云南松花粉儲(chǔ)藏溫度及離體萌發(fā)條件
2017-02-20 03:43:55周蘭英
浙江農(nóng)林大學(xué)學(xué)報(bào) 2017年1期
李 梅,周蘭英
(四川農(nóng)業(yè)大學(xué) 林學(xué)院,四川 成都611130)
云南松花粉儲(chǔ)藏溫度及離體萌發(fā)條件
李 梅,周蘭英
(四川農(nóng)業(yè)大學(xué) 林學(xué)院,四川 成都611130)
以云南松Pinus yunnanesis花粉為材料,通過設(shè)置3種儲(chǔ)藏溫度(25℃,5℃,-20℃)探討花粉生活力隨時(shí)間的變化規(guī)律;通過單因子實(shí)驗(yàn)和正交實(shí)驗(yàn),探究不同質(zhì)量濃度的蔗糖、硼酸、赤霉素、氯化鈣單獨(dú)或共同作用對(duì)花粉萌發(fā)的影響。結(jié)果表明:云南松花粉儲(chǔ)藏0 d時(shí)花粉生活力為91.77%,隨著儲(chǔ)藏時(shí)間的增加3種儲(chǔ)藏溫度的花粉生活力均下降,下降速率表現(xiàn)為25℃>5℃>-20℃,說明低溫能增加花粉的耐儲(chǔ)性。單因子萌發(fā)實(shí)驗(yàn)顯示:蔗糖的最適質(zhì)量濃度為80 g·L-1,萌發(fā)率為82.41%;赤霉素的最適質(zhì)量濃度為100 mg·L-1,萌發(fā)率為66.41%;硼酸的最適質(zhì)量濃度為300 mg·L-1,萌發(fā)率為18.23%;氯化鈣的最適質(zhì)量濃度為100 mg·L-1,萌發(fā)率為56.98%。正交實(shí)驗(yàn)顯示:4種因素的最佳組合為120 g·L-1蔗糖+150 mg·L-1赤霉素+100 mg·L-1氯化鈣+100 mg·L-1硼酸,萌發(fā)率可達(dá)89.74%。圖3表2參11
林木育種學(xué);云南松;花粉;儲(chǔ)藏溫度;生活力;萌發(fā)
云南松Pinus yunnanesis是松科Pinaceae松屬Pinus常綠針葉喬木,生長(zhǎng)快,材質(zhì)好,耐干旱瘠薄,天然更新能力強(qiáng),是西南地區(qū)荒山造林先鋒樹種和主要的用材樹種[1],具有很高的經(jīng)濟(jì)開發(fā)價(jià)值。然而,因遺傳改良滯后,云南松優(yōu)質(zhì)高效人工林的發(fā)展受到嚴(yán)重制約。有研究[2]表明,云南松樹高、通直度、結(jié)實(shí)量等性狀受基因連鎖或功效基因的作用較大,受環(huán)境的影響較小,有較高的遺傳穩(wěn)定性。同時(shí),群體內(nèi)存在許多生長(zhǎng)、形質(zhì)和材性兼優(yōu)的家系、個(gè)體,尤其是生長(zhǎng)性狀的遺傳變異潛力巨大,只要通過有效的良種選育就能發(fā)掘這些優(yōu)良基因型[3]。花粉研究是研究遺傳改良的重要內(nèi)容,對(duì)于云南松花粉,除了花粉形態(tài)[4]、花粉成分[5]研究外,尚未見有關(guān)花粉生活力方面的研究報(bào)道。本研究通過對(duì)云南松花粉儲(chǔ)藏溫度及萌發(fā)條件的探究,旨在為云南松雜交育種工作的順利開展提供技術(shù)依據(jù)。
1 材料與方法
1.1 材料
云南松花粉采自四川省會(huì)理縣太平鎮(zhèn)云南松人工林。
1.2 方法
1.2.1 花粉收集 2014年4月下旬采集成熟的云南松雄球花,置于光滑紙面上,在室內(nèi)攤晾收集,花粉混合均勻后裝入牛皮紙袋中帶回實(shí)驗(yàn)室。
1.2.2 儲(chǔ)藏溫度和儲(chǔ)藏時(shí)間對(duì)云南松花粉生活力的影響 將花粉分裝在清潔干燥帶瓶蓋的青霉素小瓶中,標(biāo)記,分別置于25℃,5℃,-20℃條件下儲(chǔ)藏。儲(chǔ)藏初期隔24 h測(cè)定3種儲(chǔ)藏溫度下花粉的生活力,后期可根據(jù)情況延長(zhǎng)測(cè)定間隔時(shí)間,直到花粉失去生活力(花粉生活力低于50%)。
1.2.3 花粉生活力測(cè)定 采用碘-碘化鉀染色法、α-萘酚-聯(lián)苯胺法、次甲基藍(lán)染色法等3種方法測(cè)定花粉生活力,篩選出能快速準(zhǔn)確測(cè)定花粉生活力的方法。測(cè)定時(shí),先將低溫條件下的云南松花粉在常溫下放置0.5 h,再用染色劑處理,重復(fù)3個(gè)·處理-1,觀察清晰視野5個(gè)·重復(fù)-1,統(tǒng)計(jì)花粉粒數(shù)≥30粒·視野-1,統(tǒng)計(jì)各視野花粉生活力,并計(jì)算各處理平均生活力。花粉生活力(%)=(染色花粉數(shù)/統(tǒng)計(jì)花粉總數(shù))×100%。
1.2.4 花粉萌發(fā)條件的測(cè)定 設(shè)置培養(yǎng)液蔗糖質(zhì)量濃度為0,50,80,100,120 g·L-1;硼酸質(zhì)量濃度為0,100,300,500 mg·L-1; 赤霉素質(zhì)量濃度為 0,50, 100,150 mg·L-1; 氯化鈣質(zhì)量濃度為0,50,100,150 mg·L-1。通過單因素實(shí)驗(yàn),篩選出培養(yǎng)液各因素的最適質(zhì)量濃度,再進(jìn)行正交實(shí)驗(yàn)設(shè)計(jì),篩選出培養(yǎng)液各因素最佳質(zhì)量濃度組合。花粉采用液體培養(yǎng)法進(jìn)行培養(yǎng),先將解剖針和雙凹片干燥滅菌,處理時(shí)用解剖針蘸取少量花粉散播在載有不同培養(yǎng)液的雙凹片凹孔中,蓋上蓋玻片,置于25℃LHP-300H智能型人工氣候箱中黑暗培養(yǎng)。3 h在顯微鏡下觀測(cè)1次,重復(fù)3次·處理-1,觀察清晰視野5個(gè)·重復(fù)-1,統(tǒng)計(jì)花粉粒數(shù)≥30粒·視野-1,當(dāng)花粉管長(zhǎng)度≥花粉直徑時(shí)視為花粉萌發(fā),觀察至萌發(fā)率不再增加為止。統(tǒng)計(jì)各視野花粉生活力,并計(jì)算各處理平均生活力。花粉萌發(fā)率(%)=(花粉萌發(fā)數(shù)/統(tǒng)計(jì)花粉總數(shù))×100%。
1.2.5 數(shù)據(jù)處理 所得數(shù)據(jù)用Excel,SPSS 20進(jìn)行統(tǒng)計(jì)分析。進(jìn)行方差分析時(shí),需對(duì)原始數(shù)據(jù)進(jìn)行反正弦轉(zhuǎn)換。
2 結(jié)果與分析
2.1 不同儲(chǔ)藏溫度和儲(chǔ)藏時(shí)間下花粉生活力變化的比較
a-萘酚-聯(lián)苯胺法染色不明顯,次甲基藍(lán)染色法在進(jìn)行染色梯度界定時(shí)無(wú)固定標(biāo)準(zhǔn),難統(tǒng)一,而經(jīng)碘-碘化鉀染色法測(cè)定結(jié)果穩(wěn)定、易辨別、最接近萌發(fā)實(shí)驗(yàn)結(jié)果,因此,本研究以碘-碘化鉀染色法測(cè)定結(jié)果作為論述依據(jù)。測(cè)定結(jié)果(圖1)顯示:隨著儲(chǔ)藏天數(shù)的增加,3種儲(chǔ)藏溫度下云南松花粉生活力均下降,但儲(chǔ)藏溫度不同,花粉生活力下降的速率不同,其中25℃條件下花粉生活力下降較快,5℃條件和-20℃條件下花粉生活力下降趨勢(shì)都較為平緩,總的花粉生活力下降趨勢(shì)表現(xiàn)為25℃>5℃>-20℃。方差分析結(jié)果表明:儲(chǔ)藏時(shí)間和儲(chǔ)藏溫度對(duì)花粉生活力的影響達(dá)極顯著水平(F儲(chǔ)藏時(shí)間=35.482>F0.01;F儲(chǔ)藏溫度= 30.404>F0.01),且儲(chǔ)藏溫度的F值較儲(chǔ)藏時(shí)間的F值大,說明儲(chǔ)藏溫度比儲(chǔ)藏時(shí)間對(duì)花粉生活力的影響大。多重比較顯示(表1):3種儲(chǔ)藏溫度對(duì)云南松花粉生活力影響顯著,均數(shù)大小表現(xiàn)為-20℃>5℃>25℃,即表明-20℃條件對(duì)花粉生活力的保持顯著優(yōu)于5℃條件和25℃條件,5℃條件顯著優(yōu)于25℃條件。
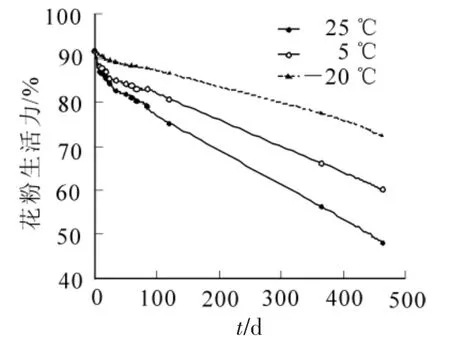
圖1 不同儲(chǔ)藏溫度下花粉生活力的比較Figure 1 Comparison ofpollen viability under different storage conditions
2.2 花粉萌發(fā)
2.2.1 花粉萌發(fā)時(shí)間和過程 云南松花粉萌發(fā)過程見圖2。云南松花粉在培養(yǎng)24 h之前不萌發(fā),培養(yǎng)24~72 h,萌發(fā)率急劇上升,72 h后萌發(fā)率趨于平穩(wěn),超過72 h以后新增花粉萌發(fā)數(shù)極少,可見,72 h后即可統(tǒng)計(jì)花粉萌發(fā)率。花粉粒萌動(dòng)時(shí)萌發(fā)溝薄壁區(qū)呈透明狀態(tài)并向兩邊突起,繼而其中一端或兩端伸長(zhǎng)成為花粉管,花粉管生長(zhǎng)后期會(huì)分支,甚至破裂。
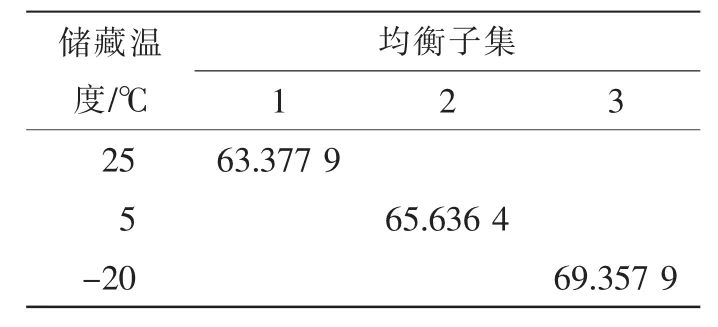
表1 均衡子集表Table 1 Balanced subsets table
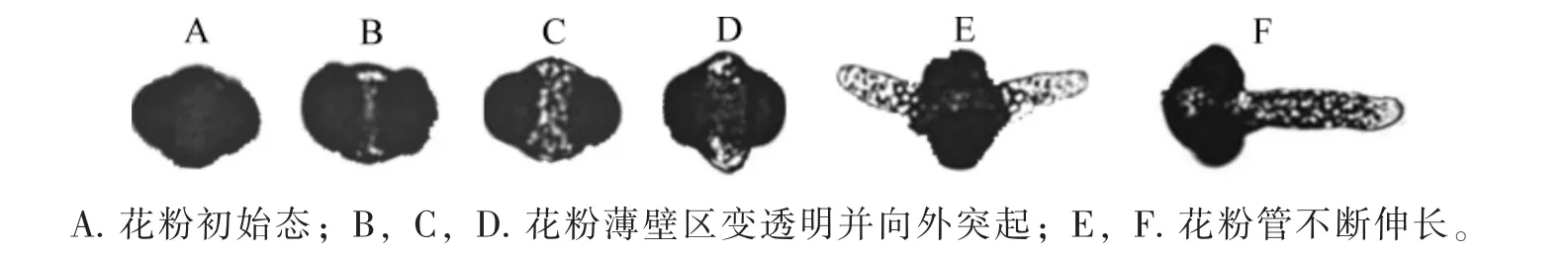
圖2 云南松花粉萌發(fā)過程Figure 2 Mination process of Pinus yunnanesis pollen
2.2.2 蔗糖、氯化鈣、硼酸、赤霉素單因子作用對(duì)云南松花粉萌發(fā)的影響 由圖3A可知:蔗糖對(duì)云南松花粉萌發(fā)存在極顯著影響。花粉在清水中也能萌發(fā),平均萌發(fā)率為9.02%,隨著蔗糖質(zhì)量濃度的增加,花粉萌發(fā)率呈先升高后降低的趨勢(shì),在蔗糖質(zhì)量濃度為80 g·L-1時(shí)最高,為82.41%。蔗糖質(zhì)量濃度超過80 g·L-1后花粉萌發(fā)率降低,可見最適蔗糖質(zhì)量濃度為80 g·L-1。由圖3B可知:不同質(zhì)量濃度的氯化鈣對(duì)云南松花粉萌發(fā)率影響程度不同。當(dāng)培養(yǎng)基氯化鈣質(zhì)量濃度為 50 mg·L-1時(shí),萌發(fā)率為13.06%;隨氯化鈣質(zhì)量濃度的增加花粉萌發(fā)率逐漸升高,當(dāng)氯化鈣質(zhì)量濃度升至100 mg·L-1時(shí),萌發(fā)率達(dá)到最高,為56.98%;但隨著氯化鈣質(zhì)量濃度繼續(xù)增加,花粉萌發(fā)率下降。可見云南松花粉萌發(fā)的適宜氯化鈣質(zhì)量濃度是100 mg·L-1。由圖3C可知:硼酸質(zhì)量濃度為300 mg·L-1時(shí),花粉萌發(fā)率達(dá)到最大值18.23%,極顯著高于對(duì)照和100 mg·L-1;而質(zhì)量濃度為500 mg·L-1時(shí),花粉萌發(fā)受到強(qiáng)烈抑制,萌發(fā)率僅為1.35%,極顯著低于清水處理。結(jié)果表明:云南松花粉萌發(fā)的較適宜硼酸質(zhì)量濃度為300 mg· L-1。由圖3D可知:當(dāng)培養(yǎng)基赤霉素質(zhì)量濃度為100 mg·L-1時(shí),花粉萌發(fā)率達(dá)到最大值66.41%,極顯著高于其他處理。蔗糖、氯化鈣、硼酸、赤霉素等單因子對(duì)云南松花粉的萌發(fā)都存在極顯著影響,添加后大部分萌發(fā)率比對(duì)照有顯著增長(zhǎng),以蔗糖作用最明顯。單因子實(shí)驗(yàn)結(jié)果顯示最適濃度組合為80 g·L-1蔗糖+100 mg·L-1赤霉素+100 mg·L-1氯化鈣+300 mg·L-1硼酸。
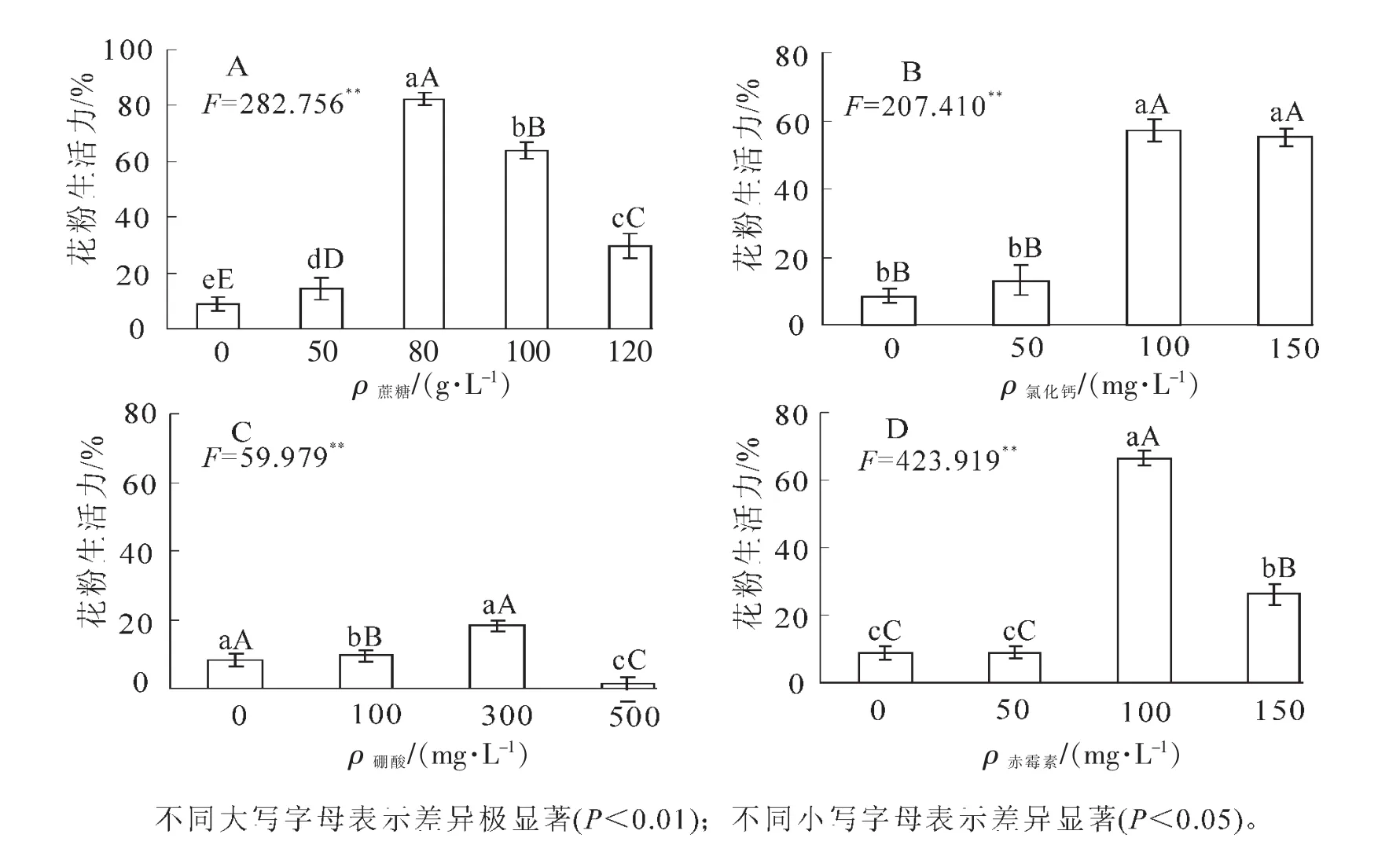
圖3 蔗糖、氯化鈣、硼酸、赤霉素對(duì)云南松花粉萌發(fā)的影響Figure 3 Effect of sucrose,CaCl2,H3BO3and GA3on in vitro pollen germinetion of Pinus yunnanesis
2.2.3 蔗糖、氯化鈣、硼酸、赤霉素等4因子正交實(shí)驗(yàn)對(duì)云南松花粉萌發(fā)率的影響 表2結(jié)果表明:蔗糖、硼酸、赤霉素、氯化鈣在一定質(zhì)量濃度范圍內(nèi)共同作用能明顯提高云南松花粉的萌發(fā)率。實(shí)驗(yàn)中各組處理對(duì)花粉萌發(fā)率的影響與對(duì)照組都存在極顯著差異,其中處理1萌發(fā)率最高,達(dá)89.74%;其次為處理2,達(dá)77.65%。正交實(shí)驗(yàn)結(jié)果顯示最優(yōu)搭配為120 g·L-1蔗糖+150 mg·L-1赤霉素+100 mg·L-1氯化鈣+100 mg·L-1硼酸,與單因子實(shí)驗(yàn)所得結(jié)果有差異,這說明各種營(yíng)養(yǎng)因子對(duì)花粉萌發(fā)的影響是相互作用的。
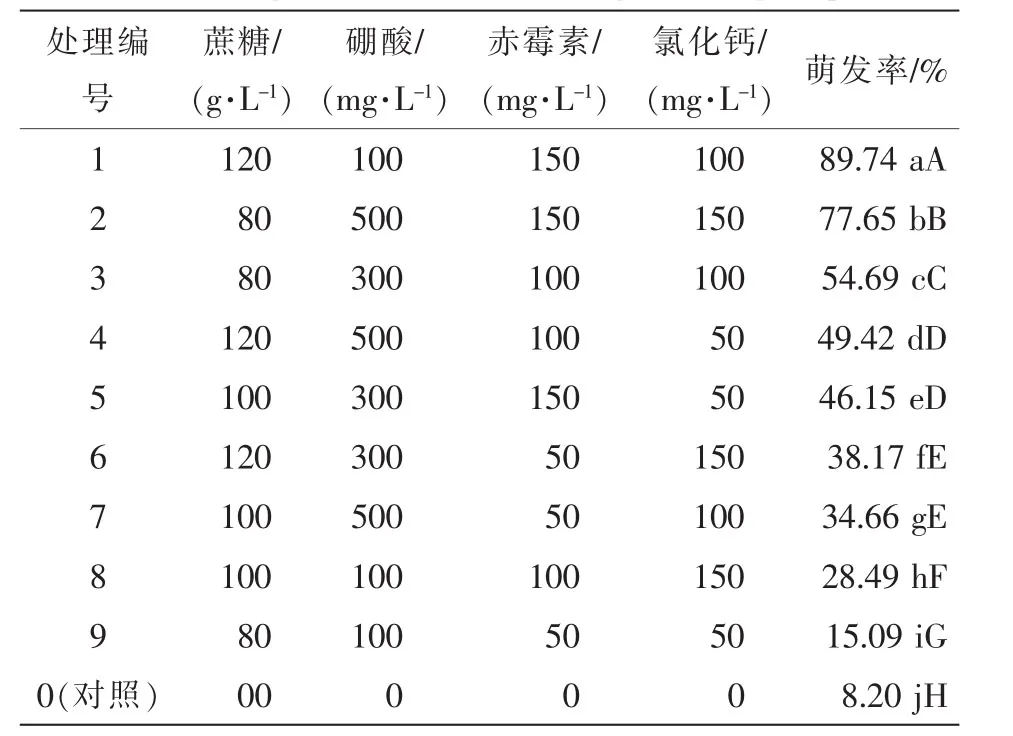
表2 花粉萌發(fā)L9(34)正交實(shí)驗(yàn)結(jié)果Table 2 Pollen germination rate in orthogonal design experiment
3 結(jié)論與討論
3.1 花粉生活力
研究表明:碘-碘化鉀染色法能反應(yīng)不同儲(chǔ)藏溫度下花粉生活力的變化趨勢(shì)。染色法測(cè)定的花粉生活力略高于培養(yǎng)基法,這是因?yàn)槿旧珪r(shí)染色劑也能對(duì)未成熟、衰老和敗育的花粉進(jìn)行染色,而這類花粉不一定都具有受精能力,以致測(cè)定結(jié)果往往偏高。雖然培養(yǎng)基法是最能準(zhǔn)確測(cè)定花粉生活力的方法,但是其耗時(shí)長(zhǎng),操作過程相對(duì)復(fù)雜,碘-碘化鉀染色法能快速準(zhǔn)確地測(cè)定云南松花粉的生活力。
報(bào)道顯示[6-8]:銀杉Cathaya argyrophylla等裸子植物的鮮花粉生活力均在90%以上。本研究通過碘-碘化鉀染色法測(cè)定云南松鮮花粉的生活力為91.77%,與以上裸子植物結(jié)果相一致,都具有較高的鮮花粉生活力。本研究還表明:云南松花粉具有較高的耐儲(chǔ)藏性,在25℃條件下儲(chǔ)藏463 d時(shí)云南松花粉生活力為48.21%,在5℃條件下儲(chǔ)藏463 d時(shí)為60.28%,在-20℃條件下463 d時(shí)為72.34%。儲(chǔ)藏時(shí)間和儲(chǔ)藏溫度對(duì)于花粉生活力都有極顯著的影響,但儲(chǔ)藏溫度影響更大。這是因?yàn)榈蜏啬芙档突ǚ酆粑鼜?qiáng)度,減少自身營(yíng)養(yǎng)物質(zhì)的消耗。
3.2 花粉萌發(fā)條件
蔗糖是許多植物花粉離體培養(yǎng)所必需的營(yíng)養(yǎng)成分,一般認(rèn)為蔗糖為花粉的萌發(fā)和花粉管的生長(zhǎng)提供了能源和碳源,同時(shí)還能調(diào)節(jié)培養(yǎng)環(huán)境滲透壓。蔗糖適宜質(zhì)量濃度通常為100~150 g·L-1,云南松花粉萌發(fā)的最適宜蔗糖質(zhì)量濃度為80 g·L-1,且總體情況是低質(zhì)量濃度比高質(zhì)量濃度對(duì)花粉萌發(fā)更有利。其原因可能是高質(zhì)量濃度蔗糖造成花粉細(xì)胞質(zhì)壁分離,使花粉粒脫水,從而影響花粉的萌發(fā)。鈣離子是花粉管生長(zhǎng)的基本因素。本研究中單因子氯化鈣質(zhì)量濃度對(duì)花粉萌發(fā)的影響極顯著,低質(zhì)量濃度時(shí)對(duì)花粉萌發(fā)率的影響不大,質(zhì)量濃度增加能顯著提高花粉萌發(fā)率,100 mg·L-1時(shí)萌發(fā)率達(dá)最大值56.98%。可能云南松花粉自身含有較少的鈣離子,萌發(fā)時(shí)外源鈣離子的適量補(bǔ)充可以促進(jìn)其萌發(fā)。硼不僅可以增加糖的吸收、運(yùn)轉(zhuǎn)和代謝,形成花粉管頂端生長(zhǎng)依賴的梯度,還可能作為一種相關(guān)因子影響關(guān)鍵酶活性,改變細(xì)胞壁延展性以至影響花粉的萌發(fā)、花粉管細(xì)胞壁的構(gòu)建和花粉管的伸長(zhǎng)[9]。一般認(rèn)為,花粉內(nèi)存在較多的鈣,而缺乏硼,因硼的缺乏往往要從柱頭和花柱內(nèi)獲得硼的補(bǔ)償[10]。本研究中,云南松花粉萌發(fā)對(duì)硼需要量十分微小,在最適質(zhì)量濃度300 mg·L-1時(shí)萌發(fā)率僅為18.23%,500 mg·L-1時(shí)即表現(xiàn)為強(qiáng)烈的抑制作用,可能花粉內(nèi)含有較多的硼元素,能在一定程度上滿足花粉萌發(fā)所需,但硼適量的補(bǔ)充也能提高花粉萌發(fā)率。雖然目前有關(guān)赤霉素等植物生長(zhǎng)調(diào)節(jié)物質(zhì)能促進(jìn)花粉的萌發(fā)和花粉管生長(zhǎng)的機(jī)制尚不清楚,但已有研究表明花粉自身富含生長(zhǎng)素及赤霉素等生長(zhǎng)調(diào)節(jié)物質(zhì),這些物質(zhì)的多少對(duì)花粉自身的萌發(fā)、生長(zhǎng)及儲(chǔ)藏壽命均有較大的影響,對(duì)花粉管在花柱內(nèi)生長(zhǎng)完成受精及促進(jìn)果實(shí)膨大也有重要的作用,因此,一定質(zhì)量濃度的外源赤霉素處理能促進(jìn)花粉萌發(fā)和花粉管生長(zhǎng)[11]。在本研究中赤霉素對(duì)花粉萌發(fā)率存在極顯著性影響,以100 mg·L-1最佳,萌發(fā)率可達(dá)66.41%。總體來說,云南松花粉生活力強(qiáng),對(duì)培養(yǎng)基營(yíng)養(yǎng)物質(zhì)的需求量不高,單因子以蔗糖作用最明顯,正交實(shí)驗(yàn)最適的液體培養(yǎng)基組合為120 g· L-1+150 mg·L-1赤霉素+100 mg·L-1氯化鈣+100 mg·L-1硼酸。這可能是因?yàn)樵颇纤苫ǚ壑凶陨砗械哪芰课镔|(zhì)較多,適量補(bǔ)充能更好地滿足萌發(fā)所需營(yíng)養(yǎng)物質(zhì),高質(zhì)量濃度的營(yíng)養(yǎng)物質(zhì)補(bǔ)充反而會(huì)抑制花粉的正常萌發(fā)。
在雜交授粉實(shí)際操作中,一般采用生活力不低于新鮮花粉生活力的50%的花粉進(jìn)行授粉,因此,云南松花粉在25℃條件下儲(chǔ)藏1 a之內(nèi)可用于授粉,低溫儲(chǔ)藏能有效延緩花粉生活力下降的速度。在授粉時(shí)可以噴施一定質(zhì)量濃度的蔗糖、赤霉素,促進(jìn)花粉萌發(fā)和花粉管伸長(zhǎng),進(jìn)而提高雜交育種成效。
[1] 黃瑞復(fù).云南松的種群遺傳與進(jìn)化[J].云南大學(xué)學(xué)報(bào)(自然科學(xué)版),1993,15(1):50-63.
HUANG Ruifu.The population genetics and evolution of Pinus yunnanensis[J].J Yunnan Univ Nat Sci,1993,15(1): 50-63.
[2] 陳強(qiáng),常恩福,董福美,等.云南松天然優(yōu)良林分疏伐營(yíng)建母樹林的研究[J].云南林業(yè)科技,2000(3):1-8.
CHEN Qiang,CHANG Enfu,DONG Fumei,et al.Study on establishment of seed production stand by thinning natural superior stands of Pinus yunnanensis[J].J Yunnan For Sci Technol,2000(3):1-8.
[3] 張躍敏,李根前,李蓮芳,等.云南松生長(zhǎng)特性及其促成培育[J].陜西林業(yè)科技,2008(3):4-7.
ZHANG Yuemin,LI Genqian,LI Lianfang,et al.The growth characteristics of Pinus yunnanensis and improvement measures[J].J Shaanxi For Sci Technol,2008(3):4-7.
[4] 楊彩云,楊貌仙.云南松花粉形態(tài)研究[J].云南植物研究,1989,11(1):71-74.
YANG Caiyun,YANG Maoxian.Study on the pollen morphology of Pinus yunnanensis[J].Acta Bot Yunnan,1989,11(1):71-74.
[5] 范國(guó)棟,劉嘉寶,馮武.破壁云南松花粉成分分析及其安全性和功能評(píng)價(jià)[J].云南林業(yè)科技,2001(1):54-57.
FAN Guodong,LIU Jiabao,FENG Wu.Evaluation on ingredients,security and function of coat-broken Pinus yunnanensis pollen[J].J Yunnan For Sci Technol,2001(1):54-57.
[6] 王紅衛(wèi),鄧輝勝,譚海明,等.銀杉花粉生命力及其變異[J].植物生態(tài)學(xué)報(bào),2007,31(6):1199-1204.
WANG Hongwei,DENG Huisheng,TAN Haiming,et al.Pollen viability and variation in Cathaya argyrophylla[J].J Plant Ecol,2007,31(6):1199-1204.
[7] 馬建偉,王軍輝,張守攻,等.云杉花粉的貯藏及萌發(fā)研究[J].林業(yè)科學(xué)研究,2012,25(3):302-307.
MA Jianwei,WANG Junhui,ZHANG Shougong,et al.Study on storage and bourgeoning of spruce pollen[J].For Res, 2012,25(3):302-307.
[8] 孫蕾,房用,馬玲,等.黑松和赤松花粉取粉處理和干燥貯藏技術(shù)研究[J].食品科學(xué),2007,28(7):189-191.
SUN Lei,FANG Yong,MA Ling,et al.Study on production and storage technology of black pines and Japanese red pines pollens[J].Food Sci,2007,28(7):189-191.
[9] 龔明,曹宗巽.鈣和鈣調(diào)素對(duì)花粉萌發(fā)和花粉管生長(zhǎng)的調(diào)控[J].植物生理學(xué)通訊,1995,31(5):321-328.
GONG Ming,CAO Zongxun.Regulation of calcium and calmodulin on pollen germination and growth of pollen tube[J].Plant Physiol Commun,1995,31(5):321-328.
[10] 張紹鈴,陳迪新,康瑯,等.培養(yǎng)基組分及pH值對(duì)梨花粉萌發(fā)和花粉管生長(zhǎng)的影響[J].西北植物學(xué)報(bào),2005,25(2):225-230.
ZHANG Shaoling,CHEN Dixin,KANG Lang,et al.Effects of medium components and pH on pollen germination and tube growth in pear(Pyrus pyrifolia)[J].Acta Bot Boreal-Occident Sin,2005,25(2):225-230.
[11] 張紹鈴,高付永,陳迪新,等.植物生長(zhǎng)調(diào)節(jié)物質(zhì)對(duì)豐水梨花粉萌發(fā)和花粉管生長(zhǎng)的影響[J].西北植物學(xué)報(bào),2003,23(4):586-591.
ZHANG Shaoling,GAO Fuyong,CHEN Dixin,et al.The effects of plant growth regulating substances on pollen germination and tube gowth in Fengshui pear(Pyrus serotina)[J].Acta Bot Boreal-Occident Sin,2003,23(4):586-591.
Storage temperature and pollen viability of Pinus yunnanensis
LI Mei,ZHOU Lanying
(College of Forestry,Sichuan Agricultural University,Chengdu 611130,Sichuan,China)
To determine the storage temperature and pollen viability of Pinus yunnanensis,different storage conditions of 25℃,5℃,and-20℃ were used to study variation in pollen viability.Different treatments of sucrose,H3BO3,GA3,and CaCl2were used to determine the effects on pollen germination.Analysis employed orthogonal experiments alone as well as interactions.Results showed that pollen viability was 91.8%at 0 d and then decreased with greater storage time at temperatures of 25℃>5℃>-20℃.Germination tests showed that the optimal concentration and the corresponding germination rate were as follows:sucrose-80 g·L-1and 82.4%,GA3-100 mg·L-1and 66.4%,H3BO3-300 mg·L-1and 18.2%,and CaCl2-100 mg·L-1and 57.0%.The best combination of four factors was 120 g·L-1sucrose+150 mg·L-1GA3+100 mg·L-1H3BO3+100 mg·L-1CaCl2with a pollen germination rate of 89.7%.Meanwhile,above a certain concentration of the culture medium had inhibitory effects on pollen germination.Thus,low temperatures increased pollen storage time.[Ch,3 fig. 2 tab.11 ref.]
forest tree breeding;Pinus yunnanensis;pollen;storage temperature;viability;germination
S722.3;Q944.42
A
2095-0756(2017)01-0063-05
2016-01-07;
2016-03-31
四川省科學(xué)技術(shù)攻關(guān)項(xiàng)目(2011NZ0098-10)
李梅,從事園林植物與觀賞園藝研究。E-mail:limei862@sina.com。通信作者:周蘭英,教授,博士,從事林木遺傳育種研究。E-mail:kelin1234@sina.com
浙 江 農(nóng) 林 大 學(xué) 學(xué) 報(bào),2017,34(1):68-77
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2017.01.011
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
少年博覽·小學(xué)高年級(jí)(2016年12期)2017-01-16 12:48:35
特別文摘(2016年19期)2016-10-24 18:38:15
37°女人(2016年5期)2016-05-06 19:44:06
爆笑show(2016年1期)2016-03-04 18:30:28
汽車觀察(2016年3期)2016-02-28 13:16:26
爆笑show(2015年6期)2015-08-13 01:45:40
