虧水處理條件下玉米氣體交換與水分利用優(yōu)化機制研究
2017-03-21 02:02:36吳立峰李培嶺楊秀霞
節(jié)水灌溉 2017年6期
吳立峰,李培嶺,楊秀霞,燕 輝
(1. 江西農(nóng)業(yè)大學(xué)江西省鄱陽湖流域農(nóng)業(yè)資源與生態(tài)重點實驗室, 南昌 330045;2. 南昌工程學(xué)院水利與生態(tài)工程學(xué)院, 南昌 330099;3. 江西農(nóng)業(yè)大學(xué)工學(xué)院, 南昌 3300451)
水資源是支撐農(nóng)業(yè)生產(chǎn)的重要自然資源,在農(nóng)業(yè)發(fā)展與糧食安全中的作用不可替代。近年來,隨著社會經(jīng)濟的不斷發(fā)展,我國農(nóng)業(yè)用水矛盾日益突出,水資源短缺已成為制約我國農(nóng)業(yè)發(fā)展的重要瓶頸。目前,發(fā)展節(jié)水灌溉農(nóng)業(yè)、提高作物水分利用效率、高效利用有限水資源,已經(jīng)成為保證我國農(nóng)業(yè)可持續(xù)發(fā)展的當(dāng)務(wù)之急。
為建立節(jié)水優(yōu)質(zhì)高效型農(nóng)業(yè)用水模式,實現(xiàn)作物水分利用的優(yōu)化,許多學(xué)者對虧水灌溉條件下作物耗水規(guī)律[1]、產(chǎn)量與品質(zhì)[2]或生理生化特性[3]進行了大量研究,而對虧水處理條件下作物水分利用優(yōu)化機制的研究相對較少。近年研究發(fā)現(xiàn),虧水處理條件下,干燥土壤中的作物根系迅速合成某些化學(xué)物質(zhì),這些作為根信號的化學(xué)物質(zhì)能夠通過木質(zhì)部蒸騰流運輸至冠層,并在葉片水分狀況尚未發(fā)生變化時誘導(dǎo)氣孔關(guān)閉[4]。從而能夠在不犧牲或較少地犧牲光合產(chǎn)量的前提下,最大限度地降低葉片“奢侈”蒸騰,提高水分利用效率。而另一些研究則認為,作物根冠間的這種信息傳遞可能不存在普遍性,多數(shù)情況下的作物葉片氣孔收縮與根信號傳輸不相關(guān)聯(lián)[5]。因此,對虧水處理條件下作物氣孔運動及氣孔運動調(diào)節(jié)下的氣體交換與水分利用機制的研究還有待進一步展開。
玉米是我國重要的旱地作物,具有糧食、工業(yè)原料及飼料等多種用途,在我國工農(nóng)業(yè)生產(chǎn)與糧食安全中的地位不可替代。然而,玉米是高耗水作物,只有在滿足其水分需求的前提下才能獲得高產(chǎn)穩(wěn)產(chǎn)。因此,玉米需水量大與灌溉用水不足的矛盾目前逐漸加劇。為充分挖掘玉米自身的生物節(jié)水潛力,提高其水分利用效率,本研究以玉米為材料,分析土壤逐步干旱條件下其葉片水勢、氣體交換及木質(zhì)部與葉片ABA的變化規(guī)律。以深入了解虧水處理對玉米氣體交換與水分利用的生理調(diào)控機制,為建立節(jié)水、優(yōu)質(zhì)、高效的玉米節(jié)水灌溉模式提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
盆栽試驗在江西農(nóng)業(yè)大學(xué)江西省鄱陽湖流域農(nóng)業(yè)資源與生態(tài)重點實驗室的溫室中進行。2015年4月中旬,將已催芽的玉米種子播種于塑料桶中,每桶播種3粒。桶底均勻布設(shè)5個小孔并鋪有紗網(wǎng)和細砂, 以確保通氣條件良好。玉米出芽后保證充分水分供應(yīng),并在兩葉一心期進行間苗,每桶保留一株長勢均勻的幼苗。
1.2 試驗設(shè)計
試驗于玉米幼苗生長至八葉齡后開始,分別設(shè)置充分供水組與虧水處理組兩種不同的水分處理方式。充分供水組于每日清晨6∶00進行補充灌溉,確保土壤含水量維持在田間持水量(重量)的80%以上,虧水處理組則不加水讓土壤逐漸干旱。試驗進行過程中每天上午對葉片水勢、氣體交換參數(shù)、葉片與木質(zhì)部汁液ABA含量及土壤含水量進行取樣測定,并計算葉片水分利用效率。
1.3 觀測項目和方法
1.3.1 葉水勢測定與木質(zhì)部汁液收集
玉米葉水勢的測定使用Scholander壓力室。切取一段玉米葉片置于壓力室中,并將中脈的切口伸出室外,密封后加壓,當(dāng)木質(zhì)部汁液剛好溢出莖切口后迅速記錄壓力表數(shù)據(jù),并在獲取葉水勢數(shù)據(jù)后繼續(xù)對葉片施壓,直到木質(zhì)部汁液溢出。用移液槍收集汁液并轉(zhuǎn)入離心管,用于測定ABA含量。
1.3.2 氣體交換參數(shù)測定
使用Li-6400型便攜式光合測定儀于上午9∶00-11∶00之間分別測定玉米葉片氣孔導(dǎo)度(gs)、光合速率(Pn)與蒸騰速率(E)。并計算葉片水分利用效率(WUE),WUE=Pn/E。
1.3.3 葉片與ABA含量測定
進行氣體交換參數(shù)測定后,用液氮收獲葉片前端,及時放入-80 ℃超低溫冰箱中儲藏。使用酶聯(lián)免疫法分別測定葉片與木質(zhì)部汁液ABA含量。
1.3.4 土壤含水量測定
土壤含水量的測定采用烘干法。對葉片取樣結(jié)束后,收集桶內(nèi)土壤稱重。然后,將土樣置于105 ℃烘箱中烘至恒重并記錄。土壤含水量依據(jù)公式:土壤含水量=(土重―干土重)/干土重來計算。
1.4 數(shù)據(jù)分析
采用 SPSS 13.0及Origin 7.5分別進行試驗數(shù)據(jù)統(tǒng)計分析并繪圖。
2 結(jié)果及分析
2.1 水分虧缺對土壤含水量及玉米葉水勢的影響
研究表明:在整個試驗過程中,充分供水條件下的土壤含水量維持在24%左右;而虧水處理2 d后,土壤含水量較對照顯著降低;同時,隨著虧水時間的延長,土壤含水量呈現(xiàn)出逐步減少的趨勢,并在虧水處理8 d后達到最小值[圖1(a)]。與土壤含水量的變化趨勢不同,玉米葉片水勢在虧水處理4 d后仍能維持在-0.4 Pa左右的對照水平[圖1(b)]。葉片水勢是描述作物水分狀況的重要指標(biāo),它反映了作物生理活動受環(huán)境水分條件的制約程度。虧水條件下穩(wěn)定的葉水勢有利于維持葉肉細胞膨壓,從而確保葉片各項生理活動的進行;但隨著土壤含水量的進一步降低,葉片水勢在虧水處理5 d后較對照顯著降低[圖1(b)]。葉片水力信號的這種變化,必然導(dǎo)致葉肉細胞膨壓降低,進而對氣孔開度、光合碳同化以及葉片水分利用造成影響。
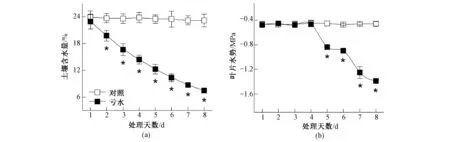
圖1 水分虧缺對土壤含水量及玉米葉水勢的影響
2.2 水分虧缺對玉米葉片氣體交換與水分利用效率的影響
氣孔是葉片與外界進行氣體交換的主要通道,其開放程度直接影響葉片光合與蒸騰作用,從而影響作物的水分利用效率。通過研究水分虧缺條件下玉米氣體交換的變化規(guī)律發(fā)現(xiàn):虧水處理2 d后,葉片氣孔導(dǎo)度與蒸騰速率較對照顯著降低,而光合速率較對照變化不大[圖2(a)~(c)]。這表明光合與蒸騰作用在水分虧缺初期對氣孔收縮的響應(yīng)存在非同步性,蒸騰速率的相對較低將有利于作物在維持光合積累的同時減少葉片“奢侈”蒸騰,從而促進其水分利用效率的提高。這與段愛旺等[6]的研究結(jié)論相一致。隨著土壤含水量的進一步減少,虧水葉片的氣孔導(dǎo)度逐步降低;受氣孔收縮的影響,葉片光合與蒸騰速率在虧水處理4 d后呈現(xiàn)出同步降低的趨勢,并在處理8 d后分別降至最低值[圖2(a)~(c)]。
作物水分利用效率是反映作物耗水與干物質(zhì)積累之間關(guān)系的重要參數(shù),它能夠有效地評價作物在一定環(huán)境條件下的水分利用狀況。通過計算光合與蒸騰的比值發(fā)現(xiàn):虧水處理2 d后,玉米葉片水分利用效率較對照顯著升高,表明玉米在適度水分虧缺條件下消耗單位水分能夠積累更多的干物質(zhì),這也是玉米對水分虧缺的生理適應(yīng)機制。隨著水分虧缺的進一步加劇,玉米水分利用效率在虧水處理4 d后達到最大值,此時氣孔已經(jīng)大幅收縮,但葉片水勢尚未發(fā)生顯著變化。伴隨著土壤含水量的進一步降低,玉米水分利用效率在虧水處理5 d后呈現(xiàn)出下降趨勢,并在虧水處理7 d后降至對照水平[圖2(d)]。

圖2 水分虧缺對玉米葉片氣體交換與水分利用效率的影響
2.3 水分虧缺對玉米木質(zhì)部與葉片ABA含量的影響
脫落酸(ABA)是一種對作物生長發(fā)育、氣孔運動、光合作用、衰老及抗逆性等生理過程都有重要調(diào)節(jié)作用的植物激素。以木本植物相思樹與銀合歡為材料對根源ABA的氣孔調(diào)節(jié)機制進行研究發(fā)現(xiàn):水分虧缺條件下,兩種木本植物根區(qū)合成的ABA能夠通過蒸騰流向上運輸,進而導(dǎo)致木質(zhì)部汁液ABA含量大幅升高[7]。本試驗以玉米為材料進行研究也證實:水分虧缺2 d后,玉米木質(zhì)部汁液ABA含量較對照顯著升高,葉片ABA含量亦呈現(xiàn)出相同的變化趨勢;隨著水分虧缺的進一步加劇,玉米木質(zhì)部汁液與葉片中的ABA含量呈現(xiàn)出同步升高的趨勢,并在虧水處理8 d后分別達到最大值[圖3(a)、(b)]。為進一步明確木質(zhì)部汁液與葉片中ABA含量的關(guān)系,本研究對木質(zhì)部汁液與葉片中ABA含量進行了相關(guān)與回歸分析,結(jié)果表明:木質(zhì)部汁液與葉片中ABA含量呈顯著正相關(guān)(R2=0.946,P<0.05)[圖3(c)]。
2.4 水分虧缺條件下葉片ABA與水勢對氣體交換與水分利用效率的調(diào)控
為進一步明確水分虧缺條件下葉片ABA與水勢對玉米氣體交換與水分利用效率的調(diào)控機制,本研究分別對葉片氣孔導(dǎo)度、光合速率、蒸騰速率及水分利用效率與葉片ABA含量及水勢的關(guān)系進行了相關(guān)及回歸分析,結(jié)果發(fā)現(xiàn):葉片氣孔導(dǎo)度、光合速率、蒸騰速率與ABA含量呈負相關(guān)(R2=0.916,P<0.05;R2=0.926,P<0.05;R2=0.935,P<0.05)[圖4(a)~(c)],而與葉片水勢呈正相關(guān)(R2=0.813,P<0.05;R2=0.911,P<0.05;R2=0.838,P<0.05)[圖5(a)~(c)];此外,葉片水分利用效率與ABA含量呈非線性相關(guān)(R2=0.470,P<0.05)[圖4(d)],而與葉片水勢不相關(guān)(P>0.05)[圖5(d)]。
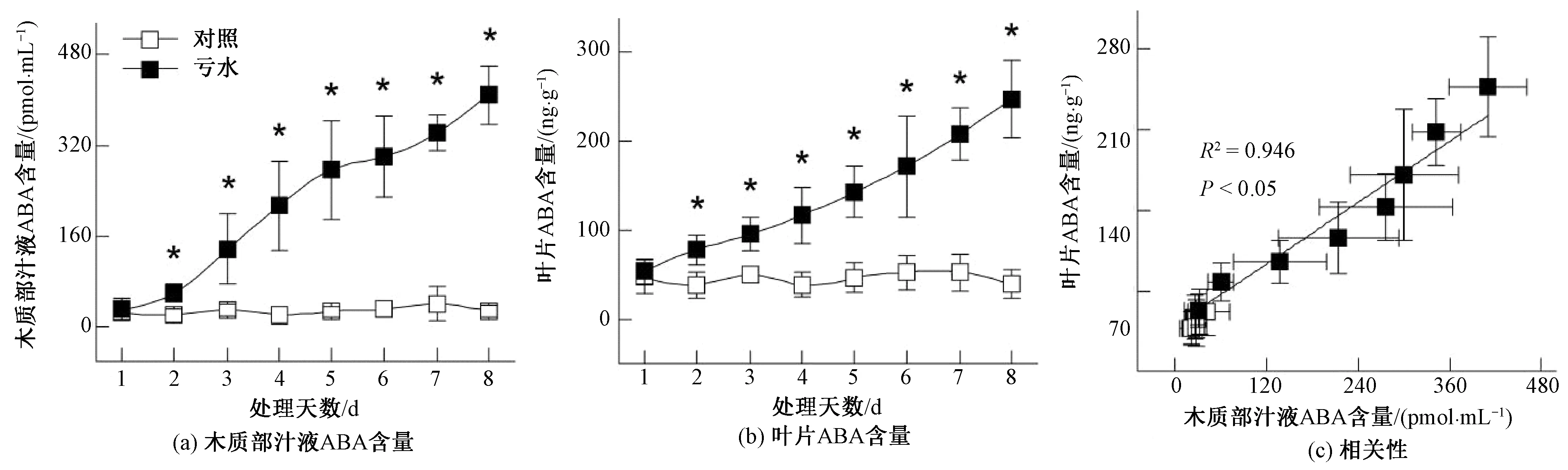
圖3 水分虧缺對玉米木質(zhì)部汁液與葉片ABA含量的影響及兩者ABA含量的相關(guān)性
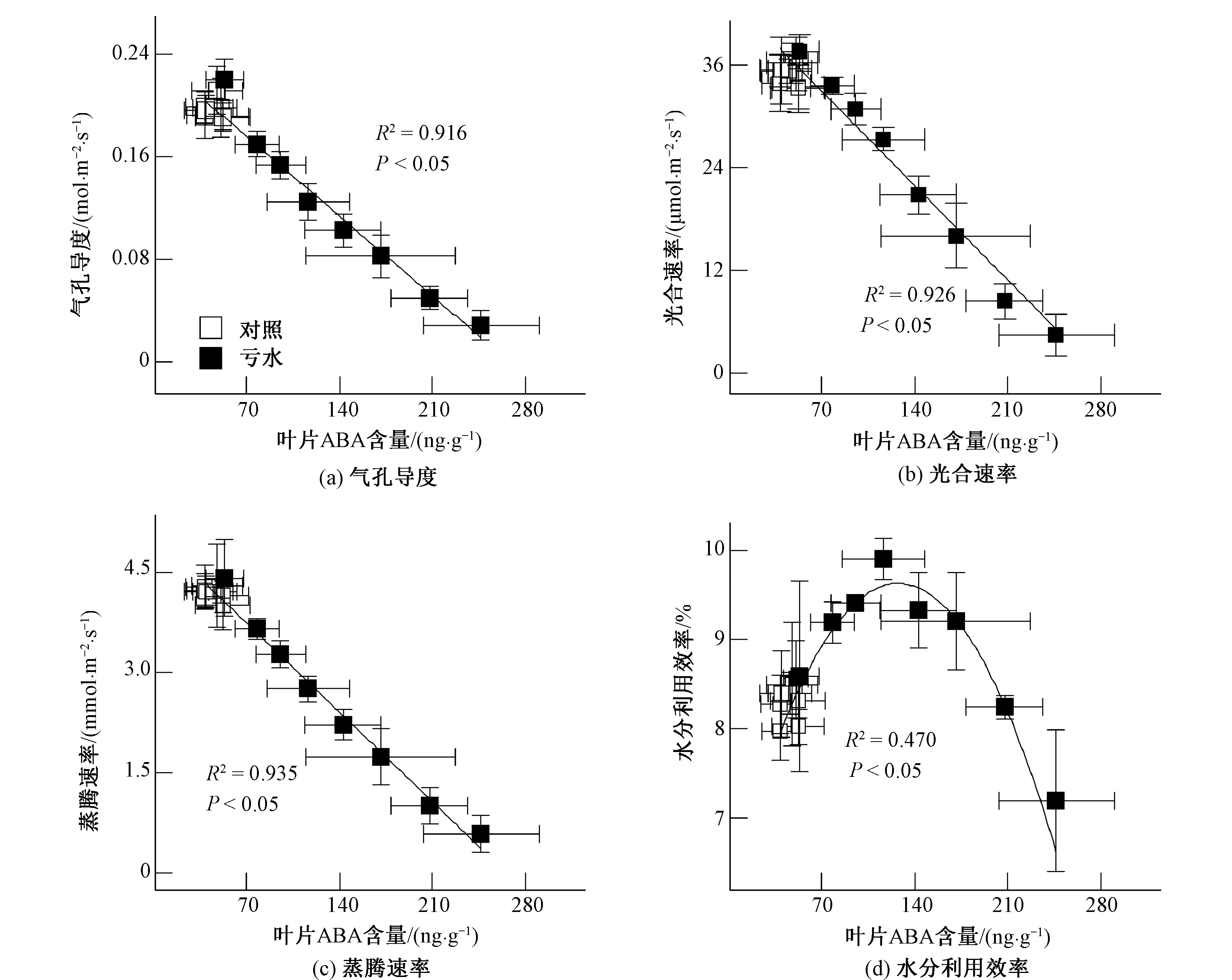
圖4 玉米葉片ABA含量與氣體交換及水分利用效率的相關(guān)及回歸分析
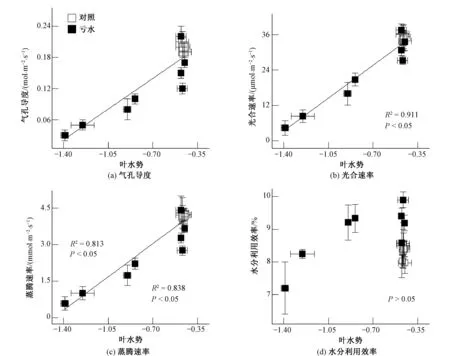
圖5 玉米葉片水勢與氣體交換及水分利用效率的相關(guān)及回歸分析
3 討 論
在水分虧缺誘導(dǎo)的根源化學(xué)信號中,研究最多、最普遍的是內(nèi)源激素ABA。Liu等[8]的研究證實:土壤逐步干燥條件下,馬鈴薯體內(nèi)的根源ABA能夠經(jīng)木質(zhì)部運輸至冠層并有效地調(diào)節(jié)氣孔運動。然而,Holbrook等[9]利用嫁接技術(shù)研究番茄ABA缺乏突變體的氣孔調(diào)節(jié)機制發(fā)現(xiàn),水分虧缺條件下的氣孔關(guān)閉能夠在不依賴于根源ABA的情況下發(fā)生。Munns與King[10]的研究也表明,小麥根信號ABA對氣孔運動的生理調(diào)節(jié)作用不顯著。因此,一些學(xué)者認為,不同作物的根源ABA對氣孔運動的調(diào)節(jié)能力存在差異,某些作物在水分虧缺條件下產(chǎn)生的根源ABA信號能夠有效地誘導(dǎo)氣孔關(guān)閉,而另一些作物產(chǎn)生的根源ABA則對氣孔運動的調(diào)節(jié)作用較弱[11]。本試驗以玉米為材料進行研究發(fā)現(xiàn):隨著土壤水分的虧缺,玉米木質(zhì)部汁液與葉片ABA含量均較對照顯著增加且呈現(xiàn)出逐步升高的趨勢[圖3(a)、(b)];相關(guān)與回歸分析亦證實,兩者呈顯著正相關(guān)(R2=0.946,P<0.05)[圖3(c)]。這表明玉米葉片ABA的積累主要源于根源ABA的木質(zhì)部運輸。同時,水分虧缺條件下,伴隨著玉米葉片ABA含量的顯著升高,葉片氣孔導(dǎo)度呈同步降低趨勢[圖2(a)、圖3(b)],且兩者呈顯著負相關(guān)(R2=0.916;P<0.05)[圖4(a)]。這表明源于根區(qū)的玉米葉片ABA能夠有效地誘導(dǎo)葉片氣孔關(guān)閉。這與Liu等[12]研究大豆氣孔運動機理獲取的結(jié)論相一致。
氣孔是外界CO2與H2O進出細胞的主要通道。因此,根源ABA對氣孔運動的調(diào)控能夠影響葉片的光合與蒸騰作用。這一假設(shè)亦被玉米葉片ABA含量與光合及蒸騰速率呈顯著負相關(guān)(R2=0.926,P<0.05;R2=0.935,P<0.05)[圖4(b)、(c)]的結(jié)論所證實。然而,根源信號ABA在水分脅迫初期誘導(dǎo)的氣孔收縮對葉片光合與蒸騰的影響存在差異:虧水處理2 d后,伴隨著氣孔導(dǎo)度的顯著降低,蒸騰速率較對照顯著降低,而光合速率則變化不大;直到水分虧缺4 d后,光合速率才呈現(xiàn)出顯著降低的趨勢[圖2(a)~(c)]。這表明虧水條件下的光合對氣孔收縮的反應(yīng)較蒸騰滯后。正是由于這種光合與蒸騰速率響應(yīng)水分虧缺的不同步,導(dǎo)致玉米葉片水分利用效率(WUE=Pn/E)在水分脅迫初期較對照顯著升高[圖2(d)]。回歸分析進一步證實,玉米葉片水分利用效率隨根源ABA的積累呈非線性相關(guān)(R2=0.470,P<0.05)[圖4(d)],表明根源ABA調(diào)控下的氣孔運動在提高玉米水分利用效率過程中發(fā)揮著重要作用。
除根信號ABA外,葉片水勢也能夠有效地調(diào)控氣孔行為。紀文龍等[13]研究干旱脅迫下葡萄葉片氣孔導(dǎo)度和水勢動態(tài)的變化規(guī)律發(fā)現(xiàn):在土壤水分快速虧缺的情況下,葡萄葉水勢在調(diào)控氣孔運動過程中起決定性作用。Alexander Christmann等[14]也認為,水力信號在調(diào)控氣孔運動過程中起重要作用。本研究發(fā)現(xiàn):水分虧缺條件下,玉米葉片水勢與氣孔導(dǎo)度(R2=0.813,P<0.05)及氣孔運動調(diào)控下的光合速率(R2=0.911,P<0.05)與蒸騰速率(R2=0.838,P<0.05)呈顯著正相關(guān)[圖5(a)~(c)]。這與葉片水勢降低能夠直接誘導(dǎo)氣孔收縮有關(guān)系。本研究中玉米葉片水勢變化與水分利用效率不相關(guān)(P>0.05)[圖5(d)]則證實葉片化學(xué)信號ABA積累而非葉水勢變化在水分虧缺過程中起到了提高玉米水分利用效率的作用。
4 結(jié) 論
(1)水分虧缺能夠造成玉米體內(nèi)化學(xué)信號ABA的木質(zhì)部傳輸與葉片積累,葉片積累的ABA誘導(dǎo)了葉水勢降低之前的氣孔收縮。
(2)由于光合速率與蒸騰速率在水分脅迫初期對氣孔響應(yīng)的非同步性,水分虧缺條件下的蒸騰速率能夠隨氣孔收縮顯著降低,但光合速率在虧水初期變化不大。從而造成了虧水初期的玉米葉片水分利用效率在ABA誘導(dǎo)的氣孔收縮條件下呈現(xiàn)出逐步升高的趨勢。
(3)通過輕度虧水處理能夠有效地創(chuàng)造玉米高效用水環(huán)境,提高玉米自身生物節(jié)水潛力,從而達到提高其水分利用效率的目的。但鑒于本研究為針對一次試驗結(jié)果的分析,因此,對玉米水分高效利用機制的研究還有待進一步深入。
[1] 黃海霞, 韓國君, 陳年來, 等. 荒漠綠洲調(diào)虧灌溉條件下辣椒耗水規(guī)律研究[J]. 自然資源學(xué)報, 2012,27(5):747-756.
[2] 胡志橋, 馬忠明, 包興國, 等.虧缺灌溉對石羊河流域主要作物產(chǎn)量和耗水量的影響[J]. 節(jié)水灌溉, 2010,(7):10-13.
[3] 李 蕊,張寬地,陳俊英. 無壓灌溉條件下不同滴灌施氮量對櫻桃生長發(fā)育及產(chǎn)量的影響[J]. 節(jié)水灌溉, 2016,(1):23-26.
[4] Jiang F, Hartung W. Long-distance signalling of abscisic acid (ABA): the factors regulating the intensity of the ABA signal[J]. Journal of Experimental Botany, 2008,59(1):37-43.
[5] Kramer P J. Changing concepts regarding plant water relations[J]. Plant, Cell and Environment, 1988,(11):565-568.
[6] 段愛旺, 肖俊夫, 張寄陽, 等. 控制交替溝灌中灌水控制下限對玉米葉片水分利用效率的影響[J]. 作物學(xué)報, 1999,25(6):766-771.
[7] Liang J, Zhang J, Wong M H. Can stomatal closure caused by xylem ABA explain the inhibition of leaf photosynthesis under soil drying?[J]. Photosynthesis Research, 1997,51(2):149-159.
[8] Liu F, Jensen C R, Shahanzari A, et al. ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying[J]. Plant Science, 2005,168(3):831-836.
[9] Holbrook N M, Shashidhar V R, James R A, et al. Stomatal control in tomato with ABA-deficient roots: response of grafted plants to soil drying[J]. Journal of Experimental Botany, 2002,53(373):1 503-1 514.
[10] Munns R, King R W. Abscisic acid is not the only stomatal inhibitor in the transpiration stream of wheat plants[J]. Plant Physiology, 1988,88(3):703-708.
[11] 王晶晶, 莫偉平, 賈文鎖, 等. 干旱條件下葡萄葉片氣孔導(dǎo)度和水勢與節(jié)位變化的關(guān)系[J]. 中國農(nóng)業(yè)科學(xué), 2013,46(10):2 151-2 158.
[12] Liu F, Andersen M N, Jacobsen S E, et al. Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying[J]. Environmental and Experimental Botany, 2005, 54(1): 33-40.
[13] 紀文龍,范意娟,李 辰,等. 干旱脅迫下葡萄葉片氣孔導(dǎo)度和水勢動態(tài)的變化規(guī)律[J]. 中國農(nóng)業(yè)大學(xué)學(xué)報, 2014,19(4):74-80.
[14] Christmann A, Weiler E W, Steudle E, et al. A hydraulic signal in root-to-shoot signalling of water shortage[J]. The Plant Journal, 2007,52(1):167-174.
