蔥黃寡毛跳甲成蟲(chóng)觸角的超微形態(tài)及感器類型
2016-12-13 08:31:27張春妮
西北農(nóng)業(yè)學(xué)報(bào) 2016年10期
張 琪,陳 帆,張春妮,戴 武
(植保資源與病蟲(chóng)害治理教育部重點(diǎn)實(shí)驗(yàn)室,西北農(nóng)林科技大學(xué) 植物保護(hù)學(xué)院,陜西楊凌 712100)
?
蔥黃寡毛跳甲成蟲(chóng)觸角的超微形態(tài)及感器類型
張 琪,陳 帆,張春妮,戴 武
(植保資源與病蟲(chóng)害治理教育部重點(diǎn)實(shí)驗(yàn)室,西北農(nóng)林科技大學(xué) 植物保護(hù)學(xué)院,陜西楊凌 712100)
蔥黃寡毛跳甲LuperomorphasuturalisChen是一種重要的農(nóng)業(yè)害蟲(chóng),主要危害蔥、韭菜等作物。為了明確蔥黃寡毛跳甲觸角形態(tài)及感器類型,利用掃描電子顯微鏡對(duì)其觸角進(jìn)行超微形態(tài)觀察。結(jié)果表明:蔥黃寡毛跳甲觸角為絲狀,由柄節(jié)、梗節(jié)和包括9亞節(jié)的鞭節(jié)組成。觸角上分布有6種感器,包括刺形感器Ⅰ、刺形感器Ⅱ、B?hm式鬃毛、錐形感器Ⅰ、錐形感器Ⅱ和毛形感器,刺形感器Ⅰ、表皮孔分布于整個(gè)觸角,刺形感器Ⅱ、錐形感器Ⅰ分布于鞭節(jié),B?hm式鬃毛、表皮附突分布于觸角基部,而錐形感器Ⅱ、毛形感器則主要分布于觸角端部。雌雄觸角除大小外,形態(tài)、感器類型和分布均無(wú)明顯差異。
鞘翅目;蔥黃寡毛跳甲;觸角;感器;掃描電鏡
昆蟲(chóng)間的信息交流、信號(hào)傳遞和感官感知主要表現(xiàn)為神經(jīng)系統(tǒng)的化學(xué)信息和化學(xué)感覺(jué),而化學(xué)通訊是昆蟲(chóng)感覺(jué)和傳遞信息的一個(gè)重要特性[1-2]。昆蟲(chóng)憑借其靈敏的感覺(jué)系統(tǒng)感知化學(xué)信號(hào)的傳遞,以滿足自身生存和繁衍的需要[3]。昆蟲(chóng)的觸角是感知化學(xué)信號(hào)的主要器官,是昆蟲(chóng)表皮特化形成的特殊結(jié)構(gòu),是昆蟲(chóng)感覺(jué)系統(tǒng)的重要組成部分,在寄主定位、識(shí)別、棲息、取食、躲避危險(xiǎn)、交配和繁殖以及信息素的接受等信息交換過(guò)程中承擔(dān)著重要角色[4]。不同類群的昆蟲(chóng)觸角形態(tài)和感器變化很大。研究觸角感器的類型、分布和數(shù)量,是探索昆蟲(chóng)一些重要行為的前提,有助于深入了解昆蟲(chóng)的觸角感器與行為反應(yīng)之間的關(guān)系,以及昆蟲(chóng)與自然界的化學(xué)通信等,并為昆蟲(chóng)系統(tǒng)發(fā)育和進(jìn)化研究提供重要依據(jù)[5]。
蔥黃寡毛跳甲LuperomorphasuturalisChen屬葉甲科Chrysomelinae,是韭菜上的重要害蟲(chóng)。成蟲(chóng)取食葉片,幼蟲(chóng)喜食韭菜須根,導(dǎo)致?tīng)I(yíng)養(yǎng)成分和水分的散失,造成韭菜大面積黃化甚至死亡[6],嚴(yán)重時(shí)可減產(chǎn)30%~50%,已成為韭菜地下害蟲(chóng)的優(yōu)勢(shì)種[7]。該蟲(chóng)發(fā)生環(huán)境隱蔽,危害時(shí)期長(zhǎng),生產(chǎn)上主要進(jìn)行化學(xué)防治,但由于大量使用殺蟲(chóng)劑導(dǎo)致該蟲(chóng)產(chǎn)生極強(qiáng)的抗藥性和過(guò)多的農(nóng)藥殘留,嚴(yán)重影響韭菜的產(chǎn)量和品質(zhì)甚至威脅人類健康[8]。
觸角感器調(diào)控昆蟲(chóng)的行為與周圍環(huán)境的化學(xué)、物理等因子密切相關(guān)。研究昆蟲(chóng)感器,有助于深入了解昆蟲(chóng)的化學(xué)感受系統(tǒng),探究其類型、數(shù)量、分布和功能,可進(jìn)一步了解昆蟲(chóng)觸角感器與行為反應(yīng)之間的關(guān)系。目前,有關(guān)對(duì)蔥黃寡毛跳甲的研究報(bào)道主要集中于生物學(xué)和生態(tài)學(xué)[6-8],還未見(jiàn)有關(guān)其觸角的報(bào)道。本文對(duì)蔥黃寡毛跳甲成蟲(chóng)觸角的形態(tài)以及感器的種類和分布進(jìn)行研究,以期為明確蔥黃寡毛跳甲嗅覺(jué)識(shí)別機(jī)制提供理論基礎(chǔ),并為設(shè)計(jì)新的高效引誘劑或趨避劑、開(kāi)展蔥黃寡毛跳甲防治提供新思路和新手段。
1 材料與方法
1.1 供試?yán)ハx(chóng)
蔥黃寡毛跳甲成蟲(chóng)于2014年8月采自陜西省渭南市華陰縣韭菜地,直接置于φ=70%酒精中保存,帶回后保存于4 ℃冰箱中,備用。
1.2 樣品制備
取雌、雄成蟲(chóng)各數(shù)頭,用φ=2.5%戊二醛固定液4 ℃固定過(guò)夜,再用磷酸緩沖液(PBS,0.1 mol/L, pH 7.2)洗滌3次,每次5 min,并在超聲波清洗儀中洗1 min,然后用φ=30%、φ=50%、φ=70%、φ=90%的乙醇和無(wú)水乙醇進(jìn)行逐級(jí)脫水,每次15 min,并分別用φ=25%、φ=50%、φ=75%、φ=100% 叔丁醇逐級(jí)置換,每次30 min。處理后的樣品放入CO2臨界干燥儀中進(jìn)行干燥。在顯微鏡下將干燥的觸角按不同面用導(dǎo)電膠粘于樣品臺(tái)上噴金,置于掃描電鏡(Hitachi S-3400N)下進(jìn)行觀察并拍照。
2 結(jié)果與分析
2.1 成蟲(chóng)觸角一般特征
蔥黃寡毛跳甲成蟲(chóng)觸角著生于兩個(gè)復(fù)眼之間,雌、雄觸角結(jié)構(gòu)無(wú)明顯差異,均為絲狀,由柄節(jié)(Scape)、梗節(jié)(Pedicel)和鞭節(jié)(Flagellomeres)組成,全長(zhǎng)約為3.24 mm(圖1-A)。柄節(jié)基部膨大、包被于膜質(zhì)的觸角窩內(nèi),粗短,近似鐘形,長(zhǎng)約100.12 μm,基部表面感器種類和數(shù)量較少,具褶皺及眾多魚(yú)鱗狀突起(圖1-B, 2-A~2-C)。梗節(jié)連接在柄節(jié)端部凹陷的窩內(nèi),呈圓柱狀,長(zhǎng)約115.61 μm,與柄節(jié)長(zhǎng)度接近,整個(gè)表面也布滿類似的魚(yú)鱗狀突起,感器種類少,但數(shù)量多于梗節(jié)(圖1-B,3-A,3-B)。鞭節(jié)細(xì)長(zhǎng)、約1.80 μm,遠(yuǎn)長(zhǎng)于柄節(jié)和梗節(jié)之和,鞭節(jié)分為9個(gè)亞節(jié)(圖1-A)。鞭末節(jié)端部1/3呈圓錐狀,其余各節(jié)均呈近圓柱狀,梗節(jié)最短,鞭節(jié)最長(zhǎng)(圖1-A)。鞭節(jié)中,第一鞭節(jié)最短而鞭末節(jié)最長(zhǎng),鞭末節(jié)約為第一鞭節(jié)的3倍,其余各鞭小節(jié)長(zhǎng)度差異不大(表1)。
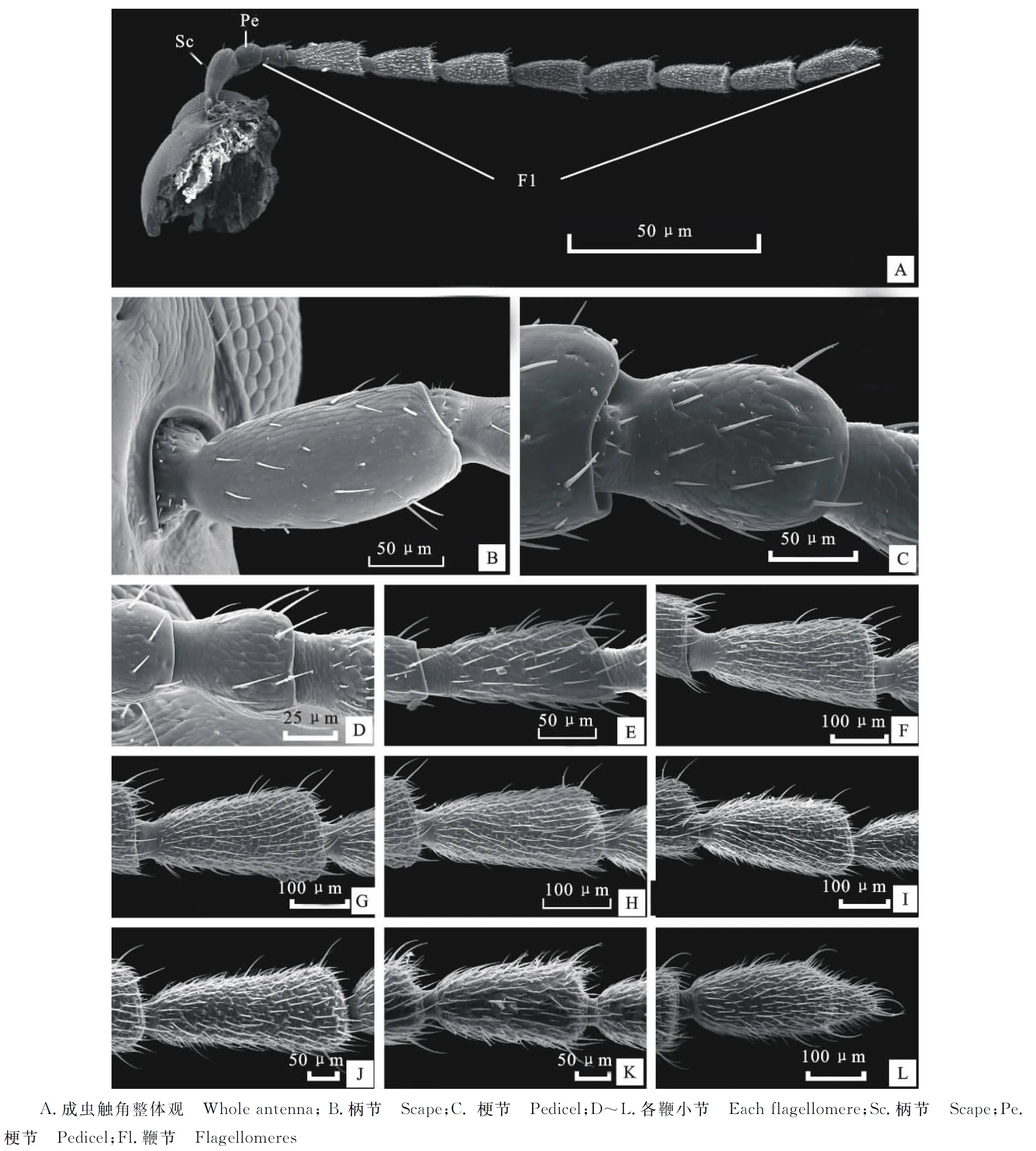
圖1 蔥黃寡毛跳甲成蟲(chóng)觸角及感器的掃描電鏡觀察
2.2 感器的種類、形態(tài)和分布
蔥黃寡毛跳甲成蟲(chóng)觸角表面分布有大量感器,依據(jù)分布位置與外部形態(tài)可將其分為6種,分別為刺形感器Ⅰ(Sensilla chaeticaⅠ)、刺形感器Ⅱ(Sensilla chaeticaⅡ)、錐形感器Ⅰ(Sensilla basiconicaⅠ)、錐形感器Ⅱ(Sensilla basiconicaⅡ)、毛形感器(Sensilla trichodea)和B?hm式鬃毛(B?hm bristles)。除此之外,蔥黃寡毛跳甲觸角著生有大量表皮孔(cuticular pores)和魚(yú)鱗狀感覺(jué)附肢(Appendages of cuticular plates),表皮孔在觸角各節(jié)表面均有發(fā)生,感覺(jué)附肢僅分布于基部的觸角窩內(nèi)。
2.2.1 刺形感器Ⅰ(Sensilla chaetica Ⅰ, ScⅠ) 數(shù)量最多,分布最廣,整個(gè)觸角均有分布。ScⅠ在柄節(jié)、梗節(jié)和第一鞭節(jié)分布較少,其中柄節(jié)內(nèi)側(cè)僅發(fā)現(xiàn)少量ScⅠ,梗節(jié)與第一鞭節(jié)ScⅠ均勻分布于整節(jié)(圖2-A~2-D,圖3-A,圖4-A)。ScⅠ在其余觸角各節(jié)呈均勻而密集地分布,數(shù)量最多(圖4-B,4-C)。ScⅠ著生于表皮特化的基節(jié)窩內(nèi),表面有縱紋,ScⅠ與觸角表面幾乎呈平行分布,直立,端部尖銳不彎曲,表面未發(fā)現(xiàn)感覺(jué)孔(圖4-E)。ScⅠ長(zhǎng)約為20~46 μm,由于各節(jié)長(zhǎng)度的差異,ScⅠ在長(zhǎng)度上也表現(xiàn)出與各節(jié)長(zhǎng)度變化一致的規(guī)律,即梗節(jié)和第1鞭小節(jié)較短,其余各節(jié)長(zhǎng)度差異不大。基節(jié)窩直徑約為(6.44±0.41) μm。
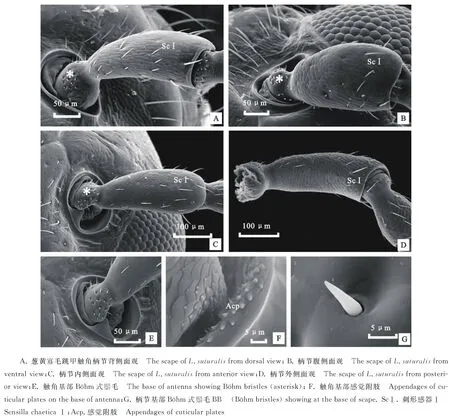
圖2 蔥黃寡毛跳甲成蟲(chóng)觸角柄節(jié)及感器的掃描電鏡觀察
2.2.2 刺形感器Ⅱ(Sensilla chaetica Ⅱ, ScⅡ) 主要呈螺紋狀分布于除第1鞭節(jié)之外各鞭節(jié)端部、末節(jié)端部圓錐狀和圓柱狀連接處以及觸角末端。ScⅡ與ScⅠ外部形態(tài)相似,但明顯長(zhǎng)于ScⅠ且端部向內(nèi)彎曲呈約30°夾角。ScⅡ著生于表皮凹陷的基節(jié)窩內(nèi),與觸角表面呈30~40°的夾角,端部鈍圓,表面無(wú)孔,有縱紋且數(shù)量明顯多于ScⅠ表面縱紋數(shù)(圖4-D,4-G)。ScⅡ長(zhǎng)約為(60.54±9.27) μm,基節(jié)窩直徑約為(6.30±0.55) μm。
2.2.3 B?hm 氏鬃毛(B?hm bristles,Bb) 主要分布于柄節(jié)基部與頭部的連接處以及梗節(jié)基部與柄節(jié)的連接處。Bb在柄節(jié)基部呈圓環(huán)狀分布,4面均有分布,梗節(jié)內(nèi)側(cè)沒(méi)有分布,其余3面均有分布(圖3-A,3-B,3-D)。柄節(jié)Bb數(shù)量相對(duì)較多。Bb短錐狀,表面光滑無(wú)縱紋并且無(wú)孔,垂直著生于觸角表面凹陷不明顯的基節(jié)窩內(nèi)(圖2-E,2-F)。Bb長(zhǎng)為(9.62±2.45) μm,基節(jié)窩直徑約為(2.57±0.28) μm。
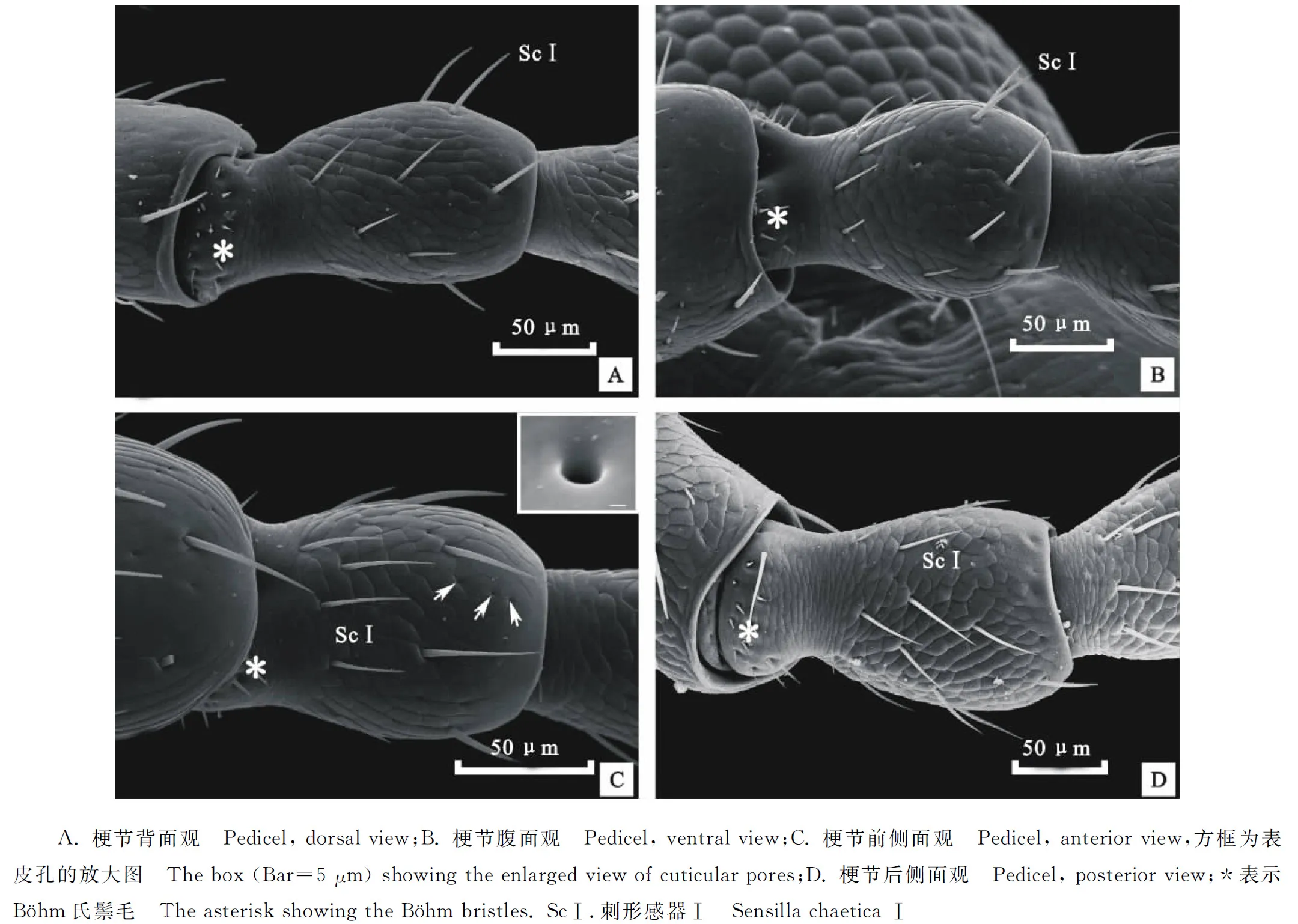
圖3 蔥黃寡毛跳甲成蟲(chóng)觸角梗節(jié)及感器的掃描電鏡觀察
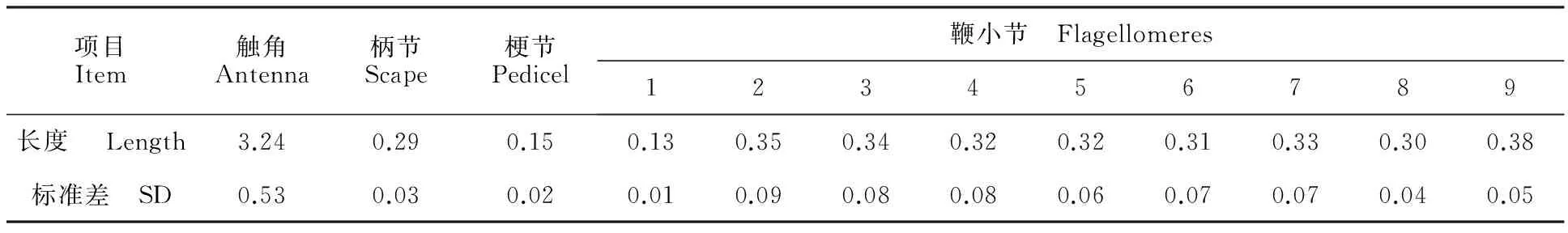
表1 觸角各節(jié)(亞節(jié))長(zhǎng)度
2.2.4 錐形感器(Sensilla basiconic Ⅰ,Sb Ⅰ) 均勻分布于除第3節(jié)之外的各鞭節(jié),柄節(jié)和梗節(jié)未發(fā)現(xiàn)此類感器。SbⅠ與ScⅠ間隔分布,與觸角表面平行,各鞭節(jié)SbⅠ數(shù)量明顯多于ScⅠ(圖4-B,4-C,4-E)。SbⅠ表面光滑無(wú)縱紋,端部鈍圓,未發(fā)現(xiàn)任何孔狀結(jié)構(gòu)。與刺形感器相比,SbⅠ基部的基節(jié)窩凹陷不明顯(圖4-F)。SbⅠ長(zhǎng)約為(10.30±1.52) μm,基節(jié)窩直徑約為(5.04±0.18) μm。
2.2.5 錐形感器Ⅱ(Sensilla basiconic Ⅱ,SbⅡ) 數(shù)量較少,僅分布于觸角端部,其他各節(jié)未發(fā)現(xiàn),其外部形態(tài)與SbⅠ相似,但明顯短于SbⅠ,且基部的基節(jié)窩有明顯差異,SbⅡ基節(jié)窩是由表皮凸起形成的,而SbⅠ基節(jié)窩略微凹陷。SbⅡ錐狀,短小,表面光滑無(wú)孔,直立,端部鈍圓不彎曲(圖4-I),長(zhǎng)約為(2.85±0.14) μm,基節(jié)窩直徑約為(1.24±0.12) μm。
2.2.6 毛形感器(Sensilla trichodea,St) 分布于觸角末節(jié)端部1/3處,其他各節(jié)未發(fā)現(xiàn)該感器分布。St較SbⅠ稍長(zhǎng),著生于表皮凸起形成的基節(jié)窩內(nèi),向頂端漸尖細(xì),端部直立或彎曲,表面光滑無(wú)孔,無(wú)縱紋(圖4-H)。St長(zhǎng)約為(31.87±5.41) μm,基節(jié)窩直徑約為(2.77±0.23) μm。
2.2.7 表皮孔(Cuticular pores,Cp) 觸角表面各感器之間分布大量由表皮凹陷形成的表皮孔,其表面光滑,個(gè)體小,不易發(fā)現(xiàn)(圖2-C)。孔徑大小約為(0.67±0.01) μm。
2.2.8 表皮附突(Appendages of cuticular plates,Acp) 蔥黃寡毛跳甲觸角基部外側(cè)圍角片內(nèi)以及觸角窩內(nèi)連接頭部的關(guān)節(jié)表面分布著大量的魚(yú)鱗狀表皮附突,基部相連、呈螺紋狀排列(圖2-F)。單獨(dú)的表皮附突長(zhǎng)約為(1.88±0.49) μm。
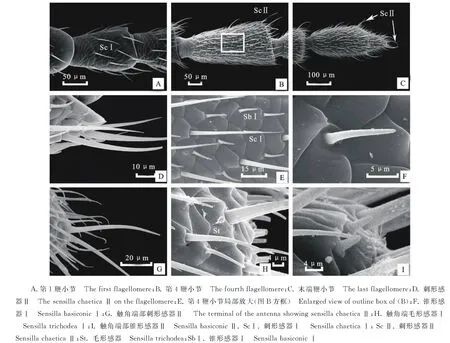
圖4 蔥黃寡毛跳甲成蟲(chóng)觸角鞭節(jié)及感器
3 討 論
蔥黃寡毛跳甲觸角為典型的絲狀,由柄節(jié)、梗節(jié)和鞭節(jié)組成,其表面著生有6種感器,包括2種刺形感器、2種錐形感器、毛形感器和B?hm氏鬃毛,與已報(bào)道大多數(shù)鞘翅目昆蟲(chóng)觸角感器種類特別相似[9-12]。但蔥黃寡毛跳甲觸角上的感器種類較少,未發(fā)現(xiàn)腔錐形感器和鐘形感器。簡(jiǎn)單而獨(dú)特的棲息環(huán)境和較少的天敵可能是造成感器種類的單一和退化的主要原因[4]。
刺形感器Ⅰ密集而均勻地分布于整個(gè)觸角表面,在多種鞘翅目昆蟲(chóng)中均有報(bào)道[11,13-14]。油菜蘭跳甲(Psylliodeschrysocephala)觸角的此類感器被認(rèn)為是毛形感器,但其與Nebriabrevicollis中所描述的刺形感器有著類似的內(nèi)部結(jié)構(gòu),僅有1個(gè)神經(jīng)元,終止于1個(gè)管狀結(jié)構(gòu)[15]。沙蒿金葉甲(Chrysolinaaeruginosa)觸角表面的此類感器被認(rèn)為具有保護(hù)其他感器的作用[11]。Daly和Ryan認(rèn)為此類感器為機(jī)械感受器[15]。
與蔥黃寡毛跳甲刺形感器Ⅱ分布和形態(tài)相似的感器在鞘翅目其他昆蟲(chóng)中也有描述[9,12,15-17]。N.brevicollis觸角上的此種感器內(nèi)部有5個(gè)神經(jīng)元,其中1個(gè)神經(jīng)元細(xì)胞是以管狀結(jié)構(gòu)結(jié)尾,其余則無(wú)分支結(jié)構(gòu)。基于其內(nèi)部結(jié)構(gòu),Daly和Ryan認(rèn)為此類感器兼有機(jī)械和化學(xué)感受器的功能[15]。P.chrysocephala觸角中ScⅡ具單孔,被認(rèn)為具有機(jī)械感受器和味覺(jué)感受器的功能[12]。Isidoro通過(guò)電生理學(xué)實(shí)驗(yàn)發(fā)現(xiàn)ScⅡ具有感受寄主植物氣體的功能[17]。
本研究中的B?hm式鬃毛與其他鞘翅目昆蟲(chóng)中相關(guān)感器非常相似[9-10,12-13],主要分布于觸角柄節(jié)、梗節(jié)基部。與Bembidionlampros相比,蔥黃寡毛跳甲的Bb基部基節(jié)窩無(wú)明顯凹陷,但其外部形態(tài)與分布位置均相似[10]。Bartlet等將P.chrysocephala觸角上的此類感器稱為毛板感受器,但其外部結(jié)構(gòu)以及分布的位置均與鞘翅目其他昆蟲(chóng)的B?hm式鬃毛相同[12]。組織學(xué)研究發(fā)現(xiàn)此類型感器由1個(gè)神經(jīng)元細(xì)胞控制,結(jié)合電生理學(xué)以及獨(dú)特的分布位置說(shuō)明此類感器是一類機(jī)械感受器(本體感受器),對(duì)昆蟲(chóng)感受觸角位置和運(yùn)動(dòng)以及感受方向和力量有著重要的作用[9]。
蔥黃寡毛跳甲觸角鞭節(jié)上分布大量的錐形感器Ⅰ,與步甲科的Bembidionproperans,B.lampros,沙蒿金葉甲,P.chrysocephala等昆蟲(chóng)觸角上的錐形感器形態(tài)和分布相似[9-12]。B.lampros的錐形感器依據(jù)其外部形態(tài)分為2個(gè)亞型[10],而蔥黃寡毛跳甲觸角的此類感器無(wú)論是在形態(tài)還有分布均無(wú)明顯差異,故未做亞型的劃分。Merivee等根據(jù)錐形感器外部形態(tài)、分布和數(shù)量推測(cè)其起著嗅覺(jué)感受器的作用[10]。另有研究發(fā)現(xiàn)此類感器表面有許多孔以及內(nèi)部有分枝狀的樹(shù)突,故確認(rèn)此類感器為嗅覺(jué)感受器[15]。
錐形感器Ⅱ著生的位置和外部形態(tài),分別與苜蓿葉象甲(Hyperapostica),B.lampros和Platynusdorsalis中描述的錐形感器相似[9,18-19]。雖然錐形感器通常被認(rèn)為屬于化學(xué)感受器,但該錐形感器的功能尚不明確,還需進(jìn)一步研究。
毛形感器略長(zhǎng)于錐形感器,基節(jié)窩隆起明顯,端部略有彎曲,在其他昆蟲(chóng)也有分布。研究發(fā)現(xiàn),在玉米螟、暗色叩頭甲和Melanoyusvillosus雄蟲(chóng)觸角的該類感器具有感受性信息素的功能[13,20],而B(niǎo).lampros和P.dorsalis雌雄觸角的此類感器被認(rèn)為具有聚集信息素的作用[10,19]。P.chrysocephala觸角表面的毛形感器多孔,能感受氣味,是一種嗅覺(jué)感受器[12]。本研究的毛形感器表面無(wú)孔,雌雄均有發(fā)生,推測(cè)其是一類感受信息素的感受器。
表皮感覺(jué)孔在直翅目、膜翅目等均有發(fā)生[21-23]。蔥黃寡毛跳甲觸角的表皮孔外形上與綠豆象觸角中所描述的腔型感器(sensilla cavity)[24]和B.properans[9]觸角中的腔錐形感器比較相似。綠豆象觸角的此類感器被認(rèn)為具有感受化學(xué)、濕度和溫度的功能。鱗翅目昆蟲(chóng)觸角上的表皮孔通過(guò)降解信息素和寄主揮發(fā)物以保護(hù)觸角的化學(xué)感受器[25-27]。而有關(guān)葉甲科觸角的研究中則認(rèn)為表皮孔有著產(chǎn)生信息素的作用[28-29]。表皮孔具體功能尚不清楚,還需進(jìn)一步研究。
本研究基于前人的研究對(duì)蔥黃寡毛跳甲觸角的感器功能進(jìn)行推測(cè),其特定的功能還不明確。為更加準(zhǔn)確地確定各個(gè)感器的功能,還需要較深入地觀察。
Reference:
[1]GREENFIELD M D.Signalers and Receivers:Mechanisms and Evolution of Arthropod Communication[M].Oxford,UK:Oxford University Press,2002.
[2]REINHARD J.Insect chemical communication [J].ChemoSense,2004,6(4):1-6.
[3]戴建青,韓詩(shī)疇,杜家緯.植物揮發(fā)性信息化學(xué)物質(zhì)在昆蟲(chóng)寄主選擇行為中的作用[J].環(huán)境昆蟲(chóng)學(xué)報(bào),2010,32 (3):407-414.
DAI J Q,HAN SH CH,DU J W.Progress in studies on behavioural effect of semiochemicals of host plant to insects [J].JournalofEnvironmentalEntomology,2010,32 (3):407-414 (in Chinese with English abstract).
[4]那 杰,于維熙,李玉萍,等.昆蟲(chóng)觸角感器的種類及其生理生態(tài)學(xué)意義[J].沈陽(yáng)師范大學(xué)學(xué)報(bào)(自然科學(xué)版),2008,26(2):213-216.
NA J,YU W X,LI Y P,etal.Types and physiological ecology significance of insect antennal sensilla [J].JournalofShenyangNormalUniversity(NaturalScience),2008,26(2):213-216 (in Chinese with English abstract).
[5]SUKONTASON K,KABKAEW L,SUKONTASON,etal.Antennal sensilla of some forensically important flies in families calliphoridae,sarcophagidae and muscidae [J].Micron,2004,35(8):671-679.
[6]張全力,李雙悅,陳 哲,等.蔥黃寡毛跳甲在辛集市嚴(yán)重發(fā)生[J].植保技術(shù)與推廣,2000(20):1.
ZHANG Q L,LI SH Y,CHEN ZH,etal.LuperomorphasuturalisChen occurs seriously in Xiji [J].PlantProtectionTechnologyandExtension,2000(20):1 (in Chinese).
[7]YANG X,JIAN H,LIU Z,etal.Evaluation of entomopathogenic nematodes for control of the beetle,LuperomorphasuturalisChen (Col.,Chrysomelidae) [J].JournalofAppliedEntomology,2003,127(7):377-382.
[8]黨志紅,高占林,賈海民,等.韭菜害蟲(chóng)蔥黃寡毛跳甲生物學(xué)特性簡(jiǎn)報(bào)[M]//成卓敏.農(nóng)業(yè)生物災(zāi)害預(yù)防與控制研究.北京:中國(guó)農(nóng)業(yè)科學(xué)技術(shù)出版社,2005:967-969.
DANG ZH H,GAO ZH L,JIA H M,etal.A Brief Account of the Biological Characteristics ofLuperomorphasuturalisChen.,a Pest of Leek [M]//CHENG ZH M,Agricultural Biohazard Prevent and Control Research.Beijing:China Agricultural Science and Technology Press,2005:967-969 (in Chinese).
[9]MERIVEE E,PLOOMI A,RAHI M,etal.Antennal sensilla of the ground beetleBembidionproperansSteph.(Coleoptera,Carabidae) [J].Micron,2002,33(5):429-440.
[10]MERIVEE E,PLOOMI A,RAHI M,etal.Antennal sensilla of the ground beetleBembidionlamprosHbst (Coleoptera,Carabidae) [J].ActaZoologica,2000,81(4):339-350.
[11]ZHANG L,REN L L,LUO Y Q,etal.Scanning electron microscopy analysis of the cephalic sensilla ofChrysolinaaeruginosaFald.(Coleoptera,Chrysomelidae) [J].MicroscopyResearchandTechnique,2013,76(4):423-431.
[12]BARTLET E,ROMANI R,WILLIAMS I H,etal.Functional anatomy of sensory structures on the antennae ofPsylliodeschrysocephalaL.(Coleoptera:Chrysomelidae) [J].InternationalJournalofInsectMorphologyandEmbryology,1999,28(99):291-300.
[13]MERIVEE E,RAHI M,LUIK A.Antennal sensilla of the click beetle,Melanotusvillosus(Geoffroy) (Coleoptera:Elateridae) [J].InternationalJournalofInsectMorphology&Embryology,1999,28(98):41-51.
[14]OKADA K,MORI M,SHIMAZAKI K,etal.Morphological studies on the antennal sensilla of the cigarette beetle,Lasiodermaserricorne(F.) (Coleoptera:Anobiidae) [J].AppliedEntomology&Zoology,1992,27(2):269-276.
[15]DALY P J,RYAN M F.Ultrastructure of antennal sensilla ofNebriabrevicollis(Fab.) (Coleoptera :Carabidae) [J].InternationalJournalofInsectMorphology&Embryology,1979,8(3):169-181.
[16]KIM J L,YAMASAKI T.Sensilla ofCarabus(Isiocarabus)fiducariussaishutoicusCsiki (Coleoptera:Carabidae) [J].InternationalJournalofInsectMorphology&Embryology,1996,25(1/2):153-172.
[17]ISIDORO N,BARTLET E,ZIESMANN J,etal.Antennal contact chemosensilla inPsylliodeschrysocephalaresponding to cruciferous allelochemicals [J].PhysiologicalEntomology,2002,23(2):131-138.
[18]BLAND R G.Antennal sensilla of the adult alfalfa weevil,Hyperapostica(Gyllenhal) (Coleoptera :Curculionidae) [J].InternationalJournalofInsectMorphology&Embryology,1981,10(3):265-274.
[19]MERIVEE E,PLOOMI A,LUIK A,etal.Antennal sensilla of the ground beetlePlatynusdorsalis(Pontoppidan,1763) (Coleoptera,Carabidae) [J].MicroscopeResearchTechnology,2002,55(5):339-349.
[20]HALLBERG E,HANSSON B S,STEINBRECHT R A.Morphological characteristics of antennal sensilla in the European cornborerOstrinianubilalis(Lepidoptera:Pyralidae) [J].Tissue&Cell,1994,26(4):489-502.
[21]BLAND R G.Antennal sensilla of Acrididae (Orthoptera) in relation to subfamily and food preference [J].AnnalsoftheEntomologicalSocietyofAmerica,1989,82(3):368-384.
[22]WCISLO W T.Sensilla numbers and antennal morphology of parasitic and non-parasitic bees (Hymenoptera :Apoidea) [J].InternationalJournalofInsectMorphology&Embryology,1995,24(1):63-81.
[23]AMORNSAK W,CRIBB B,GORDH G.External morphology of antennal sensilla ofTrichogrammaaustralicumgirault (Hymenoptera:Trichogrammatidae) [J].InternationalJournalofInsectMorphology&Embryology,1998,27(2):67-82.
[24]HU F,ZHANG G H,WANG J J.Scanning electron microscopy studies of antennal sensilla of bruchid beetles,Callosobruchuschinensis(L.) and Callosobruchus maculatus (F.) (Coleoptera:Bruchidae) [J].Micron,2009,40(3):320-326.
[25]TAYLOR T R,FERKOVICH S M,ESSEN F V.Increased pheromone catabolism by antennal esterases after adult eclosion of the cabbage looper moth[J].Cellular&MolecularLifeSciencesCmls,1981,37(7):729-731.
[26]VOGT R G,RIDDIFORD L M.Pheromone binding and inactivation by moth antennae [J].Nature,1981,293(5828):161-163.
[27]DICKENS J C,VISSER J H,PERS J N C V D.Detection and deactivation of pheromone and plant odor components by the beet armyworm,Spodopteraexigua(Hübner) (Lepidoptera:Noctuidae) [J].JournalofInsectPhysiology,1993,39(6):503-516.
[28]BARTLET E,ISIDORO N,WILLIAMS I H.Antennal glands inPsylliodeschrysocephala,and their possible role in reproductive behavior [J].PhysiologicalEntomology,1994,19(4):241-250.
[29]MEDVEDEV L N,PAVLOV S I.Mating behavior of the Chrysomelidae (Coleoptera) [J].EntomologicalReview,1988,67:100-109.
College of Plant Protection, Northwest A&F University, Yangling Shaanxi 712100, China)
(責(zé)任編輯:郭柏壽 Responsible editor:GUO Baishou)
Morphological Characters of Antenna and Types of Antennal Sensilla of Leaf BeetleLuperomorphasuturalisChen (Coleoptera:Chrysomelidae)
ZHANG Qi, CHEN Fan, ZHANG Chunni and DAI Wu
(Key Laboratory of Plant Protection Resources and Integrated Pest Management, Ministry of Education,
The fine morphology and distribution of sensilla onLuperomorphasuturalis, which is a significant agricultural pest, were investigated using scanning electron microscopy. Antennae are made up of the scape, pedicel, and nine segment of flagellomeres. Antennae are filiform in shape. Four different sensillar types were distinguished, including two types of sensilla chaetica, two types of sensilla basiconica, sensilla trichodea and b?hm bristles. Cuticular pores and appendages of cuticular plates were also found on the antenna. Among them, Sensilla chaeticaⅠ and cuticular pores were distributed on the whole antenna, sensilla chaeticaⅡ and sensilla basiconicaⅠ were found on the flagellomeres, B?hm bristles and appendages of cuticular plates were observed on the base of antenna, sensilla basiconicaⅡ and sensilla trichodea were located on the terminal of antenna.
Coleoptera;Luperomorphasuturalis; Antenna, Sensilla, Scanning electron microscopy
ZHANG Qi, male, master student. Research area: the relationship of insects and plants.E-mail:qizhang0523@163.com
DAI Wu, male,professor,Ph.D,doctoral supervisor.Research area:entomology.E-mail:daiwu@nwsuaf.edu.cn
2016-02-19
2016-03-23
公益性行業(yè)(農(nóng)業(yè))科研專項(xiàng)(201303027);中國(guó)科學(xué)院“西部之光”人才計(jì)劃(2012DF06);中央高校基本科研業(yè)務(wù)費(fèi)專項(xiàng)(YQ2013010)。
張 琪,男,在讀碩士,從事昆蟲(chóng)與植物關(guān)系研究。E-mail: qizhang0523@163.com
戴 武,男,教授,博士,博士生導(dǎo)師,主要從事昆蟲(chóng)學(xué)研究。E-mail:daiwu@nwsuaf.edu.cn
日期:2016-10-20
Q964
A
1004-1389(2016)10-1567-08
網(wǎng)絡(luò)出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20161020.1658.044.html
Received 2016-02-19 Returned 2016-03-23
Foundation item The Special Fund for Agro-scientific Research in the Public Interest(No.201303027);the West Light Foundation of the Chinese Academy of Sciences (No.2012DF06);the Chinese Universities Scientific Fund (No.YQ2013010).
