesis in vitro[J]. Biochem Pharmacol,2015,93(1):59-71.
[4] Huh JE, Lee WI, Kang JW, et al. Formononetin attenuates osteoclastogenesis via suppressing the RANKL-induced activation of NF-κB, c-Fos, and nuclear factor of activated T-cells cytoplasmic 1 signaling pathway [J].J Nat Prod,2014,77 (11):2423-2431.
[5] Vucicevic L, Misirkic-Marjanovic M, Paunovic V, et al. Autophagy inhibition uncovers the neurotoxic action of the antipsychotic drug olanzapine [J]. Autophagy,2014,10(12):2362-2378.
[6] Dipti Shastri, Pradeep Tandon, Gyan P, et al. A newer simultaneous space creation, eruption, and adjacent root control spring for the management of impacted tooth [J]. Contemp Clin Dent,2014,5(4): 555-557.
[7] Heikinheimo K, Kurppa KJ, Laiho A, et al. Early dental epithelial transcription factors distinguish ameloblastoma from keratocystic odontogenic tumor [J]. J Dent Res,2015,94(1):101- 111.
[8] Lee KE,Seymen F,Ko J,et al. RUNX2 mutations in cleidocranial dysplasia[J]. Genet Mol Res,2013,12(4):4567-4574.
[9] 李陽飛,秦晗,徐宏志.顱骨鎖骨發(fā)育不良患者特異性堿性磷酸酶和抗酒石酸酸性磷酸酶的水平:1例報(bào)告 [J].中國組織工程研究與臨床康復(fù),2011,15(41):7742-7744.
[10] Jimi E,Fukushima H. NF-κB signaling pathways and the future perspectives of bone disease therapy using selective inhibitors of NF-κB [J]. Clin Calcium,2016, 26(2): 298- 304.
[11] Efeyan A, Comb WC, Sabatini DM. Nutrient-sensing mechanisms and pathways [J]. Nature,2015,517(7534):302- 310.
[12] Zhan X, Zhang C, Dissanayaka WL, et al. Storage media enhance osteoclastogenic potential of human periodontal ligament cells via RANKL-independent signaling [J]. Dent Traumatol,2013,29(1):59- 65.
[13] Kim HN, Lee JH, Jin WJ, et al. α-Tocopheryl succinate inhibits osteoclast formation by suppressing receptor activator of nuclear factor-kappaB ligand (RANKL) expression and bone resorption[J].J Bone Metab,2012,19(2):111-120.
[14] Qin H, Cai J, Fang J, et al. Could MTA be a novel medicine on the recurrence therapy for GCTB [J]. Med Hypotheses,2010,74(2):368- 369.
[15] Qin H,Yang FS. Calcitonin may be a useful therapeutic agent for osteoclastogenesis syndromes involving premature eruption of the tooth [J].Med Hypotheses,2008,70(5):1048-1050
[16] Yang JW, Zhu LX, Yuan GH, et al. Autophagy appears during the development of the mouse lower first molar [J]. Histochem Cell Biol,2013,139(1):109- 118.
[17] Yang J, Wan C, Nie S, et al. Localization of Beclin1 in mouse developing tooth germs: possible implication of the interrelation between autophagy and apoptosis [J]. J Mol Histol,2013,44(6):619- 627.
[18] Wei T, Kang Q, Ma B, et al. Activation of autophagy and paraptosis in retinal ganglion cells after retinal ischemia and reperfusion injury in rats [J]. Exp Ther Med, 2015,9(2):476- 482.
[19] Akl MR, Ayoub NM, Ebrahim HY, et al. Araguspongine C induces autophagic death in breast cancer cells through suppression of c-Met and HER2 receptor tyrosine kinase signaling[J]. Mar Drugs,2015,13(1):288- 311.
[20] Shi M, Cheng L, Zhang Z, et al. Ferroferric oxide nanoparticles induce prosurvival autophagy in human blood cells by modulating the Beclin 1/Bcl-2/VPS34 complex[J]. Int J Nanomedicine,2014,24(10):207- 216.
[21] Lee JM, Wagner M, Xiao R, et al. Nutrient-sensing nuclear receptors coordinate autophagy[J]. Nature,2014,516(7529):112- 115.
[22] Mintern JD, Harris J.Autophagy and immunity[J].Immunol Cell Biol,2015,93(1):1-2.
國家自然科學(xué)基金資助項(xiàng)目(81500893)。
10.3969/j.issn.1002-266X.2017.19.013
R965
A
1002-266X(2017)19-0045-03
2016-10-10)
細(xì)胞庫,MDC染色試劑盒購于美國Sigma公司,SN50購于美國Senta Cruz公司,胎牛血清和a-MEM培養(yǎng)基購于美國Invitrogen 公司。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價(jià)·高一版(2020年6期)2020-11-02 02:45:24
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年3期)2019-02-01 06:12:26
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
海軍航空大學(xué)學(xué)報(bào)(2015年3期)2015-11-11 17:20:00
