叉頭框蛋白3過表達對胃癌細胞生物學行為的影響
2016-12-16 03:19:05馬桂芬潘獨伊曾昭沖陳世耀
中國臨床醫學 2016年5期
關鍵詞:胃癌
馬桂芬, 潘獨伊, 何 健, 曾昭沖, 陳世耀*
1.復旦大學附屬中山醫院放療科, 上海 200032 2.復旦大學附屬中山醫院消化內科, 上海 200032
?
·論 著·
叉頭框蛋白3過表達對胃癌細胞生物學行為的影響
馬桂芬1, 潘獨伊2, 何 健1, 曾昭沖1, 陳世耀2*
1.復旦大學附屬中山醫院放療科, 上海 200032 2.復旦大學附屬中山醫院消化內科, 上海 200032
目的: 探討叉頭框蛋白3(Forkhead box protein 3,FoxP3)過表達對胃癌細胞生物學行為的影響。方法: 用shRNA過表達質粒轉染胃癌細胞,經藥物篩選和單克隆挑選,建立穩定轉染的細胞系。用四甲基偶氮唑鹽(MTT)法檢測轉染FoxP3基因后胃癌細胞系生長增殖能力的改變;流式細胞術檢測細胞周期;MTT法測定細胞轉染FoxP3基因后對化療藥物敏感性的影響。結果: 成功建立穩定轉染FoxP3基因的胃癌細胞。與空載體轉染的對照組相比,上調FoxP3基因的胃癌細胞生長增殖能力減慢,遷移能力減弱[穿膜細胞數:(203±42)個/HPvs(891±100)個/HP,P<0.05],對化療藥物的敏感性增加(P<0.05)。結論: 過表達FoxP3基因可抑制胃癌細胞的增殖、遷移,提高化療敏感性。
FoxP3; 胃癌; 基因功能; 生物學行為
2012年全球惡性腫瘤統計中,胃癌發病率排名第5位,在亞洲國家發病率更高達24.1 /10萬,死亡率居第3位[1]。我國現有胃癌患者約300萬,嚴重威脅人們的健康。目前對胃癌主要采用以手術為主,放療、化療為輔的綜合治療,雖然療效較前有很大的改善,但中位生存期僅14.6個月,3年生存率僅27%[2]。分子靶向治療能從根本上改變癌癥的發生發展,糾正患者機體的促癌狀態,有望成為有效的胃癌治療新途徑。
人類叉頭框蛋白3(Forkhead box protein 3, FoxP3)基因突變會導致致死性疾病,如免疫功能紊亂(immune dysregulation)、多腺體病(polyendocrinopathy)、腸病(enteropathy)和X染色體連鎖綜合征(X-linked syndrome,IPEX綜合征)[3]。近年來發現,FoxP3表達于腫瘤細胞中,可能是一種新的腫瘤調控因子。在腫瘤微環境中,腫瘤細胞可能通過表達FoxP3 抑制免疫活性,以此來模擬Treg細胞的功能[4]。FoxP3陽性的胰腺癌細胞系可抑制CD4+T細胞的擴增,可能與腫瘤細胞免疫逃逸有關[5]。目前大部分腫瘤中FoxP3表達增加,如小細胞肺癌[6]、肝癌[7]和胃癌[8]等。然而FoxP3基因對胃癌細胞生物學行為的影響目前尚不清楚。
1 材料與方法
1.1 重組質粒DNA轉染胃癌細胞 AGS胃癌細胞株購于中國科學院上海生命科學院細胞庫。用含10% 胎牛血清(FBS)及100 U/mL青霉素和0.1 mg/mL鏈霉素的DMEM完全培養基(美國Gibco公司)于37℃、5% CO2的培養箱中培養。取對數生長期的細胞,調整密度至2×105個/mL,接種至12孔培養板,培養至細胞融合度達70%~75%。將pEGFP/FoxP3和pEGFP/vector shRNA質粒(由上海吉凱基因化學技術有限公司構建)分別與Lipofectamine 2000 (美國Invitrogen公司) 混合后,用Opti-MEMI無血清培養基(美國Gibco公司)稀釋至50 μL。將混合液加入培養板進行轉染,6 h后換液,轉染48 h后,收集細胞,按1∶10接種于培養皿中,使用含G418的選擇培養基進行抗性篩選,每3 d更換培養基。3周后挑取細胞克隆,擴大培養。轉染成功的陽性細胞克隆在熒光顯微鏡下可觀察到綠色熒光,挑選出增強型綠色熒光蛋白(enhanced green fluorescent protein,EGFP)表達最強的單個細胞克隆。進一步采用Western 印跡法選取FoxP3蛋白表達最強的克隆,擴大培養后用于后續實驗。
1.2 流式細胞術檢測細胞周期 細胞周期試劑盒購于南京凱基生物科技發展有限公司。取對數生長期細胞,倒去培養液,胰酶適度消化,用培養液吹打,250 ×g離心 15 min,去上清;PBS洗2次,加0.5 mL 磷酸緩沖鹽溶液(PBS)混勻,務必吹散;用5 mL注射器將細胞吸起,用力打入5 mL 70%乙醇中,固定過夜;第2天收集固定的細胞,PBS洗2次;加RNase A約3 μL至終濃度約為50 μg/mL,37℃水浴消化30 min;加碘化丙啶(PI)約50 μL至終濃度約為65 μg/mL,在冰浴中避光染色30 min;用流式細胞儀(美國BD公司)檢測細胞周期。
1.3 四甲基偶氮唑鹽(MTT)法檢測細胞增殖能力 MTT檢測試劑盒購于南京凱基生物科技發展有限公司。分別取對數生長期的pEGFP/FoxP3及pEGFP/vector轉染后的AGS細胞,調整細胞密度為104個/mL,接種至96孔板,每孔200 μL,每組設3~5個復孔;分別培養24、48、72、96 h后,每孔加入5 mg/mL濃度的MTT試劑 20 μL,繼續培養4 h,棄培養液,加入150 μL DMSO,振蕩后用酶標儀檢測550 nm處光密度(D)值,以24 h為基準,計算以后每天的增長倍數。
1.4 細胞遷移實驗 分別取對數生長期的AGS/FoxP3和AGS/vector細胞,調整其密度至5×105個/mL,接種至Transwell小室內(美國Corning公司),每孔200 μL;在下室加入0.5 mL含10%FBS的RPMI 1640完全培養基,放入培養箱培養;24 h后取出Transwell小室,用棉簽拭去上室內細胞,置于預冷的無水乙醇中固定10 min;PBS洗滌3次后將Transwell小室浸入吉姆薩染色液中染色10 min,洗滌后用刀片小心割下膜,置于載玻片上,用中性樹膠封片;在顯微鏡下,隨機選取5個視野(100×),計數穿膜細胞個數。
1.5 化療藥物藥敏試驗 分別取對數生長期的AGS/FoxP3和AGS/vector細胞,計數,接種至96孔板,每孔5 000個細胞。每組設5個復孔。細胞貼壁8 h后,分別加入不同濃度的5-FU或伊立替康;48 h后用MTT法在550 nm波長處檢測D值。
1.6 統計學處理 采用SPSS 17.0進行統計分析。滿足正態性分布和方差齊性的兩組間比較采用t檢驗,不滿足者采用非參數Kruskal-Wallis檢驗。檢驗水準(α)為0.05。
2 結 果
2.1 單克隆挑選及過表達載體的鑒定 經Western 印跡驗證,選取蛋白表達最強的5號克隆(圖1)擴大培養用于后續試驗。

圖1 篩選FoxP3蛋白過表達最強克隆
2.2 流式細胞術鑒定細胞轉染效率 經過 G418篩選和單克隆挑選擴增的穩定轉染的AGS細胞(圖2A),熒光顯微鏡下見細胞均帶有綠色熒光(圖2B),說明轉染成功,流式細胞技術顯示轉染效率近100%(圖2C)。
2.3 過表達FoxP3對胃癌細胞增殖能力的影響 細胞增殖實驗均顯示轉染FoxP3質粒后,AGS細胞的增殖能力減低(圖3),提示AGS/FoxP3具有抑制胃癌細胞增殖的作用(P<0.05)。

圖2 FoxP3穩定轉染細胞系的篩選
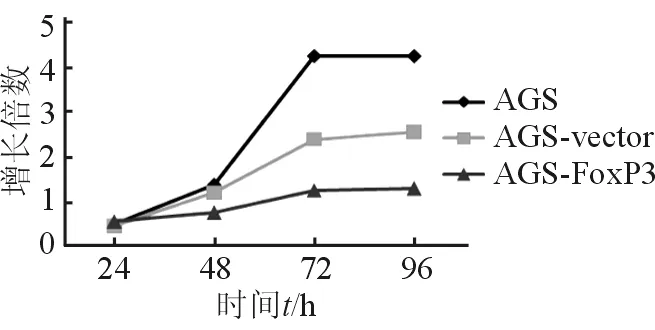
圖3 MTT示AGS/FoxP3抑制AGS細胞生長
2.4 細胞周期的變化 流式細胞術分析結果(圖4)表明:轉染FoxP3后,細胞被阻滯在G1期,G1期細胞所占百分比明顯多于未轉染vector的對照組[G1期:(85.21±10.21)%vs(72.12±7.34)%,P<0.05]。

圖4 流式細胞術提示細胞周期被阻滯在G1期
2.5 轉染FoxP3對胃癌細胞遷移能力的影響 細胞遷移實驗(圖5)表明:與對照組相比,轉染FoxP3的AGS細胞在高倍視野中平均穿膜細胞數明顯減少[(203±42)個/HPvs(891±100)個/HP,P<0.05],說明轉染FoxP3后胃癌細胞的遷移能力減弱。

圖5 轉染FoxP3胃癌細胞的遷移能力減弱
2.6 轉染后FoxP3對胃癌細胞化療敏感性的影響 在5-FU 100 μg/mL和500 μg/mL濃度下,轉染FoxP3的AGS細胞生長抑制率明顯高于對照組;在伊立替康25 μg/mL及50 μg/mL藥物濃度下,轉染FoxP3基因的AGS細胞的生長抑制率明顯升高(P<0.05)。結果表明,FoxP3可以增加胃癌細胞對化療藥物的敏感性(圖6)。
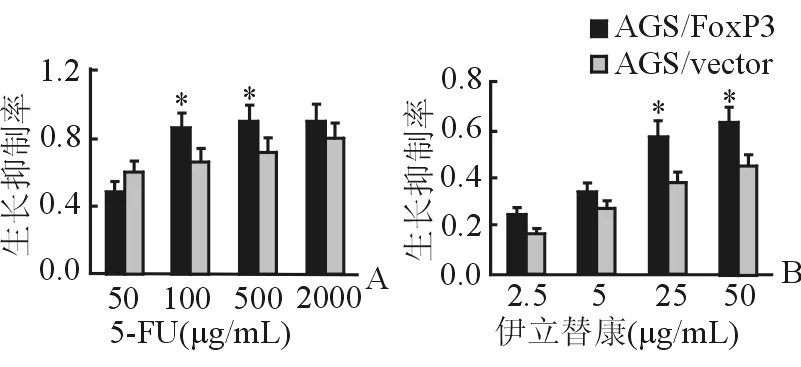
圖6 化療藥物敏感性的增加
3 討 論
FoxP3對Treg細胞的形成和功能起關鍵作用[9]。FoxP3陽性的T細胞大部分為Treg,它們是構成腫瘤免疫的重要因素。有研究[10]報道,胰腺癌新輔助放化療后腫瘤微環境中Treg減少及相關的細胞毒作用減弱。放療可使循環中的Treg明顯下降,患者無瘤生存時間延長[11-12]。但是,有證據[13]發現,Treg比細胞毒性T淋巴細胞(cytotoxic T lymphocyte, CTL) 更耐受離子照射,放療后局部原位組織中Treg不減少是致放療耐受的主要原因。在動物模型中也發現,Treg比其他淋巴細胞更耐受放療,他們可能在放療相關的組織損傷中有自穩態作用,導致放療不敏感[14]。與FoxP3相互作用的蛋白質中有視黃酸受體孤兒受體樣受體α(ROR-α),FoxP3可以抑制ROR-α介導的轉錄激活作用[15]。當缺乏配體時,ROR-α可以與轉錄共抑制因子結合;當與配體結合后,ROR-α發生形變與共活化因子相結合[16],從而調節其轉錄抑制或活化作用。
近來發現多種腫瘤細胞表達FoxP3,且FoxP3與淋巴結轉移和預后有重要關系[17-19]。在亞洲胃癌患者中,腫瘤組織中浸潤的CD3、CD8、CD45RO、FoxP3等免疫分子的表達與預后有關,說明免疫狀態水平可影響患者預后[20]。既往研究多集中于Treg在腫瘤免疫中的作用,而忽視了腫瘤細胞表達FoxP3。腫瘤細胞表達的FoxP3可通過影響T淋巴細胞的分化而影響腫瘤免疫應答,如結腸癌細胞中,FoxP3與Treg發生相互作用,影響幼稚T淋巴細胞的分化,從而影響免疫效應[21];肝癌細胞可通過分泌轉化生長因子β1(TGF-β1)誘導淋巴細胞中FoxP3的表達,從而抑制其抗腫瘤免疫反應[22]。我們的前期研究[23]也發現,將胃癌細胞與外周單個核淋巴細胞(PBMC)共培養后,FoxP3增多;而Treg也通過分泌抑制性細胞因子抑制胃癌細胞的生長。我們前期研究[24]發現,胃癌組織中胃癌細胞和淋巴細胞中均表達FoxP3,而且胃癌細胞FoxP3的表達與預后良好有關,而高密度浸潤的Treg與預后差有關,風險模型分析提示Treg是影響預后的主要因素。然而,FoxP3基因對胃癌細胞具體生物學行為的影響目前還不清楚。
乳腺癌體內外實驗[25-27]均表明,FoxP3能抑制乳腺癌的生長。乳腺癌細胞系過表達FoxP3基因后,細胞增殖、遷移和侵襲能力均降低[25]。FoxP3可通過抑制人表皮生長因子受體2(HER2)和S相激酶相關蛋白2(SKP2)的轉錄活性和誘導腫瘤抑制基因p21的活性抑制乳腺癌的生長[28-29]。前列腺癌細胞系增加FoxP3表達后生長明顯抑制。另外敲除FoxP3會導致早發的前列腺增生和上皮內瘤變[30]。本實驗發現過表達FoxP3的胃癌細胞增殖減慢、遷移能力減弱,胃癌細胞主要被阻滯在G1期。我們以往研究[31]還發現,FoxP3促進胃癌細胞的凋亡與Caspase相關的凋亡信號通路有關,因此推測FoxP3在胃癌中可能起抑癌作用。本研究發現,轉染FoxP3的胃癌細胞對5-FU和伊立替康兩種化療藥物的敏感性增加,這可能也是FoxP3影響患者預后的原因之一。Treg的免疫抑制作用是影響預后的重要因素,而胃癌細胞中FoxP3起抑癌作用,它們可能共同參與調節腫瘤微環境的狀態,最終決定患者預后。
[ 1 ] Ferlay J, Soerjomataram I, Dikshit R, et al. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012[J]. Int J Cancer,2015,136(5):e359-e386.
[ 2 ] Yoshikawa T, Sasako M, Yamamoto S, et al. Phase Ⅱ study of neoadjuvant chemotherapy and extended surgery for locally advanced gastric cancer[J]. Br J Surg,2009,96(9):1015-1022.
[ 3 ] Torgerson TR, Ochs HD. Immune dysregulation, polyendocrinopathy, enteropathy, X-linked: Forkhead box protein 3 mutations and lack of regulatory T cells[J]. J Allergy Clin Immunol,2007,120(4):744-750.
[ 4 ] 薛 劍,李美星,袁向亮,等. Foxp3在胃癌細胞中的表達[J]. 診斷學理論與實踐,2009,8(2):160-164.
[ 5 ] Hinz S, Pagerols-Raluy L, Oberg HH, et al. Foxp3 expression in pancreatic carcinoma cells as a novel mechanism of immune evasion in cancer[J]. Cancer Res,2007,67(17):8344-8350.
[ 6 ] Dimitrakopoulos FI, Papadaki H, Antonacopoulou AG, et al. Association of FOXP3 expression with non-small cell lung cancer[J]. Anticancer Res,2011,31(5):1677-1683.
[ 7 ] Wang WH, Jiang CL, Yan W, et al. FOXP3 expression and clinical characteristics of hepatocellular carcinoma[J]. World J Gastroenterol,2010,16(43):5502-5509.
[ 8 ] 袁甲翔,張謝夫,趙春臨,等. 核轉錄因子FOXP3在胃癌患者體內的表達及意義[J]. 世界華人消化雜志,2009,17(29):3052-3055.
[ 9 ] Brunkow ME, Jeffery EW, Hjerrild KA, et al. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse[J]. Nat Genet,2001,27(1):68-73.
[10] Tsuchikawa T, Hirano S, Tanaka E, et al. Novel aspects of preoperative chemoradiation therapy improving anti-tumor immunity in pancreatic cancer[J]. Cancer Sci,2013,104(5):531-535.
[11] Schmidt MA, Fortsch C, Schmidt M, et al. Circulating regulatory T cells of cancer patients receiving radiochemotherapy may be useful to individualize cancer treatment[J]. Radiother Oncol,2012,104(1):131-138.
[12] Tabachnyk M, Distel LV, Buttner M, et al. Radiochemotherapy induces a favourable tumour infiltrating inflammatory cell profile in head and neck cancer[J]. Oral Oncol,2012,48(7):594-601.
[13] Qinfeng S, Depu W, Xiaofeng Y, et al. In situ observation of the effects of local irradiation on cytotoxic and regulatory T lymphocytes in cervical cancer tissue[J]. Radiat Res,2013,179(5):584-589.
[14] Kachikwu EL, Iwamoto KS, Liao YP, et al. Radiation enhances regulatory T cell representation[J]. Int J Radiat Oncol Biol Phys,2011,81(4):1128-1135.
[15] Du J, Huang C, Zhou B, et al. Isoform-specific inhibition of ROR alpha-mediated transcriptional activation by human FOXP3[J]. J Immunol,2008,180(7):4785-4792.
[16] Atkins GB, Hu X, Guenther MG, et al. Coactivators for the orphan nuclear receptor RORalpha[J]. Mol Endocrinol,1999,13(9):1550-1557.
[17] Douglass S, Meeson AP, Overbeck-Zubrzycka D, et al. Breast cancer metastasis: demonstration that FOXP3 regulates CXCR4 expression and the response to CXCL12[J]. J Pathol,2014,234(1):74-85.
[18] Ganapathi SK, Beggs AD, Hodgson SV, et al. Expression and DNA methylation of TNF, IFNG and FOXP3 in colorectal cancer and their prognostic significance[J]. Br J Cancer,2014,111(8):1581-1589.
[19] Triulzi T, Tagliabue E, Balsari A, et al. FOXP3 expression in tumor cells and implications for cancer progression[J]. J Cell Physiol,2013,228(1):30-35.
[20] Lin SJ, Gagnon-Bartsch JA, Tan IB, et al. Signatures of tumour immunity distinguish Asian and non-Asian gastric adenocarcinomas[J]. Gut,2015,64(11): 1721-1731.
[21] Grimmig T, Kim M, Germer CT, et al. The role of FOXP3 in disease progression in colorectal cancer patients[J]. Oncoimmunology,2013,2(6):e24521.
[22] Wang Y, Deng B, Tang W, et al. TGF-beta1 secreted by hepatocellular carcinoma induces the expression of the Foxp3 gene and suppresses antitumor immunity in the tumor microenvironment[J]. Dig Dis Sci,2013,58(6):1644-1652.
[23] Ma GF, Miao Q, Zeng XQ, et al. Transforming growth factor-b1 and -b2 in gastric precancer and cancer and roles in tumor-cell interactions with peripheral blood mononuclear cells in vitro[J]. PLoS ONE,2013,8(1):e54249.
[24] Ma GF, Miao Q, Liu YM, et al. High FoxP3 expression in tumour cells predicts better survival in gastric cancer and its role in tumour microenvironment[J]. Br J Cancer,2014,110(6):1552-1560.
[25] Zhang HY, Sun H. Up-regulation of Foxp3 inhibits cell proliferation, migration and invasion in epithelial ovarian cancer[J]. Cancer Lett,2010,287(1):91-97.
[26] Liu Y, Wang Y, Li W, et al. Activating transcription factor 2 and c-Jun-mediated induction of FoxP3 for experimental therapy of mammary tumor in the mouse[J]. Cancer Res,2009,69(14):5954-5960.
[27] Li W, Wang L, Katoh H, et al. Identification of a tumor suppressor relay between the FOXP3 and the Hippo pathways in breast and prostate cancers[J]. Cancer Res,2011,71(6):2162-2171.
[28] Liu R, Wang L, Chen G, et al. FOXP3 up-regulates p21 expression by site-specific inhibition of histone deacetylase 2/histone deacetylase 4 association to the locus[J]. Cancer Res,2009,69(6):2252-2259.
[29] Zuo T, Wang LZ, Morrison C. FOXP3 is an X-linked breast cancer suppressor gene and an important repressor of the HER-2/ErbB2 oncogene[J]. Cell,2007,129(7):1275-1286.
[30] Wang L, Liu R, Li W, et al. Somatic single hits inactivate the X-linked tumor suppressor FOXP3 in the prostate[J]. Cancer Cell,2009,16(4):336-346.
[31] Ma GF, Chen SY, Sun ZR, et al. FoxP3 inhibits proliferation and induces apoptosis of gastric cancer cells by activating the apoptotic signaling pathway[J]. Biochem Biophys Res Commun,2013,430(2):804-809.
[本文編輯] 姬靜芳
The role and mechanism of FoxP3 in biological behavior of gastric cancer cells
MA Gui-fen1, PAN Du-yi2, HE Jian1, ZENG Zhao-chong1, CHEN Shi-yao2*
1. Department of Radiotherapy, Zhongshan Hospital, Fudan University, Shanghai 200032, China 2. Department of Gastroenterology, Zhongshan Hospital, Fudan University, Shanghai 200032, China
Objective: To explore the role and mechanism of Forkhead box protein 3 (FoxP3) in biological behavior of gastric cancer (GC) cells.Methods:Invitro, the plasmid of FoxP3-shRNA was transfected into GC cells, and then the stably transfected cells were established by drug-screening and monoclone-selection. MTT assay was used to detect the growth and proliferation of GC cells. And cell cycle was detected by flow cytometry. MTT assay was used to measure the difference regarding sensitivity to chemotherapy drugs.Results: GC cell line which was stably transfected with FoxP3 gene, was established. GC cell line with up-regulation of FoxP3 gene, compared to the vector-transfected control, showed slower growth and proliferation rate, weaker ability of invasion (transmembrane cell counts: [203±42] cells/HPvs[891±100] cells/HP,P<0.05), and higher sensitivity to chemotherapy drugs(P<0.05).Conclusions: FoxP3 gene plays a role in inhibiting the growth of GC cells.
Forkhead box protein; gastric cancer; gene function; biological behavior
2015-12-22 [接受日期] 2016-05-19
國家自然科學基金青年科學基金項目(81502005),上海市自然科學基金(14ZR1406600). Supported by Youth Program of National Natural Science Foundation of China (8150200) and Natural Science Foundation of Shanghai (14ZR1406600).
馬桂芬,碩士,住院醫師. E-mail:ma.guifen@zs-hospital.sh.cn
*通信作者(Corresponding author). Tel: 021-64041990, E-mail: chen.shiyao@zs-hospital.sh.cn
10.12025/j.issn.1008-6358.2016.20160931
R 735.2
A
猜你喜歡
昆明醫科大學學報(2022年1期)2022-02-28 07:43:36
昆明醫科大學學報(2021年5期)2021-07-22 07:32:22
基層中醫藥(2020年2期)2020-07-27 02:46:06
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
中外醫療(2015年18期)2016-01-04 06:51:55
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:25
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
