水分脅迫下檸條葉片氣孔導度對環境因子的響應
2017-02-05 14:52:07韓磊孫兆軍焦炳忠
江蘇農業科學 2016年10期
韓磊+++孫兆軍++焦炳忠

doi:10.15889/j.issn.1002-1302.2016.10.073
摘要:以寧夏河東沙地沙生灌木檸條為試驗材料,研究不同水分條件下檸條葉片氣孔導度對環境主要驅動因子的響應及敏感性。結果表明:隨著水分脅迫程度的增強,檸條葉片氣孔導度明顯降低,其過程與蒸騰速率的變化一致但不同步;在檸條葉片氣孔導度對光合有效輻射的響應方面,隨著土壤含水量的降低,氣孔啟動速度下降,但輕度水分脅迫下該響應過程的敏感性較高;充分供水、輕度水分脅迫、中度水分脅迫條件下,檸條葉片氣孔導度的主導因素為氣溫;對于重度水分脅迫(田間最大持水量的20%~40%),檸條葉片氣孔導度對溫度的敏感性降低,重度脅迫下水汽壓飽和虧缺成為影響檸條氣孔導度的主要因素,其貢獻率為76.6%(P<0.01)。
關鍵詞:檸條;氣孔導度;水分脅迫;環境因子;響應機制;河東沙地
中圖分類號: S184;S157文獻標志碼: A文章編號:1002-1302(2016)10-0265-04
收稿日期:2016-04-14
基金項目:寧夏自然科學基金(編號:NZ13025)。
作者簡介:韓磊(1985—),男,寧夏石嘴山人,博士,副研究員,主要從事水土保持、生態修復研究。E-mail:layhan@163.com。檸條(Caragana korshinskii Kom.)作為治理水土流失和退化沙化草場的先鋒植物,具有廣泛的適應性和很強的抗逆性。處于干旱、半干旱農牧交錯帶的寧夏河東沙區,長期以來大面積種植檸條人工林用于防風固沙和草地生態系統的恢復。但是由于該地區降水量稀少且無效蒸發量大,人工林的營造將改變沙地水分的再分配。近年來,隨著全球氣溫升高,該地區降水量以20~40 mm/10年的速度逐漸減少[1],氣候總體上呈現干化趨勢[2-3]。植物葉片對氣溫升高、降水減少的響應比較明顯,植物氣孔影響蒸騰、光合等生理機能,且隨著所處的環境狀況而時刻發生變化,在植物中起平衡調節作用[4],氣孔特性的具體變化則隨物種、干旱程度而呈現不同的特點。如干旱環境使大多數植物氣孔密度增大,氣孔相對開度變小[5-6];在干旱條件下,檸條受到明顯的水分脅迫,表現出明顯的光抑制現象[7]。諸多研究表明,植物葉片氣孔特性的異質性使其對脅迫環境響應也有很大差異[8],因此,研究不同水分條件下檸條葉片氣孔導度對環境主要驅動因子響應的敏感性,在全球變暖加速地球系統水循環的背景下,對于分析干旱脅迫下環境因子對河東沙地林木水分利用的影響機制尤為重要。
1材料與方法
1.1研究區概況
試驗地點位于寧夏河東沙地南緣的同心縣,地理坐標為36°52′06″N,105°59′7″E,海拔1 568 m,在氣候區上屬于溫帶大陸性氣候,年平均氣溫為8.7 ℃,極端最高氣溫為38.5 ℃,極端最低氣溫為-27.3 ℃,≥10 ℃年平均積溫為 2 963.1 ℃,無霜期為165~183 d;雨熱同季,多年平均降水量為251 mm,降水多集中在7—9月,占年降水量的72.4%;多年平均蒸發量為 2 340 mm;年日照時間多年平均值為 2 900~3 055 h。試驗區土壤類型為灰鈣土區的沙化土壤,田間最大持水量為21.3%,表層土壤平均容重為1.47 g/cm3。
1.2試驗方法
在試驗區選取3年生檸條植株,于2014年4月植入試驗花盆(高33 cm,上口徑35 cm,下口徑29 cm),試驗盆內土壤與試驗區大田土壤一致,栽植后將其置于試驗地的溫室內并充分補水,使其成活并正常生長。7月分別人工控制土壤水分含量(SWC)為田間最大持水量(w)的80%~100%(T1處理:充分供水)、60%~80%(T2處理:輕度水分脅迫)、40%~60%(T3處理:中度水分脅迫)、20%~40%(T4處理:重度水分脅迫),即各處理土壤水分含量:T1處理,17.0%~21.3%;T2處理,12.8%~17.0%;T3處理,8.5%~12.8%;T4處理,4.3%~8.5%。以上區間均含下不含上。每個處理3次重復,待長勢穩定后,于2014年8月開始觀測不同水分梯度下苗木葉片氣孔導度變化節律及其影響因子。
1.2.1植物葉片氣孔導度采用Li-6400(Li-Cor Inc.,Lincoln,USA)觀測,選擇苗木中上部向陽面生長良好的葉片,在生長季典型晴天,從7:00—19:00每隔2 h測定1次,每張葉片每次連續采集6個穩定的數據,取平均值。測定內容有氣孔導度(Gc)、蒸騰速率(Tr)、胞間CO2濃度(Ci)等。
1.2.2氣孔導度對光合有效輻射(PAR)的響應利用Li-6400-02B紅藍光源(Li-Cor Inc.,Lincoln,USA)設定模擬光輻射強度,模擬光輻射強度的梯度設置為0、50、100、200、400、600、800、1 000、1 200、1 500、1 800、2 000、2 200 μmol/(m2·s),觀測苗木葉片氣孔導度對光輻射強度的響應。
1.2.3環境因子采用美國Vantage pro 2 自動氣象站(Davis Inc.,USA),10 min記錄1次氣溫、相對濕度(2 m處)、光量子通量密度、降水量等氣象因子;同時采用ML3X土壤水分速測儀(Delta-T Inc.,UK)測定盆栽土壤單位體積含水量;土壤水分速測儀在測定前用烘干法標定。
1.3數據分析
采用SPSS 19. 0統計分析軟件對數據進行相關分析、單因素方差分析、多變量偏相關分析。
2結果與分析
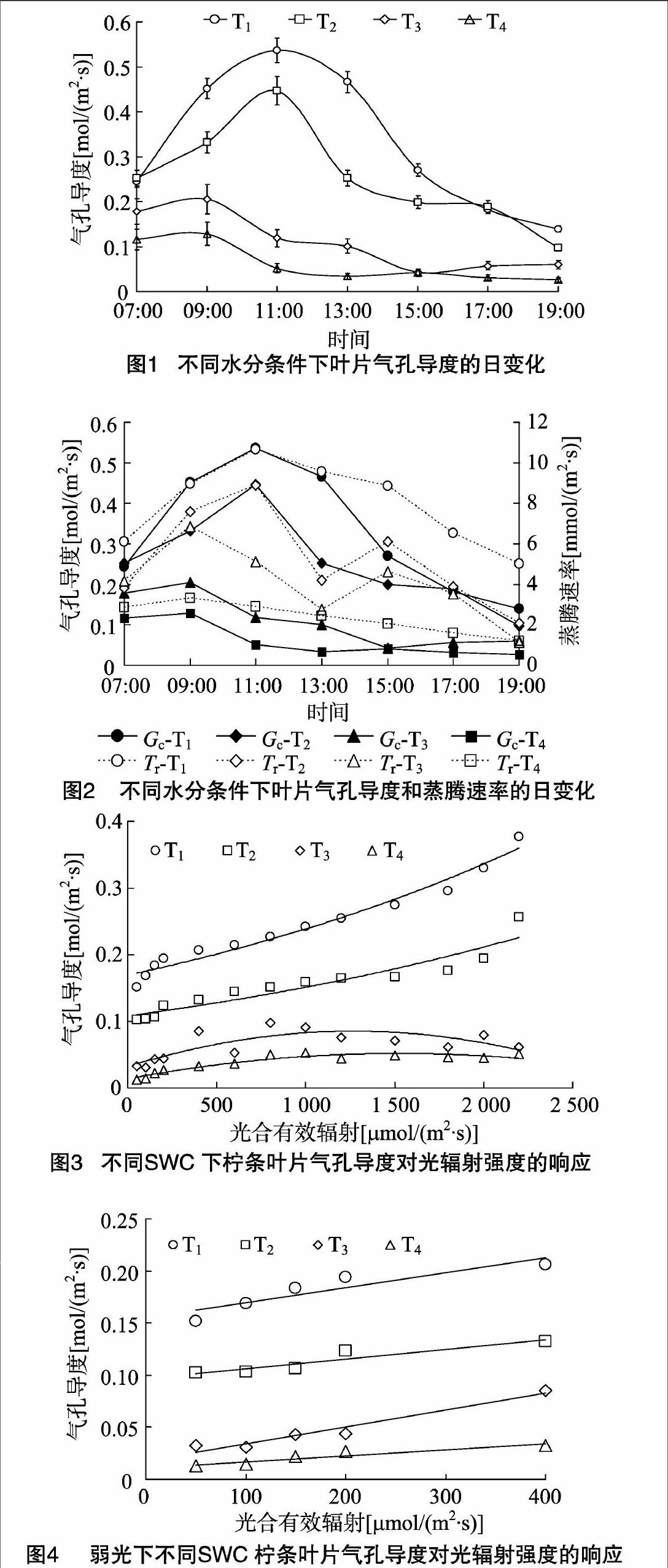
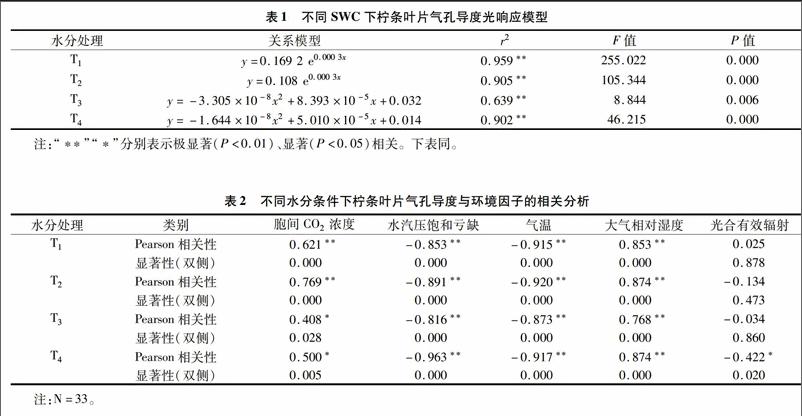
2.1不同土壤水分下檸條葉片氣孔導度的日變化節律
氣孔是植物葉片與外界環境進行氣體、水分交換的重要通道,在調節植物水分狀況中起關鍵作用。當供水和蒸騰需水之間不平衡,特別是供小于需時,植物會適時作出各種響應以減輕脅迫[9]。由圖1可見,檸條葉片在水分脅迫下(T3、T4處理),氣孔導度峰值前移,于09:00達到峰值,之后葉部水分虧缺提早出現,正午前后葉片氣孔逐漸閉合,氣孔導度逐漸降低;而T1、T2處理下氣孔導度在11:00達到峰值,T1、T2處理日均值分別為(0.327±0.156)、(0.253±0.112)mol/(m2·s);在低水平的土壤含水量下,苗木葉片氣孔導度的日變化相差不大,都維持在較低的水平,T3、T4處理的日均值分別為(0.109±0.063)、(0.061±0.043) mol/(m2·s)。受水分脅迫,T3、T4處理植物葉片氣孔導度峰值與T1、T2處理相比分別下降了61.7%、54.1%,76.1%、71.4%。氣孔的運動直接調控著蒸騰速率的變化,檸條氣孔導度與蒸騰速率的變化規律較為一致但并不同步(圖2)。中午由于光照度大、氣溫高、空氣濕度低,植物為減少過度蒸騰失水,葉片氣孔開度會減小或關閉;15:00,在T2、T3處理下,蒸騰的減弱使葉細胞的含水量得到一定程度的恢復,Tr分別出現峰值,而Gc值則處于較低點,這是由于此時大氣溫度較高甚至達到最高值,保衛細胞因氣溫升高失水使氣孔逐漸關閉,而植物的蒸騰作用因組織儲存水而滯后于氣孔變化[10-11]。隨著光照度的減弱,日落前后氣孔逐步關閉,蒸騰作用逐漸減弱。
2.2不同土壤水分條件下檸條葉片氣孔導度對光合有效輻射的響應
在假設水汽、溫度、濕度恒定的條件下,分析Gc對PAR的響應。由圖3、圖4可見,T1、T2水分處理下,Gc、PAR呈現指數正相關關系(P<0.001),擬合方程為Gc=aePAR,可以解釋95.9%、90.5%的變量,同時隨著土壤含水量的增加,Gc對PAR的敏感性(a值)增加;當水分脅迫(T3、T4處理)增大時,檸條葉片氣孔導度隨著光輻射強度的增加先增大后減小,即在PAR<400 μmol/(m2·s)時,Gc隨著PAR的增加迅速增大,之后檸條葉片氣孔導度對光合有效輻射的響應強度變弱;當光合有效輻射達到1 000 μmol/(m2·s)左右時,PAR進一步增強,植物葉片氣孔導度逐漸下降,以防組織水勢過度降低造成植物萎蔫,用擬合方程Gc=aPAR2+bPAR+c來描述,可以解釋63.9%、90.2%的變量(P<0.01),詳見表1。在較低光照度[PAR<400 μmol/(m2·s)]下,無論是充分供水還是水分脅迫條件下,檸條葉片氣孔導度隨著PAR的增加而增大,擬合方程為Gc=kPAR+c,但啟動速度(可用常數c代表)不同,由小到大依次為T1處理(0.155)>T2處理(0.097)>T3處理(0.018)>T4處理(0.011);在低光照度下,Gc對PAR的敏感性(k)由高到底依次為T3處理(k=0.000 2,r2=0932**,P=0.005)>T1處理(k=0.000 1,R2=0.776*,P=0.031)>T2處理(k=9.322×10-5,r2=0.829*,P=0.02)>T4處理(k=5.838×10-5,r2=0.871*,P=0.013)。由此可見,在低光照度下,隨著土壤含水量的降低,檸條葉片氣孔導度對光合有效輻射的響應表現為氣孔啟動速度下降,但輕度水分脅迫下該響應過程的敏感性較高。
2.3不同土壤水分條件下檸條葉片氣孔導度與主要環境因子的相關性分析
由表2可以看出,從環境因子對檸條葉片氣孔導度的影響來看,檸條葉片氣孔導度與氣溫(Ta)、水汽壓飽和虧缺(VPD)、相對濕度(RH)、輻射強度、胞間CO2濃度有顯著相關關系,但在不同土壤水分條件下,影響氣孔導度的主要因子略有不同。在T1、T2處理下,Ta、VPD、RH、Ci是影響檸條葉片氣孔導度的主要因子,且呈現極顯著的相關關系。由表3偏相關分析結果可知,在T1、T2、T3處理下,檸條葉片氣孔導度的主導因素為大氣溫度(Ta),貢獻率r2分別為49.7%、292%、72.4%。在充分供水(T1處理)下,檸條氣孔導度的主要影響因子由大到小排序:Ta(偏相關系數r=-0.705,P<0.01)>VPD(偏相關系數r=0.399,P<0.05);在輕度水分脅迫(T2處理)下,除Ta外(偏相關系數r=-0.540,P<001),其他氣象因子對氣孔導度的影響不顯著;在中度水分脅迫(T3處理)下,檸條氣孔導度的主要影響因子由大到小排序:Ta(偏相關系數r=-0.851,P<0.01)>RH(偏相關系數r=-0.831,P<0.01)>VPD(偏相關系數r=0.773,P<001)>Ci(偏相關系數r=0.415,P<0.05),其貢獻率r2分別為 72.4%、69.1%、56.7%、17.2%;在重度水分脅迫(T4處理)下,檸條葉片氣孔導度對溫度的敏感性降低,且隨著溫度的升高呈現顯著的正相關,其影響因子由大到小排序:VPD(偏相關系數r=-0.875,P<0.01)>Ta(偏相關系數r= 0639,P<0.01)>Ci(偏相關系數r=0.530,P<0.01)。可以看出,重度脅迫下VPD成為影響檸條氣孔導度的主要因素,其貢獻率為76.6%。
3討論與結論
植物氣孔是調控“土壤-植被-大氣”連續體之間物質和能量交換的關鍵環節[12],氣孔對環境因子的響應過程及其對主要驅動因子響應的敏感性隨物種、干旱程度而不同。水分對葉片氣孔導度的影響表現為水分的降低會對氣孔導度產生抑制作用。隨著水分脅迫程度的增強,檸條葉片氣孔導度明顯降低,蒸騰失水減弱,檸條氣孔導度與蒸騰速率的變化規律較為一致但并不同步,表現在午后(15:00)植物的蒸騰作用因組織儲存水而滯后于氣孔變化。正午前后氣孔關閉后的葉組織細胞水分得到一定程度的恢復,Tr出現2次峰值,而此時Gc值仍處于較低點;之后隨著光照度的減弱,蒸騰作用隨著氣孔導度的降低逐漸下降。該過程說明,檸條通過控制氣孔的開放程度來適應較干旱的環境,以至于在較干旱、高溫的環境下表現出較強的水分調節能力。這與柯世省等研究水分脅迫對云錦杜鵑、紫花苜蓿蒸騰速率和氣孔導度的影響結果[13-14]較為一致。
光合有效輻射強度對檸條葉片氣孔導度的影響與土壤有效水供給能力有關。在充分供水、輕度水分脅迫下,氣孔導度對光合有效輻射的敏感性隨著土壤含水量的增加而增加;當中度、重度水分脅迫出現時,檸條葉片氣孔導度隨著光輻射強度的增加先增大后減小,即在PAR<400 μmol/(m2·s)時,Gc隨著PAR的增加迅速增大,之后檸條葉片氣孔導度對光合有效輻射的響應強度變弱,當光合有效輻射達到 1 000 μmol/(m2·s) 左右時,PAR進一步增強,植物葉片氣孔導度逐漸下降,以防止組織水勢過度降低造成植物萎蔫,這與干旱脅迫下檸條葉片蒸騰的光響應過程較為一致[15]。在低光照度下,檸條葉片氣孔導度對光合有效輻射的響應表現為隨著土壤含水量的降低,氣孔啟動速度下降,但輕度水分脅迫下該響應過程的敏感性較高。
由于不同植物的遺傳特性、測定時的環境、時間尺度的不同,以及未考慮各個環境因子的相互作用對氣孔導度的影響,由此得到的氣孔導度與環境因子之間的關系也不盡一致[16]。不同土壤水分條件下主要環境因子對檸條葉片氣孔導度的綜合影響表明,在充分供水、輕度水分脅迫、中度水分脅迫條件下,檸條葉片氣孔導度的主導因素為大氣溫度;對于重度水分脅迫(田間最大持水量的20%~40%),檸條葉片氣孔導度對溫度的敏感性降低,重度脅迫下水汽壓飽和虧缺(VPD)成為影響檸條氣孔導度的主要因素,說明在缺水情況下檸條氣孔對VPD的響應比較高土壤水分含量處理敏感。氣孔對環境因子的響應規律是十分復雜的,各種環境因子間以及環境因子對植物體的作用都存在交互作用[17-20],葉片氣孔特性具有異質性,且其對脅迫環境響應也有很大的差異[8,21-22],如左應梅等分析土壤相對含水量、光合有效輻射和空氣相對濕度對華南8號木薯的綜合影響發現:當土壤相對含水量較低時,土壤相對含水量是影響氣孔導度的主導因子[23];隨著土壤相對含水量的增加,影響氣孔導度的主導因子由土壤相對含水量轉變為光合有效輻射。孫林等對華北落葉松冠層氣孔導度的影響因子研究認為,PAR是Gc的驅動因子,VPD則是Gc的限制因子[24]。氣孔對干旱的適應方式有前饋式反應、反饋式反應2種[4],對于檸條在重度水分脅迫下的表現則為前饋式反應,即水氣壓虧缺直接作用而引起氣孔關閉以阻止水勢的進一步下降,從而建立預警系統。
參考文獻:
[1]方鋒,孫蘭東,郭俊琴,等. 中國西北地區城市經濟發展對降水趨勢的影響[J]. 自然資源學報,2014,29(11):1878-1887.
[2]徐小玲,延軍平. 近30年毛烏素沙區的氣候與厄爾尼諾/拉尼娜事件的相關分析[J]. 干旱區研究,2003,20(2):117-122.
[3]王亞軍,高尚玉,馬玉貞,等. 基于樹輪重建的寧夏河東沙地西部公元1899年來年降水量的變化[J]. 干旱區地理,2010,33(3):377-382.
[4]司建華,常宗強,蘇永紅,等. 胡楊葉片氣孔導度特征及其對環境因子的響應[J]. 西北植物學報,2008,28(1):125-130.
[5]高彥萍,馮瑩,馬志軍,等. 水分脅迫下不同抗旱類型大豆葉片氣孔特性變化研究[J]. 干旱地區農業研究,2007,25(2):77-79.
[6]Martinez J P,Silva H,Ledent J F,et al. Effect of drought stress on the osmotic adjustment,cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.)[J]. European Journal of Agronomy,2007,26(1):30-38.
[7]肖春旺. 施水量變化對毛烏素沙地4種優勢植物葉綠素熒光的影響[J]. 草業學報,2002,11(2):85-90.
[8]王碧霞,曾永海,王大勇,等. 葉片氣孔分布及生理特征對環境脅迫的響應[J]. 干旱地區農業研究,2010,28(2):122-126.
[9]孫谷疇,趙平,曾小平,等. 亞熱帶森林演替樹種葉片氣孔導度對環境水分的水力響應[J]. 生態學報,2009,29(2):698-705.
[10] Han L,He K N,Hu X B,et al. Canopy transpiration response to environmental variations in Platycladus orientalis:Properties and modeling[J]. Pakistan Journal of Botany,2012,44(2):541-545.
[11]趙秀華,趙平,朱麗薇,等. 不同樹高對荷木液流密度、整樹蒸騰量和時滯的效應[J]. 廣西植物,2015,35(1):53-60.
[12]唐鳳德,武耀祥,韓士杰,等. 長白山闊葉紅松林葉片氣孔導度與環境因子的關系[J]. 生態學報,2008,28(11):5649-5655.
[13]柯世省,魏燕,陳賢田,等. 云錦杜鵑氣孔導度和蒸騰速率對水分的響應[J]. 安徽農業科學,2007,35(21):6363-6365.
[14]羅永忠,成自勇. 水分脅迫對紫花苜蓿葉水勢、蒸騰速率和氣孔導度的影響[J]. 草地學報,2011,19(2):215-221.
[15]韓磊,孫兆軍,展秀麗,等. 寧夏河東沙區檸條植株葉片蒸騰對干旱脅迫的響應[J]. 生態環境學報,2015,24(5):756-761.
[16]高冠龍,張小由,常宗強,等. 植物氣孔導度的環境響應模擬及其尺度擴展[J]. 生態學報,2016,36(6):1491-1500.
[17]吳大千,徐飛,郭衛華,等. 中國北方城市常見綠化植物夏季氣孔導度影響因素及模型比較[J]. 生態學報,2007,27(10):4141-4148.
[18]劉廣志,陳炳佑,侍福梅. MAP18參與了脫落酸調控的擬南芥氣孔關閉及根生長[J]. 江蘇農業科學,2015,43(11):55-57.
[19]楊天梅,許宗亮,左應梅,等. 橫斷山區珠子參不同居群葉片形態變異及其與環境因子的關系[J]. 江蘇農業科學,2015,43(9):276-280.
[20]朱美瑛,王康才,李雨晴. 金線蓮葉片氣孔與光合特性研究[J]. 江蘇農業科學,2014,42(11):210-212.
[21]馮海萍,曲繼松,張麗娟,等. 典型旱作區不同田間持水量對塑料拱棚春茬番茄光合生理特性的影響[J]. 江蘇農業學報,2014,30(2):381-387.
[22]徐剛,彭天沁,高文瑞,等. 氮肥水平對木薯渣復合基質栽培黃瓜生長及光合作用的影響[J]. 江蘇農業學報,2015,31(1):68-72.
[23]左應梅,陳秋波,鄧權權,等. 土壤水分、光照和空氣濕度對木薯氣孔導度的影響[J]. 生態學雜志,2011,30(4):689-693.
[24]孫林,管偉,王彥輝,等. 華北落葉松冠層平均氣孔導度模擬及其對環境因子的響應[J]. 生態學雜志,2011,30(10):2122-2128.
