沙米和藜麥種子代謝組比較分析
2017-01-17 02:29:31趙杰才馬小飛陳國雄
中國食物與營養(yǎng) 2016年12期
趙杰才,麻 彥,周 琴,趙 昕,馬小飛,陳國雄
(中國科學(xué)院寒區(qū)旱區(qū)環(huán)境與工程研究所/沙坡頭沙漠試驗(yàn)研究站/甘肅省寒區(qū)旱區(qū)逆境生理與生態(tài)重點(diǎn)實(shí)驗(yàn)室,蘭州 730000)
沙米和藜麥種子代謝組比較分析
趙杰才,麻 彥,周 琴,趙 昕,馬小飛,陳國雄
(中國科學(xué)院寒區(qū)旱區(qū)環(huán)境與工程研究所/沙坡頭沙漠試驗(yàn)研究站/甘肅省寒區(qū)旱區(qū)逆境生理與生態(tài)重點(diǎn)實(shí)驗(yàn)室,蘭州 730000)
為解析沙米和藜麥兩種植物的種子代謝物,利用GC×GC-TOF MS平臺開展非靶向沙米和藜麥種子代謝組分析。科爾沁沙地沙米、騰格里沙漠沙米和山西靜樂藜麥種子檢測到了106個代謝物。沙米代謝物相對含量有6個顯著高于、17個顯著低于藜麥,其余代謝物沒有顯著差異。在沙米代謝組中相對含量較高,而在稻米和小麥代謝組中都沒有檢測到的代謝物有棉籽糖、原兒茶酸和木糖醇等,這些代謝物可彌補(bǔ)稻米和小麥的營養(yǎng)不足。對以稻米或小麥為主食的人群來說,沙米是理想的藥食同源食物。
沙米;藜麥;代謝物;藥食同源;流動沙丘
沙蓬米即沙米,是莧科藜亞科一年生野生草本植物,生長于干旱半干旱地區(qū)流動沙丘或裸露沙地上,是沙區(qū)的傳統(tǒng)美食。沙米種子蛋白質(zhì)、脂肪酸和碳水化合物含量分別為23%、10%、45%左右,其營養(yǎng)價(jià)值不亞于藜麥[1]。藜麥也是莧科藜亞科一年生草本植物,原產(chǎn)于南美洲安第斯山脈,已有5 000年的馴化歷史。由于藜麥的高營養(yǎng)價(jià)值及其耐旱性,聯(lián)合國糧農(nóng)組織把2013年定為國際藜麥年[2],旨在向全世界推薦藜麥。
代謝組是生物體內(nèi)小分子(MW<1 000)代謝物總和,是基因表達(dá)和代謝產(chǎn)生的最終產(chǎn)物或中間產(chǎn)物,處于生命信息傳遞的終端,它能表現(xiàn)出生物體系整體功能或狀態(tài)的最終結(jié)果[3]。代謝組分析可應(yīng)用于微生物學(xué)、植物學(xué)、食品及營養(yǎng)科學(xué)、毒理學(xué)研究、臨床疾病診斷及藥物開發(fā)等領(lǐng)域[3-6],在農(nóng)作物種子分析方面的應(yīng)用有黃豆[7-9]、小麥[10,11]、水稻[12,13]、大麥[14]等。本文用非靶向方法開展了沙米和藜麥種子的代謝組分析,以揭示2種植物種子代謝物的異同。
1 材料與方法
1.1 試驗(yàn)材料
選用2個野生沙米群體種子,一種為地處科爾沁沙地的遼寧省阜新市彰武縣阿爾鄉(xiāng)群體種子(此后稱阿爾鄉(xiāng)沙米);另一種是地處騰格里沙漠的甘肅省民勤縣群體種子(此后稱民勤沙米);藜麥種子,購自山西省靜樂縣,經(jīng)過發(fā)芽試驗(yàn),取發(fā)芽率和發(fā)芽勢皆高的6個不同商家的藜麥種子。2個沙米群體和藜麥種子各有6個生物學(xué)重復(fù)。
1.2 代謝組測定分析
種子代謝組的測定分析由上海阿趣生物科技有限公司完成。代謝組平臺:GC×GC-TOF MS,型號:Pegasus 4D,廠家:LECO (USA)。
基礎(chǔ)數(shù)據(jù)分析包括:數(shù)據(jù)預(yù)處理,偏最小二乘法-判別分析(PLS-DA),正交偏最小二乘法-判別分析(OPLS-DA),差異化合物篩選和鑒定,代謝通路分析。
2 結(jié)果與討論
2.1 代謝組數(shù)據(jù)可靠性分析
沙米和藜麥共18個樣本,檢測到483個峰。對第一、二主成分進(jìn)行建模分析。阿爾鄉(xiāng)沙米與民勤沙米比較的OPLS-DA得分如圖1A所示,可見2個沙米群體可以顯著分開,沙米和藜麥也能清晰分開(圖1B)。分析顯示本代謝組分析數(shù)據(jù)可靠,可以進(jìn)行代謝物及代謝通路分析。
2.2 沙米和藜麥的代謝物及代謝通路
沙米和藜麥代謝組分析鑒定出106個代謝物,其中89個代謝物定位到簡化代謝通路網(wǎng)絡(luò)(圖2),未能找到相應(yīng)的代謝通路。沙米和藜麥種子的主要代謝通路有:三羧酸循環(huán)、醣酵解/糖質(zhì)新生、淀粉與蔗糖代謝、戊糖和葡萄糖醛酸酯互變、果糖和甘露糖代謝、半乳糖代謝、嘧啶代謝、甘氨酸、絲氨酸和蘇氨酸代謝、丙氨酸、天冬氨酸和谷氨酸代謝、精氨酸和脯氨酸代謝、苯丙氨酸代謝、脂肪酸生物合成等代謝通路。
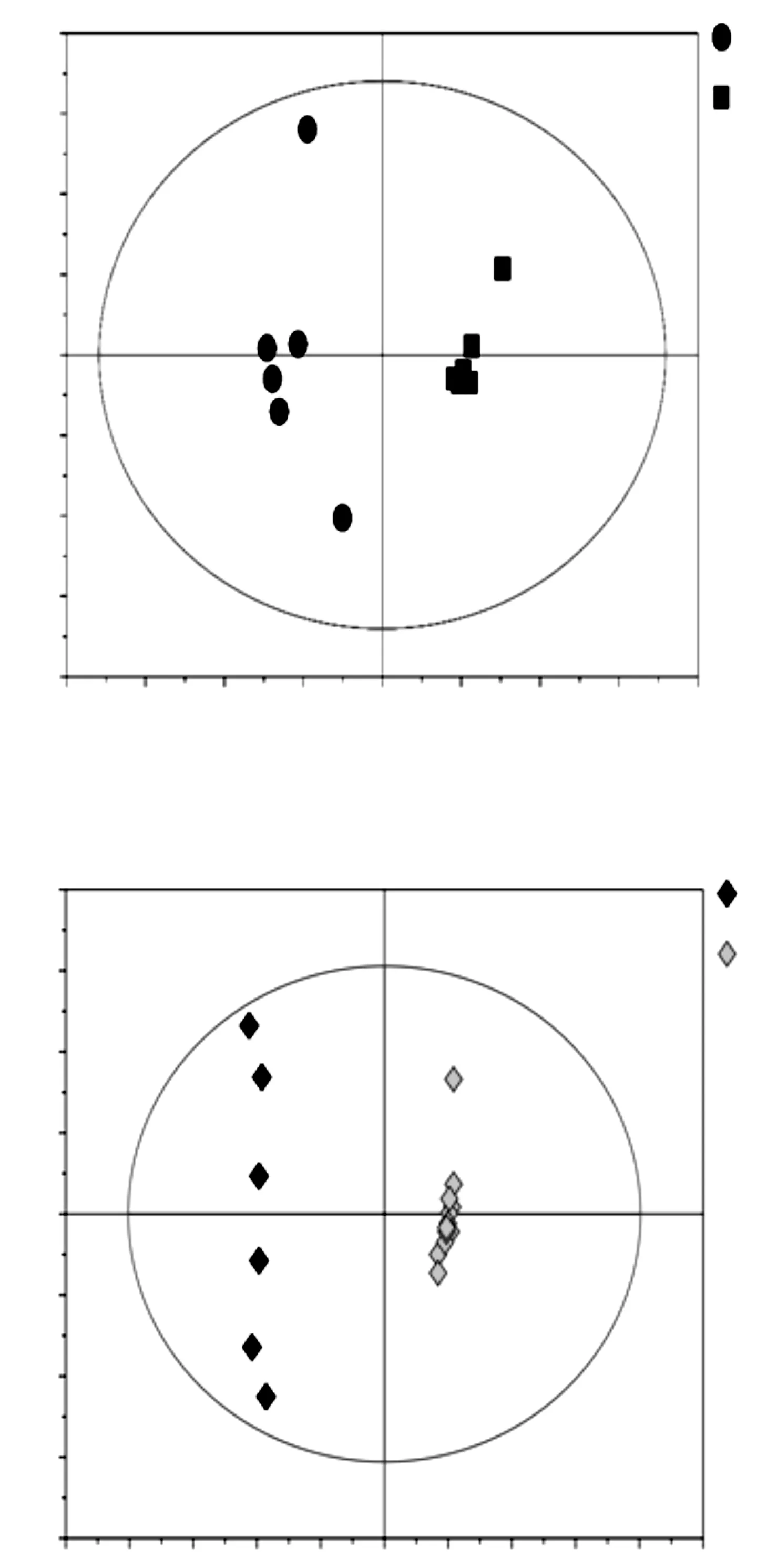
圖1 阿爾鄉(xiāng)沙米與民勤沙米(A)及沙米與藜麥(B)的OPLS-DA得分圖
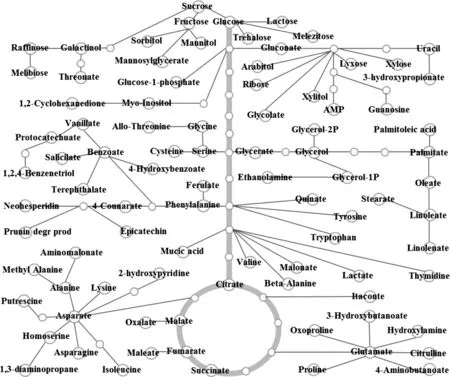
圖2 沙米和藜麥種子簡化代謝通路網(wǎng)絡(luò)
2.3 沙米及藜麥代謝組比較
科爾沁沙地的阿爾鄉(xiāng)沙米、騰格里沙漠的民勤沙米及山西省靜樂縣的藜麥代謝組都含有106個顯著代謝物,只是相對含量有差異。阿爾鄉(xiāng)沙米和民勤沙米的差異代謝物有10種(表1),其中阿爾鄉(xiāng)顯著低于民勤的代謝物有3種,阿爾鄉(xiāng)高于民勤的代謝物有7種。沙米種子有10%左右的代謝物響應(yīng)不同的生態(tài)環(huán)境,這些代謝物可能與沙米的環(huán)境適應(yīng)性有關(guān)。
沙米和藜麥的差異代謝物有23種(表2),其中沙米比藜麥高的代謝物有6種,沙米比藜麥含量低的代謝物有17種。
肌醇半乳糖苷和棉籽糖在民勤沙米和阿爾鄉(xiāng)沙米中的相對含量差異不顯著、其中沙米中的相對含量顯著大于藜麥中的相對含量。民勤處于干旱區(qū)沙漠,而阿爾鄉(xiāng)處于半干旱區(qū)沙地,藜麥?zhǔn)寝r(nóng)作物,環(huán)境條件顯著不同,由此推斷,肌醇半乳糖苷和棉籽糖是沙米適應(yīng)沙漠逆境的標(biāo)志性代謝物。肌醇半乳糖苷和棉籽糖是典型的耐逆物質(zhì),與多種耐逆響應(yīng)有關(guān)[15-19],它們不僅是滲透調(diào)節(jié)物質(zhì)和細(xì)胞膜穩(wěn)定劑,還是活性氧清除劑,可以減輕干旱、鹽及冷脅迫所引起的活性氧的危害[20],它們在擬南芥耐強(qiáng)光適應(yīng)中積累,說明其在植物耐強(qiáng)光中的作用[21]。沙米能夠適應(yīng)流動沙丘,與它的耐強(qiáng)光性有關(guān),午間可以同時受高強(qiáng)光和高溫脅迫。肌醇半乳糖苷在種子發(fā)育后期脫水過程中產(chǎn)生,其含量可以反映環(huán)境的變化[22]。由于肌醇半乳糖苷和棉籽糖的抗氧化特性,在沙米代謝組中,它們的相對含量較高,在阿爾鄉(xiāng)沙米中排名分別為第3和第5,在民勤沙米中排名分別為第1和第5,沙米應(yīng)該具有很好的保健作用。在稻米種子代謝組中肌醇半乳糖苷相對含量很低,排在最后幾個代謝物中,而棉籽糖沒有檢測到[23]。在小麥種子代謝組中沒有檢測到肌醇半乳糖苷和棉籽糖[24],可見在種子發(fā)育過程中,肌醇半乳糖苷和棉籽糖積累并不是一種普遍現(xiàn)象。
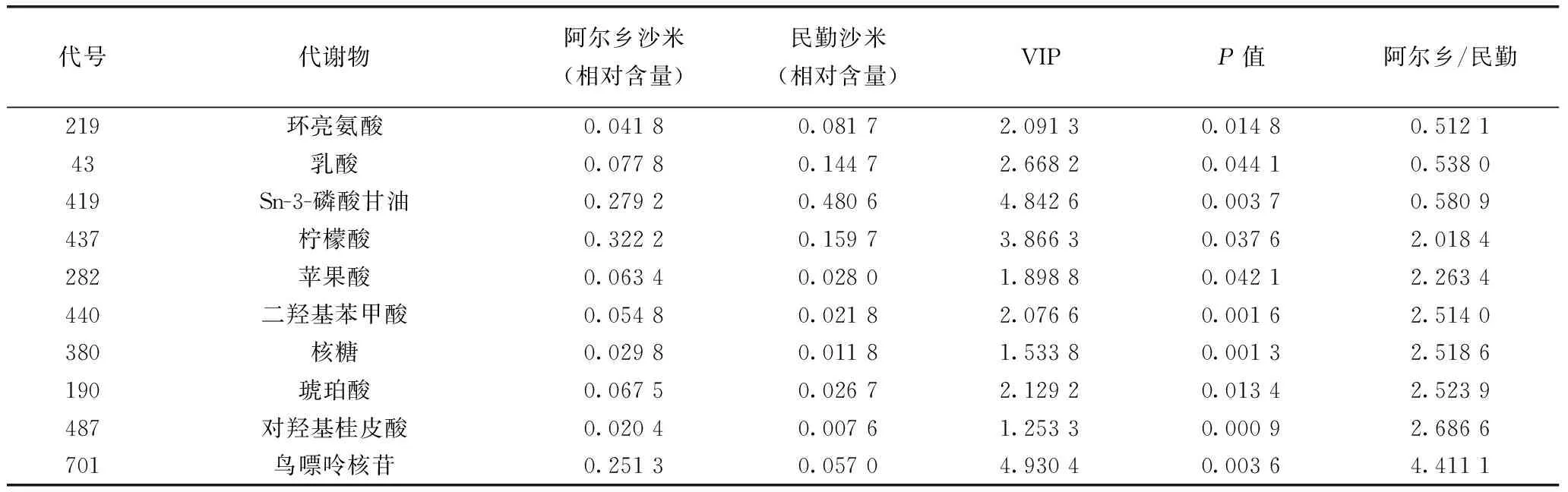
表1 阿爾鄉(xiāng)沙米和民勤沙米顯著差異代謝物
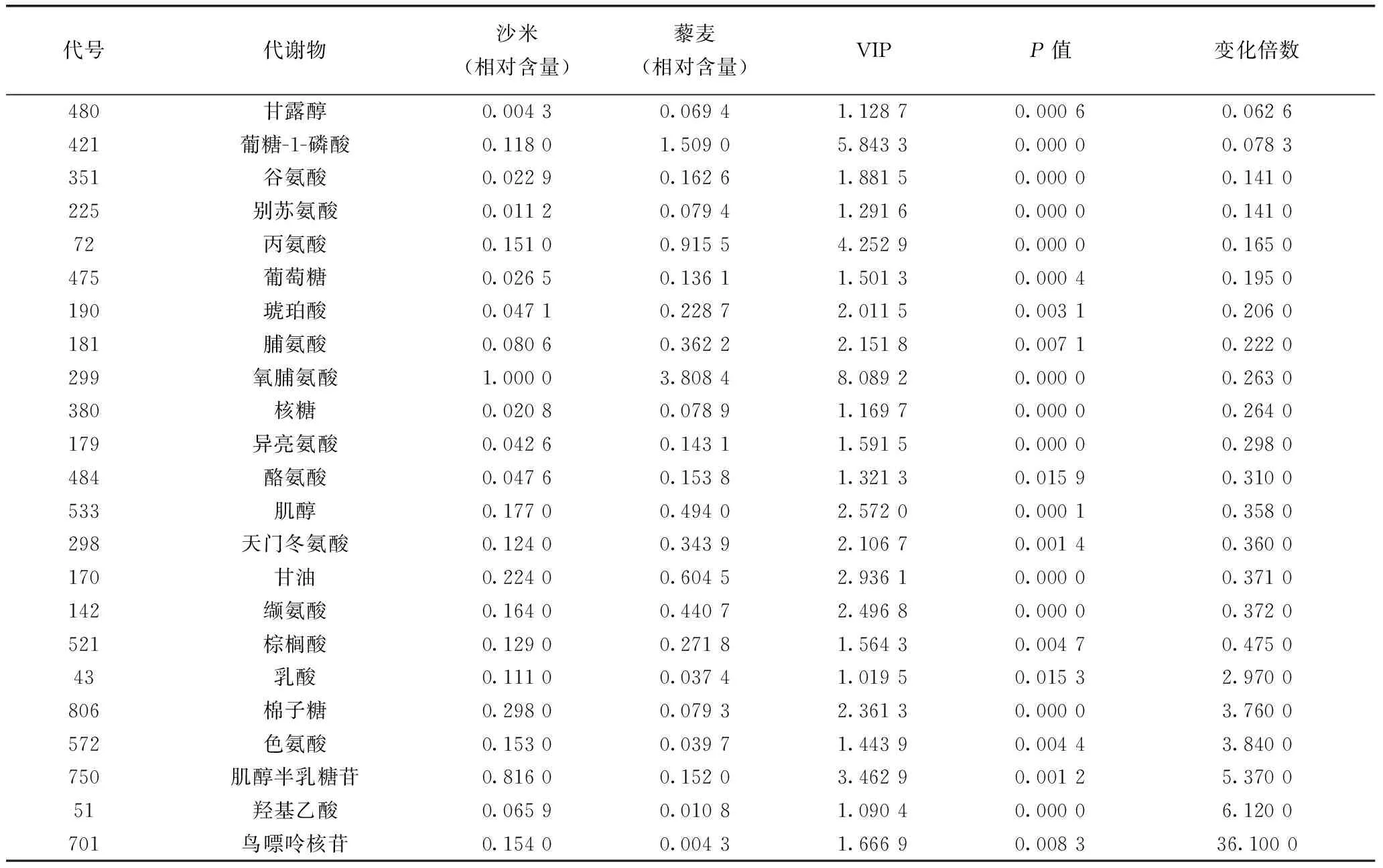
表2 沙米和藜麥顯著差異代謝物
脯氨酸相對含量在藜麥中排在前10位,在沙米中相對含量沒在前10位,處于中上水平,脯氨酸相對含量在藜麥代謝組中高于在沙米代謝組中。細(xì)菌、真菌、原生動物、海洋無脊椎動物和植物受各種脅迫后,就會發(fā)生脯氨酸積累,這些脅迫包括鹽脅迫、干旱、高溫、低溫、重金屬、病害、厭氧、營養(yǎng)缺乏、空氣污染、紫外輻射等[25]。在未受脅迫的植物中,種子的脯氨酸含量比其它器官高,脯氨酸在種子發(fā)育過程中起到滲透調(diào)節(jié)作用。脯氨酸在藜麥種子中保護(hù)作用比在沙米種子中的保護(hù)作用更重要,沙米種子的保護(hù)作用主要由肌醇半乳糖苷和棉籽糖提供。在稻米種子代謝組中[23]和小麥種子代謝組中[26]都有較高的脯氨酸含量,可見在種子發(fā)育過程中,積累脯氨酸是一種比較普遍的現(xiàn)象。
在沙米代謝組中,相對含量較高(相對含量>0.01),而在稻米和小麥代謝組中[24,25]都沒有檢測到棉籽糖、甘油磷酸、葡萄糖-1-磷酸、2-羥基吡啶、原兒茶酸、羥胺、對苯二甲酸、木糖醇。這些代謝物可能彌補(bǔ)稻米和小麥的營養(yǎng)不足,對以稻米或小麥為主食的人群來說,沙米是理想的藥食同源食物。棉籽糖是一種功能性低聚糖,在人體小腸中不降解、不吸收、能量低、有甜味,在大腸中是雙歧桿菌的增殖因子,能抑制腐敗菌產(chǎn)生的有害物質(zhì),有整腸作用,預(yù)防便秘,對雙歧桿菌有益,而雙歧桿菌可以合成維生素B1、維生素B2、維生素B6、維生素B12和煙酸及葉酸等營養(yǎng)物質(zhì),提高機(jī)體免疫力[27]。葡萄糖-1-磷酸(G1P),受葡萄糖磷酸變位酶催化,生成葡萄糖-6-磷酸,進(jìn)入糖酵解途徑[28],是重要的能量物質(zhì);G1P是糖核苷酸生物合成的主要成分,也是海藻糖合成的主要底物,能提高鈣在小腸內(nèi)的運(yùn)輸活性[29];G1P抑制細(xì)胞生長,可用于癌癥和心臟病的治療[30,31]。原兒茶酸具有增加冠脈流量、制血小板聚集和抗菌消炎作用[32,33],還有明顯的神經(jīng)保護(hù)作用[34],可作為防治帕金森病藥物中的主要成分[35]。我國“四大南藥”之一益智仁的主要成分包含原兒茶酸,具有抗癌和保護(hù)心血管等作用[36]。木糖醇,一種健康的甜味劑,是糖尿病患者最適宜的營養(yǎng)性食糖代替品,有益于肝糖元合成,可輔助治療肝炎并發(fā)癥,應(yīng)用于各種減肥食品中,因其難被細(xì)菌代謝,影響血鏈球菌的致齲性[33]。
3 結(jié)論
沙米種子與藜麥種子代謝組基本一致,證明了沙米營養(yǎng)價(jià)值可與藜麥營養(yǎng)價(jià)值相媲美。沙米種子有10%左右的代謝物響應(yīng)不同的生態(tài)環(huán)境,這些代謝物可能與沙米的環(huán)境適應(yīng)性有關(guān)。肌醇半乳糖苷和棉籽糖在民勤沙米和阿爾鄉(xiāng)沙米中的相對含量差異不顯著,沙米中的相對含量顯著大于藜麥中的相對含量,它們是沙米適應(yīng)沙漠逆境的標(biāo)志性代謝物,在農(nóng)作物種子中的積累并不是一種普遍現(xiàn)象。脯氨酸在藜麥種子中保護(hù)作用比在沙米種子中的保護(hù)作用更重要,在稻米種子和小麥種子代謝組中都有較高的脯氨酸含量,可見在種子發(fā)育過程中,積累脯氨酸是一種比較普遍的現(xiàn)象。在沙米代謝組中,相對含量較高,而在稻米和小麥代謝組中都沒有檢測到的代謝物有棉籽糖、甘油磷酸、葡萄糖-1-磷酸、2-羥基吡啶、原兒茶酸、羥胺、對苯二甲酸、木糖醇,這些代謝物可彌補(bǔ)稻米和小麥的營養(yǎng)不足。對以稻米或小麥為主食的人群來說,沙米是理想的藥食同源食物。◇
[1]Chen G X,Zhao J C,Zhao X,etal.A psammophyteAgriophyllumsquarrosum(L.)Moq.:a potential food crop [J]. Genetic Resources and Crop Evolution,2014,61:669-676.
[2]Bioversity International and FAO.Concept note:celebrating the international year of quinoa[EB/OL]. http://www.fao.org/quinoa-2013/publications/fao/concept-note-celebratingthe-%20international-year-of-quinoa/ar/?no_mobile=1,2012.
[3]許國旺,路鑫,楊勝利.代謝組學(xué)研究進(jìn)展[J]. 中國醫(yī)學(xué)科學(xué)院學(xué)報(bào),2007,29:701-711.
[4]李寧,范雪梅,王義明,等.代謝組學(xué)及其分析技術(shù)的研究進(jìn)展[J]. 中南藥學(xué),2014,12:668-673.
[5]許國旺,楊軍.代謝組學(xué)及其研究進(jìn)展[J]. 色譜,2003,21:316-320.
[6]邱緒建,耿偉,劉光明,等.代謝組學(xué)方法在食品安全中的應(yīng)用研究進(jìn)展[J]. 食品工業(yè)科技,2012,33:369-373.
[7]Dclarke J,Calexander D,Pward D,etal.Assessment of Genetically Modified Soybean in Relation to Natural Variation in the Soybean Seed Metabolome [J]. Scientific Reports,2013,3:3082.
[8]Millar K,Jgibson D,Gyoung B,etal.Impact of interspecific competition on seed development and quality of five soybean cultivars [J]. Animal Production Science,2007,47:1455-1459.
[9]Lin H,Rao J,Shi J,etal.Seed metabolomic study reveals significant metabolite variations and correlations among different soybean cultivars [J]. Journal of Integrative Plant Biology,2014,56:826-836.
[10]Zhen S X,Dong K,Xiong D,etal.Dynamic metabolome profiling reveals significant metabolic changes during grain development of bread wheat (TriticumaestivumL.)[J]. Journal of the Science of Food & Agriculture,2016,doi:10:1002/jsfa.7561.
[11]De Leonardis A M,F(xiàn)ragasso M,Beleggia R,etal.Effects of Heat Stress on Metabolite Accumulation and Composition,and Nutritional Properties of Durum Wheat Grain [J]. International Journal of Molecular Sciences,2015,16:30382-30404.
[12]Nam K,Jaeshin H,Pack I,etal.Metabolomic changes in grains of well-watered and drought‐stressed transgenic rice [J]. Journal of the Science of Food and Agriculture,2016,96:807-814.
[13]Nam M,Bang E,Kwon T,etal.Metabolite Profiling of Diverse Rice Germplasm and Identification of Conserved Metabolic Markers of Rice Roots in Response to Long-Term Mild Salinity Stress [J]. International Journal of Molecular Sciences,2015,16:21959-21974.
[14]Gorzolka K,Lissel M,Kessler N,etal.Metabolite fingerprinting of barley whole seeds,endosperms,and embryos during industrial malting [J]. Journal of Biotechnology,2012,159:177.
[15]Taji T,Ohsumi C,Iuchi S,etal.Important roles of drought and cold inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana [J]. Plant Journal,2002,29:417-426.
[16]Panikulangara T,Eggersschumacher G,Wunderlich M,etal.Galactinol synthase1.A Novel Heat Shock Factor Target Gene Responsible for Heat-Induced Synthesis of Raffinose Family Oligosaccharides in Arabidopsis [J]. Plant Physiology,2004,136:3148.
[17]Kaplan F,Kopka J,Haskell D,etal.Exploring the Temperature-Stress Metabolome of Arabidopsis [J]. Plant Physiology,2004,136:4159-4168.
[18]Kaplan F,Kopka J,Sung D,etal.Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content [J]. Plant Journal,2007,50:967-981.
[19]Peters S,Mundree S,Thomson J,etal.Protection mechanisms in the resurrection plant Xerophyta viscosa (Baker):both sucrose and raffinose family oligosaccharides (RFOs)accumulate in leaves in response to water deficit [J]. Journal of Experimental Botany,2007,58:1947-1956.
[20]Nishizawa A,Yabuta Y,Yoshida E,etal.Arabidopsis heat shock transcription factor A2 as a key regulator in response to several types of environmental stress [J]. Plant Journal,2006,48:535-547.
[21]Jankanpaa H,Mishra Y,Schroder W,etal.Metabolic profiling reveals metabolic shifts in Arabidopsis plants grown under different light conditions [J]. Plant Cell and Environment,2012,35:1824-1836.
[22]Clarke S,Kingston J,James K,etal.Acceptance and Commitment Therapy group for treatment-resistant participants:A randomized controlled trial [J]. Journal of Contextual Behavioral Science,2014,3:179-188.
[23]Hu C,Tohge T,Chan S A,etal.Identification of Conserved and Diverse Metabolic Shifts during Rice Grain Development [J]. Scientific Reports,2016,6:20942.
[24]M.Francki,S.Hayton,J.Gummer,etal.Metabolomic profiling and genomic analysis of wheat aneuploid lines to identify genes controlling biochemical pathways in mature grain [J]. Plant Biotechnology Journal,2015,14:649-660.
[25]N.Verbruggen,C.Hermans.Proline accumulation in plants:a review [J]. Amino Acids,2008,35:753-759.
[26]Zhen S,Dong K,Deng X,etal.Dynamic metabolome profiling reveals significant metabolic changes during grain development of bread wheat (TriticumaestivumL.)[J]. Journal of the Science of Food and Agriculture,2016,doi:10:1002/jsfa.7561.
[27]袁美蘭,溫輝梁,黃紹華.功能性低聚糖-棉籽糖的開發(fā)應(yīng)用現(xiàn)狀[J]. 中國食品添加劑,2002,9:54-57.
[28]Kitaoka M,Hayashi K.Carbohydrate-processing phosphorolytic enzymes [J]. Trends in Glycoscience & Glycotechnology,2002,14:35-50.
[29]Fraser D,Kodicek E.Unique Biosynthesis by Kidney of a Biologically Active Vitamin D Metabolite [J]. Nature,1970,228:764-766.
[30]Vandamme,E Van Loo J,Machtelinckx L,etal.Microbial Sucrose Phosphorylase:Fermentation Process,Properties,and Biotechnical Applications [J]. Advances in Applied Microbiology,1987,32:163-201.
[31]Parish C,Cowden WB,Willenborg DO.Phosphosugar-based anti-inflammatory and/or immunosuppressive drugs [P]. 1996,Patent US5506210.
[32]徐宗佩,張伯禮,李尚珠,等.中藥單體原兒茶醛對瘀血證患者單核細(xì)胞趨化游走能力的影響[J]. 天津中醫(yī),2002,26:49-50.
[33]趙光潔,趙燕霞,姜新亞,等.蔗糖及木糖醇對血鏈球菌和變異鏈球菌相互作用的影響[J]. 牙體牙髓牙周病學(xué)雜志,2016,(01):7-11,20.
[34]An L,Guan S,Shi G,etal.Protocatechuic acid fromAlpiniaoxyphyllaagainst MPP+-induced neurotoxicity in PC12 cells [J]. Food and Chemical Toxicology,2006,44:436.
[35]劉葉明,鄭甜甜,徐美娟,等.益智仁中的原兒茶酸對魚藤酮損傷PC12細(xì)胞的保護(hù)作用[J]. 遼寧大學(xué)學(xué)報(bào)(自然科學(xué)版),2016,43:61-67.
[36]代汝偉,章時杰,蔡偉彬,等.原兒茶酸對β淀粉樣蛋白誘導(dǎo)PC12細(xì)胞毒性的保護(hù)作用及機(jī)制[J]. 廣州中醫(yī)藥大學(xué)學(xué)報(bào),2016,33:66-70.
(責(zé)任編輯 李婷婷)
Comparative Analysis on Metabolomics of Agriophyllum squarrosum and Chenopodium quinoa Seeds
ZHAO Jie-cai,MA Yan,ZHOU Qin,ZHAO Xin,MA Xiao-fei,CHEN Guo-xiong
(Cold and Arid Regions Environmental and Engineering Research Institute,Chinese Academy of Sciences/Shapotou Desert Experiment and Research Station/Key Laboratory of Stress Physiology and Ecology in Cold and Arid Regions in Gansu Province,Lanzhou 730000,China)
To understand seed metabolites ofAgriophyllumsquarrosumandChenopodiumquinoa,untargeted metabolomics analysis of sand rice and quinoa seeds had been conducted using GC×GC-TOF MS platform.A total of 106 metabolites were identified in sand rice seeds harvested from Horqin sandy land and Tengger Desert and Quinoa seeds produced in Jingle,Shanxi,China.The metabolites had similar relative contents between sand rice and quinoa except of six metabolites higher and 17 metabolites lower in sand rice than in quinoa.Some metabolites such as raffinose,dihydroxybenzoic acid,and xylitol had a high content in sand rice but could not be identified in rice and wheat.These metabolites may complement the lack of nutrients in rice and wheat.Sand rice is a perfect affinal drug and diet for people who eat mainly rice or wheat.
Agriophyllumsquarrosum;Chenopodiumquinoa;metabolite;affinal drug and diet;active sand dune
國家重點(diǎn)基礎(chǔ)研究發(fā)展計(jì)劃(項(xiàng)目編號:2013CB429904);中國科學(xué)院寒區(qū)旱區(qū)環(huán)境與工程研究所成果轉(zhuǎn)化引導(dǎo)基金(項(xiàng)目編號:Y655831001)。
趙杰才(1986— ),男,在讀博士研究生,研究方向:植物生理生態(tài)學(xué)。
陳國雄(1963— ),男,博士,研究員,研究方向:植物生理生態(tài)學(xué)。
