藏族雙語者與漢族非雙語者腦灰質VBM-MRI的初步研究
2017-02-02 05:43:59馮祥鮑海華王芳芳何盈秦粽園
磁共振成像 2017年10期
馮祥,鮑海華,王芳芳,何盈,秦粽園
青海省少數民族人數所占比重僅次于西藏、新疆,居全國第3位,而青海少數民族中藏族人數所占比重僅次于漢族,居全省第2位。因此,青海省是少數民族尤其是藏族人民聚居的地方,許多藏民從小開始就學習藏語和漢語兩種語言。漢語和藏語雖同屬漢藏語系,但屬不同語族。因此,漢語和藏語相比有著許多不同之處:(1)文字結構不同,漢字是方塊文字,屬象形文字,而藏文是拼音文字,屬于線性文字;(2)意思表達方式不同,漢字主要通過字形表達字意,而藏文則主要是通過讀音直接表達字意。人類有學習兩種或多種語言的能力,通常將那些可以正式使用兩種或多種語言的人稱為雙語者或多語者。目前國內外很多學者主要致力于對雙語或多語者腦功能的研究,關于雙語或多語的習得對大腦結構影響的研究則相對較少,而目前關于藏族藏-漢雙語的習得對腦結構影響的研究則更為少見。本試驗通過對藏族雙語者與漢族非雙語者全腦灰質結構的研究,初步探討少數民族第二語言習得對大腦結構的影響。
基于體素形態學測量(voxel-based morphometry,VBM)技術是一種能夠對腦結構形態進行自動分析的技術,可定量檢測腦組織各組分的密度和體積,從而可以全面評估腦結構的形態學改變。相比感興趣區(region of interest,ROI)分析方法,此技術可以消除人為主觀因素的影響,具有客觀性好、操作簡便、可重復操作等優點,因而在臨床得到廣泛應用。
1 材料與方法
1.1 一般資料
招募70名青海本地健康志愿者參與此次研究,分為兩組。雙語組:35名掌握藏-漢雙語的藏族健康志愿者,男20例,女15例,年齡在18~25歲,平均年齡為(21.09±1.26)歲,受教育年限時間平均(15.26±1.42)年,所有藏族志愿者世代出生并長期居住于青海地區。對照組:35名漢族非雙語健康志愿者,男18例,女17例,平均年齡為(21.23±1.61)歲,受教育年限時間平均(15.71±1.76)年,所有漢族志愿者來自兩代以上出生并長期居住于青海地區。納入標準:(1)無MRI檢查禁忌證;(2)常規頭顱MRI掃描腦實質無異常;(3)無神經病和精神病癥狀及體征;(4)右利手。排除標準:(1)有慢性高原病;(2)有腦部外傷史;(3)被確診的腦神經失調。所有受檢者在檢查前均知道此次檢查的內容及意義,并簽署了知情同意書,本研究經我院倫理委員會批準。
1.2 MRI掃描方法
應用Philips Achieva 3.0 T TX多源發射MR成像系統,標準頭顱8通道相控陣線圈,向所有受檢者詳細說明檢查的全部過程以取得其更好的配合,囑受檢者平臥于檢床上,保持靜息狀態。先行T1WI和T2WI序列掃描,確定腦實質無異常者再行VBM掃描。應用超快速場回波序列獲得3D-T1結構像,掃描參數:TR=7.5 ms,TE=3.7 ms,矩陣256×256,層厚2 mm,激發角度7°,全腦掃描共獲取176幅圖像。所有志愿者由同一名的優秀影像科技師完成掃描。
1.3 數據處理及統計分析
首先應用Dcm2AsiszImg軟件對原始的DICOM數據進行分類整理,然后采用統計參數圖(statistical parametric mapping,SPM8)的嵌套軟件VBM8 toolbox進行數據處理,最后在MATLAB平臺上進行計算和圖像處理。數據處理的具體步驟包括:(1)原始數據分類整理;(2)頭動較正;(3)分割3D-T1結構像,獲得灰質、白質及腦脊液圖像,在圖像模板上進行圖像配準,然后空間標準化;(4)采用半高全寬為8 mm的三維高斯核進行圖像空間平滑。最后采用兩樣本t檢驗進行比較(P<0.05;Alphasim校正),選取相鄰體素大于389的組塊視為有差異的腦區,將有差異的腦區疊加到T1結構像的模板上,分析研究對象腦灰質體積的變化。
2 結果
本次研究中藏族雙語者較漢族非雙語者腦灰質體積增加的腦區包括:左側中央后回、緣上回、顳上回和右側顳上回、豆狀核、頂下小葉以及左側小腦8區(見表1,圖1)。另外,左側豆狀核、右側三角部額下回、雙側前扣帶回和扣帶旁回、右側小腦4、5、6區及左側小腦9區等因體素增加較小,未列入表內。
3 討論
近年來國內外學者從不同層面、不同水平對語言腦功能區進行了深入研究,但主要集中在對不同語言腦激活區方面的研究,而關于雙語或多語的習得導致腦微觀結構改變的研究報道則較少,關于藏族雙語習得導致腦微觀結構改變的研究則更少。

表1 藏族雙語組與漢族非雙語組相比腦灰質體積增加的統計分析結果Tab.1 The statistical analysis results of increased brain gery matter volume in Tibetain Bilingual group compared with Chinese monoligual group
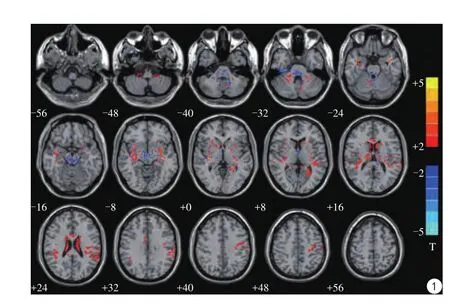
圖1 藏族雙語組與漢族非雙語組相比灰質體積有差異的腦區統計參數圖。紅色代表藏族雙語組較漢族非雙語組灰質體積增加的腦區,包括左側中央后回、左側緣上回、左側顳上回、右側顳上回、右側豆狀核、右側頂下小葉、左側小腦8區(統計閾值設為P<0.05,cluster size>389,Alphasim校正)Fig.1 The statistical parametric map of the brain areas with different grey matter volumes compared Tibetan-Chinese bilingual group with Chinese monoligual group. Red represents the Tibetan-Chinese bilingual group than Chinese monoligual group increased gray matter volume of the brain,including: left postcentral gyrus, left supramarginal gyrus, left superior temporal gyrus, right superior temporal gyrus, right putamen, right inferior parietal lobule, left cerebellum area 8. The statistical threshold is set to P<0.05,cluster size>389, Alphasim correction.
3.1 結構差異區的討論
大腦皮層為大腦的表層部分,是生物進化的最高成果和人類思維的神經基石,也是我們意識活動的物質基礎。大量研究表明,人腦解剖結構具有可塑性[1],可以隨著特殊生活經歷而發生改變[2],以適應周圍客觀事實的改變。國外學者Mechelli[3]、Christian[4]、Eleanor[5]等分別通過對意-英雙語者和英語非雙語者、專業和業余音樂家、出租車和非出租車司機的研究,發現后天的鍛煉和學習可以導致大腦微觀結構的改變,神經元的可塑性是導致人類發展過程中大腦灰質隨著環境發生灰質結構變化的重要原因[6]。
顳上回為聽覺性語言中樞。Siok等[7]研究發現,使用拼音文字的人,常運用的腦語言功能區為威爾尼克區(wernick-area)。Wernick區包括左側顳上回、顳中回后部及左側頂下小葉的角回和緣上回。楊振燕等[8]對中英文語言活動區的研究中發現完成中文任務時激活左右側顳上回,而完成英文任務時只激活左側顳上回。王云玲等[9]研究多語者語言功能區時發現,在完成漢語任務時明顯激活右側顳上回,而右側顳上回在完成英語任務時未見激活。Tan等[10]利用功能性磁共振成像(functional magnetic resonance imaging,fMRI)研究發現在完成漢字閱讀任務時能激活右顳上回前部等腦區,這些腦區以往在拼音文字的語言活動中很少激活。Pierce等[11]研究發現漢-法雙語者在完成聽到漢語辨別詞匯聲調任務時,雙側顳上回激活。本研究結果雙側顳上回灰質體積均增加與上述研究相符,藏文屬于拼音文字,長期的文字學習和語言交流導致左側顳上回腦結構的改變;漢語是一種象形表意文字,具有復雜的空間二維結構,通過字形提取字義是需要更多右側大腦半球的支持,而右側顳上回僅在完成漢語任務時明顯激活,筆者推測右側顳上回與第二語言漢語的習得有關,并可能為處理漢語的特異性腦區。
頂下小葉被稱為“Geschwind區”[12],Geschwind提出頂下小葉可能與語言的表征有關。頂下小葉可分為角回和緣上回兩個亞區。目前很多相關研究認為涉及雙語轉換腦區包括額下回、前扣帶回、頂下小葉(尤其是角回)和左側文狀體等腦區[13-16]。Elise等[17]研究用第二語言法語進行閱讀任務時發現左側頂下小葉激活,而第一語言英語進行閱讀任務時左側頂下小葉未見激活。Jasińska等[18]研究發現在完成任務時雙語者較單語者雙側頂下小葉明顯激活。本研究中右側頂下小葉、左側緣上回體積明顯增加與上述研究基本相符,藏-漢雙語者漢語的熟練程度低于藏語,因此在漢語的使用過程中常涉及漢語與藏語的轉換以便語義的理解,長期第二語言與母語的轉換導致腦語言轉換區頂下小葉腦結構發生改變。
豆狀核由核殼和蒼白球組成,是構成基底神經節核團必不可少的組分。近年來我國學者通過正電子發射型計算機斷層顯像(positron emission computed tomography,PET)發現雙側豆狀核參與了漢字的書寫。朱麗娜等[19]通過fMRI對維吾爾族漢語功能區的研究發現雙側豆狀核激活。刑一蘭等[20]通過PET-CT研究發現右側核殼參與了中文的書寫。目前通過進一步研究發現,基底節可能參與了對語音和語義的處理、詞法和語法的記憶,對語言進行加工、整理和協調,具有皮層下整合中樞的作用[19]。本研究中右側豆狀核體積增加與上述研究基本相符,藏-漢雙語者在學習漢語的同時,不僅要學習漢族的語言,還要學習漢族的文字,后天的語言說話和文字書寫共同作用使豆狀核的微觀結構發生了改變。通過此研究,也進一步證明皮層下神經核團參與了對語言的加工、處理。
小腦是運動的重要調節中樞,主要參與對運動的控制。近幾年來許多學者對小腦在語言加工中的作用進行了深入的探討[21],發現小腦在語言功能方面對其他腦功能區主要起支持協助作用。Pillai等[22]通過fMRI對西班牙語-英語雙語者的研究發現,無論給予何種語言任務,左側小腦半球都被激活。揭冰等[23]研究發現在完成對語義的辨別任務中右側小腦半球明顯被激活,并認為小腦對其他腦語言功能區起支持作用。王云玲等[9]通過fMRI研究維吾爾族多語者發現無論給予英語、漢語還是維語語言刺激時,都會有小腦的不同腦區被激活。本研究中雙側小腦半球多個腦區體積增加與上述研究基本相符,語言的各腦功能區并不是單獨存在的,而是相互聯系共同構成一條語言加工的神經通路,小腦對藏語、漢語的加工可能起著支持輔助的作用。
中央后回為軀體深感覺和淺感覺的皮質中樞。本研究中藏族雙語者左側中央后回灰質體積較漢族非雙語者增加,這與晁芳等[6]的研究結果白-漢雙語者較漢族非雙語者左側中央后回密度增高一致。盡管目前國內外關于中央后回與語言有關的相關文獻及報道并不多,但筆者認為左側中央后回可能與雙語者第二語言的習得相關,其可能是第二語言神經通路的一部分,筆者會在今后的研究中進一步加以證實。
本研究所得的結果與國內外研究的結果并不完全一致,考慮原因可能由以下兩方面所致:(1)語系不同。漢語和藏語雖屬漢藏語系,但維吾爾語屬于阿爾泰語系,英語屬于印歐語系,不同語系在學習過程中可能激活的神經元不同。(2)受試者個體差異。受試者第二語言的熟練程度、學習年齡等也與腦語言區結構的改變相關。
3.2 結論
本研究應用VBM技術對藏族雙語者與漢族非雙語者腦灰質進行了初步的研究,證明第二語言的習得可以導致部分腦語言區體積的增加,雙語學習可以促進腦語言區皮質的發育。右側顳上回可能為處理漢語的特異性腦區,左側中央后回可能是第二語言神經通路的一部分,筆者會在今后的研究中進一步加以證實。
3.3 不足之處
通常把可以使用兩種正式語言的人稱為雙語者,并且兩種語言都要達到同等熟練的程度,如果按照這樣的標準,本研究中的藏族雙語志愿者并不是最完美的雙語志愿者。此外,目前VBM技術雖然已經比較成熟,但仍存在一些局限性,例如組織分割、空間標準化及平滑處理存在一定的缺陷,但相信隨著后處理技術的不斷改進,研究結果的準確性也將不斷提高。
3.4 臨床意義
今后,筆者也會逐步完善對雙語者或多語者的選擇條件,包括語言的學習時間、學習方式、各種語言的熟練程度等,以更加準確地探討雙語或多語的習得對腦語言區結構的影響,進一步增加對腦語言結構區的認識,為臨床手術過程中減少或避免對腦語言區的損害以及術后腦語言功能區的恢復評估提供更多的影像學依據。
[References]
[1] Draganski B, Gaser C, Busch V, et al. Neuroplasicity: changes in grey matter induced by training. Natural, 2004, 427(6972): 311-312.
[2] Zou L, Ding G, Abutalebi J, et al. Structural plasticity of the left caudate in bimodal bilinguals. Cortex, 2012, 48(9): 1197-206.
[3] Mechelli A, Crinion JT, Noppeney U, et al. Neurolinguistics:structural plasticity in the bilingual brain. Nature, 2004, 431(7010): 757.
[4] Gaser C, Schlaug G. Brain structures differ between musician and non-musicians. Neurosci, 2003, 23(27): 9240-9245.
[5] Maguire EA, Gadian DG, Johnsrude IS, et al. Navigation-related sturctural change in the hippocampi of taxi dricers. Proc Natl Acad Sci USA, 2000, 97(8): 4398-4403.
[6] Chao F. Study of differences in brain structure and function between Bai-Chinese bilinguals and Chinese monoliguals. Kunming:Kunming Med Univ, 2013.晁芳. 白—漢雙語者與漢族非雙語者的全腦結構與功能差異研究.昆明: 昆明醫科大學, 2013.
[7] Siok WT, Perfetti CA, Tan LH, et al. Biological abnormality of impaired reading is constrained by culture. Nature, 2004, 431(7004):71-76.
[8] Yang ZY, Zhao XH, Dai GH. Functional magnetic resonance imaging study Chinese and English language active areas. Chin J Tissue Engin Rese, 2003, 7(10): 1492-1494.楊振燕, 趙小虎, 戴工華. 中英文語言活動區功能磁共振成像研究.中國臨床康復, 2003, 7(10): 1492-1494.
[9] Wang YL. fMRI study on the language areas of the Xinjiang Uygur multi-speakers. Urumqi: Xinjiang Med Univ, 2012.王云玲. 新疆維吾爾族多語者語言功能區的fMRI的研究. 烏魯木齊: 新疆醫科大學, 2012.
[10] Tan LH, Feng CM, Gao JH, et al. An fMRI study with written Chinese. Neuroreport, 2000, 12(1): 83-88.
[11] Pierce LJ, Klein D, Chen JK, et al. Mapping the unconscious maintenance of a lost first language. Proc Natl Acad Sci U S A, 2014,111 (48): 17314-17319.
[12] Catani M, Jones DK, Ffutche DH. Perisylvian langguage networks of the human brian. Ann Neuro, 2005, 57(1): 8-16.
[13] Chandrasekaran B, Chan AH, Wong PC. Neural processing of what and who information during spoken languange processing. Cogn Neuropsychol, 2011, 23(10): 2690-2700.
[14] Tan IH, Chen I, Yip V, et al. Activity levels in the left hemisphrer caudate-fusiform circuit predict how well a second language will be learned. Proc Natl Acad Sci U S A, 2011, 10(6): 2540-2544.
[15] Szaflarski JP, Vannest J, Wu SW, et al. Excitatory repetitive transcranial magnetic stimulation induces improvements in chronic post-stroke aphasia. Med Sci Monit, 2011, 17(3): 132-139.
[16] Kozub J, Urbanik A, Chrzan R, et al. Presurgical functional brain examination MR (fMRI). Przegl lek, 2010, 67(4): 326-329.
[17] Barbeau EB, Chai XJ, Chen JK, et al. The role of the left inferior parietal lobule in second language learning: An intensive language training fMRI study. Neuropsychologia, 2016, 10(3): 169-176.
[18] Jasińska KK, Petitto LA. Development of neural systems for reading in the monolingual and bilingual brain: new insights from functional near infrared spectroscopy neuroimaging. Dev Neuropsychol, 2014,39(6): 421-439.
[19] Zhu LN. Magnetic resonance imaging study of functional areas of Chinese of vigeo. Urumqi: Xinjiang Med Univ, 2010.朱麗娜. 維吾爾族漢語功能區的磁共振研究. 烏魯木齊: 新疆醫科大學, 2010.
[20] Xing YL. Mandarin and English writing recruit basal ganglia:Evidence from PET-CT. Guangzhou: South Med Univ, 2010.邢一蘭. 基底神經節參與中英文書寫—-來自PET-CT的證據. 廣州: 南方醫科大學, 2010.
[21] Gil RS, Gatignol P, Capelle L, et al. The role of dominant striatum in language:a study using intraoperative electrical stimulations. J Neurol Neurosurg Psychiatry, 2005, 76(7): 940-946.
[22] Pillai JJ, Araque JM, Allison JD, et al. Functional MRI study of semantic and phonological language processing in bilingual subject:preliminary findings. Neuroimage, 2003, 19(3): 565-576.
[23] Jie B, Zhao XH, Yang ZY, et al. fMRI preliminary evaluation of cerebellar language function. J Tongji Univ, 2005, 26(6): 22-24.揭冰, 趙小虎, 楊振燕, 等. 小腦語言功能的fMRI初步評價. 同濟大學學報, 2005, 26(6): 22-24.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
哲學評論(2021年2期)2021-08-22 01:53:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
文苑(2020年4期)2020-05-30 12:35:30
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生作文(中高年級適用)(2018年3期)2018-04-18 01:24:47
華北電力大學學報(社會科學版)(2016年4期)2016-12-01 03:59:30
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
