田間煙蚜種群對3種殺蟲劑抗性與酶活性關系的研究
2017-03-31 01:10:58劉童童楊茂發
廣東農業科學 2017年1期
張 超,劉童童,2,楊 洪,3,王 召,4,楊茂發,3
(1.貴州大學昆蟲研究所,貴州 貴陽 550025;2.貴州省林業科學研究院,貴州 貴陽 550000;3.貴州大學煙草學院,貴州 貴陽 550025;4.凱里學院環境與生命科學學院,貴州 凱里 556011)
田間煙蚜種群對3種殺蟲劑抗性與酶活性關系的研究
張 超1,劉童童1,2,楊 洪1,3,王 召1,4,楊茂發1,3
(1.貴州大學昆蟲研究所,貴州 貴陽 550025;2.貴州省林業科學研究院,貴州 貴陽 550000;3.貴州大學煙草學院,貴州 貴陽 550025;4.凱里學院環境與生命科學學院,貴州 凱里 556011)
為了探討煙蚜對殺蟲劑抗性與酶活關系,就田間煙蚜種群對樂果、滅多威和阿維菌素3種殺蟲劑的抗性與酶活性的關系進行研究。對具有不同抗性水平的幾個田間煙蚜種群的酶活性測定結果顯示:抗樂果、滅多威、阿維菌素品系中AchE、GSTs和CAT 3種酶的活性都高于敏感品系,其中抗樂果的興仁和湄潭抗性品系AchE酶活性為敏感品系的1.212、1.588倍,GSTs酶活性為敏感品系的1.625、1.900倍,CAT酶活性是敏感品系的1.439、2.018倍;抗滅多威品系的清鎮和湄潭抗性品系AchE酶活性為敏感品系的1.078、1.647倍,GSTs酶活性為敏感品系的 1.142、1.935倍,CAT酶活性為敏感品系的1.142、1.935倍;抗阿維菌素的興仁和長順抗性品系AchE 酶活性為敏感品系的1.370、1.664倍,GSTs酶活性為敏感品系的1.830、2.318倍,CAT酶活性為敏感品系的1.800倍、3.708倍。研究結果可為闡明田間煙蚜種群抗性產生機制提供參考。
煙蚜;抗藥性;酶活性;乙酰膽堿酯酶;過氧化氫酶;谷胱甘肽S-轉移酶
煙蚜Myzus persicae(Sulzer)又名桃蚜,屬半翅目(Hemiptera)蚜科(Aphidiade)瘤蚜屬(Myzus),是一種重要的農業害蟲,更是煙草的主要害蟲。煙蚜除吸取煙株汁液引起植株營養不良及生長畸形造成直接危害外,還可傳播煙草黃瓜花葉病毒(CMV)、馬鈴薯Y病毒(PVY)等多種煙草病毒病,對煙草造成更為嚴重的間接危害[1]。目前,在田間對煙蚜的綜合治理中以化學防治為主,這也是農民選擇的首要手段。但殺蟲劑不合理使用和施藥器械落后等原因使得煙蚜長期處于高選擇和高殘留環境中,加上煙蚜具有較強的環境適應能力和較短的生活史,敏感性快速減弱,耐藥性問題亦日趨嚴重[2]。酶(Enzyme)是一類具有特定催化活性的蛋白質,昆蟲的生命活動幾乎都伴隨著酶的反應。因此,對生物的酶系進行研究不僅對闡明生物生命現象的本質十分重要,也是闡明生物對逆境產生適應性機制的核心[3-4]。過去的研究表明,逆境脅迫下昆蟲體內的主要酶系其活性和含量都會發生變化,這是昆蟲抗性形成的重要生化機制[5]。郭天鳳等和宋春滿等對煙蚜的研究結果表明具有不同抗性水平煙蚜室內種群體內解毒酶活力有較大差異[6-8],王澤華等和張鵬等對昆蟲的研究結果也顯示,不同齡期昆蟲對殺蟲劑敏感性不同[9-10]。蟲態和殺蟲劑敏感性水平是煙蚜防治適期選擇、提高殺蟲劑防效、治理和延緩耐藥性的重要依據。
貴州省主要煙區的田間煙蚜種群已對樂果、滅多威和阿維菌素已產生低倍抗性[11]。本研究以田間煙蚜抗性種群為研究對象,測定已對樂果、滅多威和阿維菌素產生抗性的田間煙蚜種群體內的乙酰膽堿酯酶(AChE)、谷胱甘肽S-轉移酶(GST)和過氧化氫酶(CAT)活性的變化,探索煙蚜對這3類殺蟲劑產生抗藥性的生化機制,并對二者間關系進行分析,旨在為貴州煙區煙蚜的田間有效防控和耐藥性管理提供理論依據。
1 材料與方法
1.1 試驗材料
1.1.1 供試蟲源及飼養方法 相對敏感品系的煙蚜種群:采自長順縣馬路鄉煙葉生產基地,在不接觸任何藥劑的情況下已在實驗室內連續飼養5年。飼養條件:溫度:25(±1)℃、相對濕度:60%~70%、光周期14L∶10D。
抗性品系的煙蚜種群:選取對樂果具有抗性的兩個田間煙蚜種群:湄潭抄樂煙蚜種群(對樂果抗性倍數為6.946倍)、興仁四聯煙蚜種群(對樂果抗性倍數為3.214倍),選取對滅多威具有抗性的兩個田間煙蚜種群:湄潭抄樂煙蚜種群(對滅多威抗性倍數為5.05倍)、清鎮流長煙蚜種群(對滅多威抗性倍數為2.616倍),選取對阿維菌素具有抗性的兩個田間煙蚜種群:長順改堯煙蚜種群(對阿維菌素抗性倍數為6.043倍)、興仁四聯煙蚜種群(對阿維菌素抗性倍數為2.952倍),以上煙蚜種群均于2013年7月采自貴州省煙田。
1.1.2 供試儀器 電子分析天平,德國Sartorious公司;電熱恒溫水溫箱,北京永光明醫療儀器廠;電熱鼓風干燥箱101-OA型,天津市泰斯特儀器有限公司;Haier冰箱,海爾集團;酶標儀,奧地利Tecan公司;試管、容量瓶等其他儀器均為國產;Sigma-3K30 高速冷凍離心機,德國Sartorious公司;移液槍,大龍興創實驗儀器(北京)有限公司;自動雙純水系統purelab plus p15123,Pall Comp(USA)公司;RGX250人工培養箱,天津市泰斯特儀器有限公司;755型紫外分光光度計。移液器 2.5、10、100、200、1000 μL,大龍興創試驗儀器(北京)有限公司50~300 μL八孔道移液器,大龍醫療設備(上海)有限公司
1.1.3 主要試劑 BCA蛋白濃度測定試劑盒,北京鼎國昌盛生物技術有限責任公司;乙酰膽堿酯酶測試盒,南京建成科技有限公司;谷胱甘肽-S轉移酶測定試劑盒,南京建成科技有限公司;過氧化氫酶(Catalase CAT)測試盒,南京建成科技有限公司;氯化鈉注射液(0.9%),貴州科倫藥液有限公司。
1.2 試驗方法
1.2.1 酶液制備 分別取煙蚜敏感種群、湄潭抄樂煙蚜種群和興仁四聯煙蚜種群無翅成蚜10頭,加入900 μL的生理鹽水,用玻璃勻漿器置冰上勻漿,4℃條件下3 000 r/min離心10 min,取上清液為酶源液。
1.2.2 酶源蛋白含量測定 配制BCA工作液:根據標準品和樣品數量,按50體積試劑A,1體積試劑B配制適量BCA工作液,充分混勻。將蛋白標準品按0、1、2、4、6、8、10μL加到96孔板的蛋白標準品孔中,加滅菌雙蒸水補足到10μL;取10μL待測樣品加入96孔板。每個測定3次重復。向待測樣品孔和蛋白標準品孔中加入200μL BCA工作液混勻。60℃溫浴30 min,冷卻至室溫。
酶標儀562 nm波長下測定吸光度。以標準蛋白濃度為X軸,OD值為Y軸,制作蛋白質標準曲線,根據各樣品的OD值,從標準曲線中求出樣品濃度。
1.2.3 酶活性測定 乙酰膽堿酯酶(AchE)、谷胱甘肽-S-轉移酶(GST)和過氧化氫酶(CAT)3種酶的活性均采用南京建成科技有限公司的試劑盒進行測定。
采用Microsoft Excel 2007進行數據處理,采用SPSS16.0軟件進行單因素方差分析和多重比較。
2 結果與分析
2.1 煙蚜對樂果的抗性與酶活性的關系
測定敏感品系煙蚜和田間抗樂果品系體內的乙酰膽堿酯酶、谷胱甘肽S-轉移酶和過氧化氫酶活性,結果(表1)表明,興仁煙蚜與湄潭煙蚜品系均高于敏感品系體內的乙酰膽堿酯酶,比值分別為1.212和1.588。煙蚜體內谷胱甘肽S-轉移酶活性測定結果顯示,湄潭煙蚜品系最高,為敏感品系煙蚜的1.90倍,興仁煙蚜品系較低為1.625倍。同理,興仁煙蚜與湄潭煙蚜品系的過氧化氫酶活性分別是相對敏感品系的1.439倍和2.018倍。由此可見,隨著3個煙蚜種群抗性水平的增加,其體內乙酰膽堿酯酶、谷胱甘肽S-轉移酶和過氧化氫酶3種酶活性均呈現升高趨勢。表明煙蚜對樂果產生抗性后,其體內保護酶和解毒酶的活性增強。
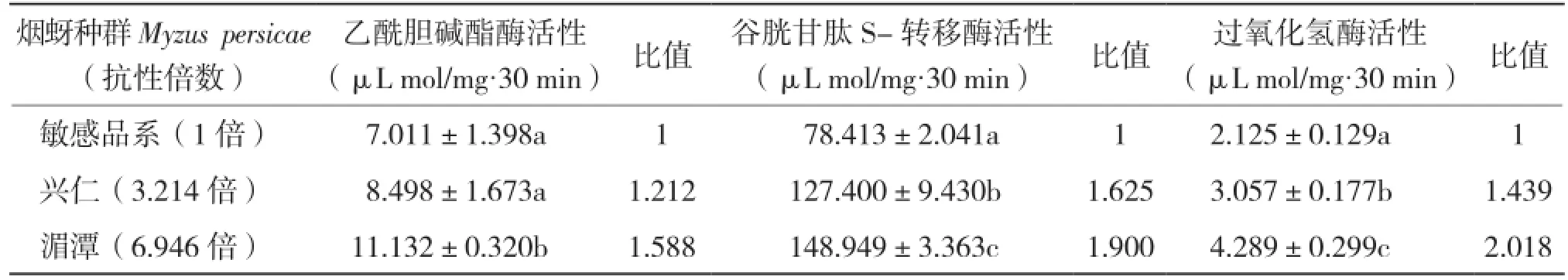
表1 抗樂果煙蚜種群的3種酶活性比較
2.2 煙蚜對滅多威的抗性與酶活性的關系
測定敏感品系煙蚜和田間抗滅多威品系體內的乙酰膽堿酯酶、谷胱甘肽S-轉移酶和過氧化氫酶活性,結果(表2),清鎮煙蚜與湄潭煙蚜品系均高于敏感品系體內的乙酰膽堿酯酶,比值分別為1.078和1.641。煙蚜體內谷胱甘肽S-轉移酶活性測定結果顯示,湄潭煙蚜品系最高,為敏感品系煙蚜的1.935倍,興仁煙蚜品系較低為1.142倍。同理,興仁煙蚜與湄潭煙蚜品系的過氧化氫酶活性分別是相對敏感品系的1.553倍和2.456倍。由此可見,隨著3個煙蚜種群抗性水平的增加,其體內乙酰膽堿酯酶、谷胱甘肽S-轉移酶和過氧化氫酶3種酶活性均呈現升高趨勢。表明煙蚜對滅多威產生抗性后,其體內保護酶和解毒酶的活性增強。

表2 抗滅多威煙蚜種群的3種酶酶活性比較
2.3 煙蚜對阿維菌素的抗性與酶活性的關系
測定敏感品系煙蚜和田間抗阿維菌素品系體內的乙酰膽堿酯酶、谷胱甘肽S-轉移酶和過氧化氫酶活性,結果(表3)表明,興仁煙蚜與長順煙蚜品系均高于敏感品系體內的乙酰膽堿酯酶,比值分別為1.370和1.664。煙蚜體內谷胱甘肽S-轉移酶活性測定結果顯示,長順煙蚜品系最高,為敏感品系煙蚜的2.318倍,興仁煙蚜品系較低為1.830倍。同理,興仁煙蚜與湄潭煙蚜品系的過氧化氫酶活性分別是相對敏感品系的1.800倍和3.708倍。由此可見,隨著3個煙蚜種群抗性水平的增加,其體內乙酰膽堿酯酶、谷胱甘肽S-轉移酶和過氧化氫酶3種酶活性均呈現升高趨勢。表明煙蚜對阿維菌素產生抗性后,其體內保護酶和解毒酶的活性增強。

表3 抗阿維菌素煙蚜種群的3種酶酶活性比較
3 討論
害蟲對殺蟲劑產生抗藥性是不斷適應殺蟲劑的過程。昆蟲的一切生命活動均有酶的參與,當藥劑進入蟲體內,經過體內酶的氧化、還原、水解或結合后,降低其毒性,或增強其水溶性,將其排出體外,達到解毒目的[12-13]。研究發現在殺蟲劑的選擇壓力下,昆蟲體內解毒酶酶活性產生了相應的變化,導致抗性昆蟲中解毒酶酶活性有所增高,引起了害蟲對殺蟲劑抗性的產生[14]。李周直等用藥劑處理褐刺蛾和褐邊綠刺蛾后發現蟲體內保護酶的活性均高于敏感體系,推測昆蟲體內的保護酶的變化與產生抗藥性有很大的關系[15]。由此可見昆蟲體內酶活性的變化是昆蟲產生抗藥性的原因之一。
Smissaert首先在二斑葉螨Tetranychus urticae上發現了導致對殺蟲劑不敏感的AChE變構[16],隨后在許多種昆蟲中如家蠅(Musca domestica)、按蚊(Anopheles)、小菜蛾(Plutella xylostella)和棉蚜(Aphis gossypii)等昆蟲中陸續發現了不敏感AChE的存在[17]。除了酶性質發生改變外,昆蟲也可通過AChE基因表達量的增加而對殺蟲劑產生抗性。在AChE所介導的抗性機理研究中,還有另一些原因如AChE活性的增加、AChE酶量的增加都可能與昆蟲抗性水平成正比[18-19]。本研究通過對具有不同抗性水平的幾種煙蚜種群的乙酰膽堿酯酶活性測定結果顯示,部分抗性品系的AChE活性與敏感品系有顯著性差異,有的差異不顯著。但是隨著抗性提高,AChE的活性都有一定程度的提高。這可能與殺蟲劑的種類有關,大量的研究表明,AChE對有機磷和氨甲基酸酯類殺蟲劑敏感性的降低是昆蟲對這些殺蟲劑產生抗性的重要機理之一。
谷胱甘肽S-轉移酶是昆蟲對殺蟲劑產生抗性的重要因素,在保護組織以抵御氧化侵害及氧化壓力中起重要作用。本研究中3種殺蟲劑的抗性品系中,所有抗性煙蚜品系的GSTs的活性均顯著高于相對,GSTs的活性明顯高于敏感品系煙蚜的GSTs活性,并且同一殺蟲劑的抗性煙蚜品系之間,GSTs的活性亦達到顯著水平。抗性品系中GSTs活性提高,這在其他的抗性昆蟲中也呈現相同的規律[20-21]。說明GSTs可能是是昆蟲產生抗性的一個很重要的因素。
過氧化氫酶作為生物體內重要物質,具有非常重要的生理功能,其中最為主要的就是參與活性氧代謝過程。李周直等證明菜粉蝶、刺蛾體內CAT與它們的耐藥性有關[15]。陳尚文認為馬尾松毛蟲體內CAT與其耐藥性存在一定的相關性[22]。吳小鋒等發現家蠶血液CAT 與蠶體抗逆性有一定的相關性[23]。徐欣等用敵敵畏和高效氯氰菊酯殺蟲劑在不同抗性選育方式下對家蠅抗藥性發展和CAT活性變化的作用,經處理后CAT活性均有不同程度的增加[24]。在本實驗中樂果和滅多威的抗性品系中,所有抗性煙蚜品系的CAT的活性均顯著高于相對敏感品系煙蚜的CAT活性,并且同一殺蟲劑的抗性煙蚜品系之間,CAT的活性差異性亦達到顯著水平;在阿維菌素的抗性品系中,長順煙蚜品系的CAT活性與其他兩個煙蚜品系的CAT活性差異顯著;相對敏感品系煙蚜的CAT活性與興仁煙蚜品系的CAT活性差異不顯著。
本研究對具有不同抗性水平的田間煙蚜種群的酶活性進行測定表明,對樂果、阿維菌素、滅多威具有抗性的煙蚜種群所測定的3種酶活性均比敏感品系高,由次可見當煙蚜對藥劑抗性增強時煙蚜體內的AchE、GSTs和CAT 3種酶的酶活性均比敏感品系高,而且酶活性隨著抗性的增強呈正相關關系。由于所選取的田間煙蚜種群對這3種殺蟲劑的抗性都是低倍抗性或者只是敏感性降低。因此,需要繼續監測這些煙蚜種群對殺蟲劑抗性變化,進一步研究以上3種酶的活性變化的分子機制,才能探明田間煙蚜種群對這3種殺蟲劑產生抗性的機理,為田間煙蚜防控的用藥策略提供參考。
[1] 秦煥菊,王桂芬. 我國煙田蚜蟲與病毒病害[J]. 中國煙草學報,1996,3(3):75-78.
[2] Bass C,Puinean AM,Zimmer CT,et al. The evolution of insecticideresistance in the peach potato aphid,Myzus persicae[J]. InsectBiochemistry and Molecular Biology,2014,51:41-51.
[3] 劉冰,梁嬋娟. 生物過氧化氫酶研究進展[J].中國農學通報,2005,21(5):223-225.
[4] 張明,李盾,陳儀本,等. 羅雪桃1乙酰膽堿酯酶分子生物學研究進展[J]. 農藥,2006,4(1):8-11.
[5] Hayes J D,Flanag an J U,Jow sey I R. Glutathio ne tr ansfer ases[J]. Annu Rev Pharmacol Toxicol,2005,45:51-88.
[6] 郭鳳英,趙志模,鄧新平. 朱砂葉螨抗藥性與羧酸酯酶活性的關系[J]. 農藥學學報,1999,1(2):91-93.
[7] 郭天鳳,史雪巖,高希武,等. 棉蚜吡蟲啉,啶蟲脒不同品系解毒酶活性測定和增效劑作用的研究[J]. 環境昆蟲學報,2014,36(3):388-394.
[8] 宋春滿,鄧建華,吳興富,等. 云南主要煙區煙蚜種群解毒酶活力比較[J]. 昆蟲天敵,2006,28(2):55-61.
[9] 王澤華,石寶才,康總江,等. 甜菜夜蛾不同齡期幼蟲對藥劑的敏感性及其與酶活力的關系[J]. 應用昆蟲學報,2014,51(1):185-193.
[10] 張鵬,李慧,王秋紅,等. 齡期、飼養條件和測定方法對韭菜遲眼蕈蚊藥劑敏感性的影響[J]. 環境昆蟲學報,2014,36(5):730-736.
[11] 劉童童,楊 洪,等. 貴州省煙蚜抗藥性監測.[J]. 環境昆蟲學報2014,36(2):205-212.
[12] Huang Z W,Shi P,Dai J Q,et al. Protein metabolism in Spodopteralitura(F.)is influenced by the botanical insecticide azadirachtin[J].Pesticide Biochemistry and Physiology,2004,80(2):85-93.
[13] Oppenoorth F J. Biochemical genetics of insecticide resistance[J].Annual Review of Entomology,1965,10(1):185-206.
[14] 宋春滿,鄧建華. 云南煙蚜抗藥性機制研究[J]. 環境昆蟲學報,2012,34(3):310-314.
[15] 李周直,沈惠娟,蔣巧根,等. 幾種昆蟲體內保護酶系統活力的研究[J]. 昆蟲學報,1994,37(4):400-403.
[16] Smissaert H R. Cholinesterase Inhibition in Spider Mites Susceptible and Resistant to Organophosphate[J]. Science,1964,143:129-131.
[17] Weill M,Lutfalla G,Mogensen K,et al. Comperative genomics insecticide resistance in mosquito vectors[J]. Nature,2003,423:136-137.
[18] 韓啟發,莊佩君,唐振華. 桃蚜抗藥性的初步研究[J]. 昆蟲學研究集刊(第九集),1988:19-25.
[19] 程新勝,張現. 對藥劑敏感性不同的煙蚜體內幾種關鍵性酶活性的比較分析[J]. 中國煙草學報,2006,12(3):35-38.
[20] 呂敏,劉慧霞,吳君如. 谷胱甘肽S-轉移酶與昆蟲抗藥性的關系[J]. 昆蟲知識,2003,40(3):204-208.
[21] 金 濤,梁廣文,曾玲,等. 不同地理區域桔小實蠅解毒酶系活性及其與抗藥性水平關系[J].環境昆蟲學報,2014,36(1):58-67.
[22] 陳尚文. 馬尾松毛蟲過氧化氫酶及過氧化物酶與耐藥性的關系[J]. 昆蟲學報,2001,44(1):9-14.
[23] 吳小鋒,徐俊良,崔為正,等. 家蠶血液過氧化氫酶活力及其與蠶體抗逆性的關系[J]. 昆蟲學報,1998,41(2):124-129.
[24] 徐欣,霍新北,付榮恕. 不同抗性選育方式下家蠅過氧化氫酶活性及抗性的變化[J]. 中國媒介生物學及控制雜志,2006,17(3):194-197.
(責任編輯 楊賢智)
Relationship between three insecticides resistance and enzyme activity in natural population of Myzus persicae(Sulzer)
ZHANG Chao1,LIU Tong-tong1,2,YANG Hong1,3,WANG Zhao1,4,YANG Mao-fa1,3
(1. Institute of Entomology,Guizhou University,Guiyang 550025,China;
2. Guizhou Academy of Forestry Science,Guiyang 550000,China;
3. College of Tobacco Science, Guizhou University,Guiyang 550025,China;
4. School of Environment and Life Sciences,Kaili College,Kaili 556011,China)
In order to explore the relationship between insecticide resistance and enzyme activity in natural population of Myzus persicae(Sulzer),we studied the activities of Ache,GST and CAT in M. persicae(Sulzer)insecticide resistance to dimethoate,methomyl and avermectins. The results showed that,the resistance stains(different regions)of dimethoate,methomyl and avermectins had higher enzyme activity of AchE,GSTs and CAT than susceptible strain. For the resistance stain of dimethoate,the enzyme activities of AchE in Xingren and Meitan resistant strain were 1.212 times,1.588 times than susceptible strain;the enzyme activities of GSTs resistant strain were 1.625 times,1.900 times than susceptible strain,and the enzyme activities of CAT resistant strain were 1.439 times and 2.018 times than susceptible strain. For the resistance strain of methomyl,the enzyme activities of AchE in Qingzhen and Meitan resistant strain were 1.078 times 1.647 times than susceptible strain;the enzyme activities of GSTs resistant strain were 1.142 times,1.935 times than susceptible strain,and the enzyme activities of CAT resistant strain were 1.142 times,1.935 times than susceptible strain. Forthe resistance strain of avermectins,the enzyme activities of AchE in Xingren,Changshun resistant strain were 1.370 times,1.664 times than susceptible strain;the enzyme activities of GSTs resistant strain were 1.830 times,2.318 times than susceptible strain,and the enzyme activities of CAT resistant strain were 1.800 times,3.708 times than susceptible strain. The results provide a reference for elucidating the field resistance mechanism of M. persicae(Sulzer)population.
Myzus persicae;insecticide resistance;enzyme activity;AChE;CAT;GST
S435.72
A
1004-874X(2017)01-0100-06
2016-10-10
中國煙草總公司貴州省公司科技項目(201022)
張超(1992-),男,在讀碩士生,E-mail:zc0178sfp@163.com
楊洪(1975-),男,博士,教授,E-mail:agr.hyang@gzu.edu.cn
張超,劉童童,楊洪,等. 田間煙蚜種群對3種殺蟲劑抗性與酶活性關系的研究[J].廣東農業科學,2017,44(1):100-105.
