miR-506對(duì)結(jié)腸癌細(xì)胞增殖的影響及其作用機(jī)制
2016-12-05 06:04:56祖彩華張國(guó)梁劉濤
山東醫(yī)藥 2016年40期
祖彩華,張國(guó)梁,劉濤
(1 天津醫(yī)科大學(xué)一中心臨床學(xué)院,天津300192;2 天津市第一中心醫(yī)院)
?
miR-506對(duì)結(jié)腸癌細(xì)胞增殖的影響及其作用機(jī)制
祖彩華1,張國(guó)梁2,劉濤2
(1 天津醫(yī)科大學(xué)一中心臨床學(xué)院,天津300192;2 天津市第一中心醫(yī)院)
目的 探討miR-506對(duì)結(jié)腸癌細(xì)胞增殖的影響及其作用機(jī)制。方法 將結(jié)腸癌細(xì)胞SW620隨機(jī)分為四組,分別轉(zhuǎn)染pcDNA3空載體(pcDNA3組)、miR-506過表達(dá)質(zhì)粒pcDNA3/pri-506(miR-506組)、pSIH1空載體(pSIH1組)及miR-506抑制表達(dá)質(zhì)粒pSIH1/TuD-506(TuD-506組)。采用qRT-PCR法檢測(cè)各組miR-506 mRNA的相對(duì)表達(dá)量,CCK-8法檢測(cè)各組的細(xì)胞活性,集落形成試驗(yàn)檢測(cè)各組的集落形成能力。采用生物信息學(xué)軟件預(yù)測(cè)miR-506的潛在靶基因,熒光報(bào)告載體試驗(yàn)驗(yàn)證結(jié)腸癌細(xì)胞SW620中miR-506對(duì)靶基因mRNA的調(diào)控作用。采用qRT-PCR法和Western blotting法檢測(cè)miR-506對(duì)靶基因mRNA及其蛋白表達(dá)的影響。結(jié)果 與pcDNA3組比較,miR-506組miR-506 mRNA的相對(duì)表達(dá)量明顯升高,而細(xì)胞活性、集落形成能力明顯減弱(P均<0.01);TuD-506組與pSIH1組miR-506 mRNA的相對(duì)表達(dá)量、細(xì)胞活性、集落形成能力比較差異均無統(tǒng)計(jì)學(xué)意義(P均>0.05)。生物信息學(xué)軟件預(yù)測(cè)發(fā)現(xiàn),層黏連蛋白γ1(LAMC1)是miR-506的潛在靶基因,經(jīng)熒光報(bào)告載體試驗(yàn)驗(yàn)證,miR-506能夠直接作用于LAMC1 mRNA并負(fù)性調(diào)控其表達(dá)。miR-506組LAMC1 mRNA及其蛋白的相對(duì)表達(dá)量均低于pcDNA3組(P均<0.05),TuD-506組與pSIH1組LAMC1 mRNA及其蛋白的相對(duì)表達(dá)量比較差異均無統(tǒng)計(jì)學(xué)意義(P均>0.05)。結(jié)論 miR-506可抑制結(jié)腸癌的細(xì)胞活性、集落形成能力,LAMC1可能是miR-506的直接靶基因;miR-506可能通過抑制LAMC1的表達(dá)而抑制結(jié)腸癌細(xì)胞增殖。
結(jié)腸癌;微小RNA-506;細(xì)胞增殖;層黏連蛋白γ1
微小RNA(miRNA)是一類內(nèi)源性、高度保守的單鏈非編碼小RNA,長(zhǎng)約22個(gè)核苷酸,其主要作用是與下游靶mRNA的3′-非編碼區(qū)(3′UTR)配對(duì);當(dāng)二者不完全配對(duì)時(shí),可抑制靶mRNA的翻譯或促進(jìn)靶mRNA的降解,導(dǎo)致相應(yīng)的蛋白表達(dá)下降[1,2]。有研究表明,miRNAs在細(xì)胞增殖、分化和凋亡中具有重要作用,某些miRNA還具有類似癌基因或抑癌基因作用[3,4]。層黏連蛋白γ1(LAMC1)是層黏連蛋白家族中的一員,在基膜中顯著表達(dá),可促進(jìn)腫瘤轉(zhuǎn)移。有研究發(fā)現(xiàn),LAMC1可能是miR-506的潛在靶點(diǎn),有可能成為治療腫瘤的新方向。2015年7月~2016年3月,本研究觀察了miR-506對(duì)結(jié)腸癌細(xì)胞增殖的影響,并探尋其下游靶基因,旨在為結(jié)腸癌治療提供新靶點(diǎn)。
1 材料與方法
1.1 材料 結(jié)腸癌細(xì)胞系SW620,由中國(guó)科學(xué)院上海生命科學(xué)研究院細(xì)胞庫(kù)提供。miR-506過表達(dá)質(zhì)粒pcDNA3/pri-506和miR-506抑制表達(dá)質(zhì)粒pSIH1/TuD-506,由本實(shí)驗(yàn)室前期構(gòu)建。miR-506、U6 snRNA、LAMC1及β-actin特異性PCR引物,由北京天潤(rùn)奧科生物科技有限公司合成。DMEM/F12(1∶1)培養(yǎng)基、FBS,美國(guó)Gibco BRL;Opti-MEM Ⅰ無血清培養(yǎng)液、脂質(zhì)體轉(zhuǎn)染試劑Lipofectamine 2000 Reagent,美國(guó)Life Technologies Corporation;兔源人LAMC1和GAPDH抗體,美國(guó)Santa Cruz;HRP標(biāo)記的羊抗兔IgG Fc抗體,美國(guó)Abcam;mirVana RNA提取試劑盒,美國(guó)Ambion;PrimeScript Ⅱ 1st Strand cDNA Synthesis Kit逆轉(zhuǎn)錄試劑盒和SYBR Premix Ex Taq Ⅱ熒光定量PCR試劑盒,日本TaKaRa。LightCycle 96熒光定量PCR儀,瑞士Roche;Alpha Innotech FluorChem FC2凝膠成像分析系統(tǒng),美國(guó)Alpha Innotech;水平/垂直電泳槽和電泳設(shè)備,北京六一儀器廠;EnSpireTMMultilabel Reader酶標(biāo)儀,美國(guó)PerkinElmer。
1.2 細(xì)胞培養(yǎng)及轉(zhuǎn)染 取結(jié)腸癌細(xì)胞SW620置于含10% FBS的高糖DMEM培養(yǎng)液中,37 ℃、5% CO2培養(yǎng)箱中培養(yǎng),每2~3天按1∶3傳代一次。取對(duì)數(shù)生長(zhǎng)期細(xì)胞,接種于細(xì)胞培養(yǎng)板中,待細(xì)胞貼壁超過80%時(shí)進(jìn)行轉(zhuǎn)染。質(zhì)粒轉(zhuǎn)染按照Lipofectamine 2000試劑說明書進(jìn)行。分別轉(zhuǎn)染pcDNA3空載體(pcDNA3組)、miR-506過表達(dá)質(zhì)粒pcDNA3/pri-506(miR-506組)、pSIH1空載體(pSIH1組)及miR-506抑制表達(dá)質(zhì)粒pSIH1/TuD-506(TuD-506組)。
1.3 相關(guān)指標(biāo)觀察
1.3.1 細(xì)胞活性 采用CCK-8法。調(diào)整各組轉(zhuǎn)染后的細(xì)胞密度為1×105個(gè)/mL,接種于96孔板中,每孔100 μL。培養(yǎng)48 h,每孔加入CCK-8試劑10 μL,繼續(xù)培養(yǎng)2 h。酶標(biāo)儀檢測(cè)各組450 nm波長(zhǎng)處的吸光度(A450)值。試驗(yàn)重復(fù)3次,取平均值。
1.3.2 細(xì)胞集落形成能力 各組轉(zhuǎn)染24 h,消化計(jì)數(shù),并接種于12孔板,每孔100個(gè)細(xì)胞。每3天更換一次培養(yǎng)液,觀察至細(xì)胞出現(xiàn)單個(gè)集落,停止培養(yǎng)。用2%結(jié)晶紫染液染色10 min,1×PBS溶液脫色后,顯微鏡下拍照觀察。以大于50個(gè)細(xì)胞作為集落形成,計(jì)數(shù)集落個(gè)數(shù)。試驗(yàn)重復(fù)3次,取平均值。
1.3.3 miR-506的靶基因預(yù)測(cè)及驗(yàn)證 應(yīng)用生物信息學(xué)軟件TargetScanHuman (http://www.targetscan.org/vert_71/),輸入miRNA的名稱hsa-miR-506,預(yù)測(cè)miR-506的靶基因。根據(jù)生物信息學(xué)軟件預(yù)測(cè)的結(jié)果,將LAMC1 3′UTR上的miR-506結(jié)合位點(diǎn)克隆插入GFP熒光報(bào)告基因編碼區(qū)下游,同時(shí)將miR-506“種子序列”位點(diǎn)突變插入GFP編碼區(qū)下游,分別構(gòu)建帶有野生型(pcDNA3/EGFP-LAMC1-wtUTR)和突變型(pcDNA3/EGFP-LAMC1-mutUTR)結(jié)合位點(diǎn)的熒光報(bào)告載體。轉(zhuǎn)染前1天,將結(jié)腸癌細(xì)胞SW620接種于24孔板中。轉(zhuǎn)染當(dāng)天,將載有熒光報(bào)告載體的質(zhì)粒和載有miR-506過表達(dá)或miR-506抑制表達(dá)質(zhì)粒共同轉(zhuǎn)染細(xì)胞,每組設(shè)3個(gè)復(fù)孔。轉(zhuǎn)染24 h,將細(xì)胞原培養(yǎng)液吸棄,用1×PBS浸洗1次,加入放免分析裂解緩沖液100 μL/孔,冰浴搖床搖動(dòng)60 min。用移液器反復(fù)吹打各孔細(xì)胞,將細(xì)胞裂解液移至1.5 mL離心管中,4 ℃ 12 000 r/min離心10 min,吸取上清至另一管,得到細(xì)胞蛋白樣品。取各孔蛋白樣品100 μL置于酶標(biāo)板,用酶標(biāo)儀檢測(cè)蛋白樣品中綠色熒光蛋白(GFP)的表達(dá)強(qiáng)度(激發(fā)光為488 nm,發(fā)射光為507 nm),以對(duì)照組GFP的表達(dá)量為1,計(jì)算各組校正后GFP的相對(duì)表達(dá)量。試驗(yàn)重復(fù)3次,取平均值。
1.3.4 miR-506、LAMC1 mRNA表達(dá) 采用qRT-PCR技術(shù)。采用RNA提取試劑盒提取結(jié)腸癌細(xì)胞的RNA。以此為模板,逆轉(zhuǎn)錄生成cDNA。按照qRT-PCR SYBR Green法進(jìn)行PCR擴(kuò)增。引物序列:miR-506-Fwd:5′-TGCGGTAAGGCACCCTTCTGA-GTAG-3′;U6-Fwd:5′-TGCGGGTGCTCGCTCGGCAG-C-3′;miR-506/U6-Rvs:5′-CCAGTGCAGGGTCCGAG-GT-3′;LAMC1-Fwd:5′-CATGTATCTGAATACCTCC-3′;LAMC1-Rvs:5′-CATAGCTTGTAACCTG-3′;β-ac-tion-Fwd:5′-CGTGACATTAAGGAGAAGCTG-3′;β-action-Rvs:5′-CTAGAAGCATTTGCGGTGGAC-3′;LA-MC1-wtUTR-top:GATCCACTGTGGACCTTTTATATG-TGCCTTCACTTTAGCTGTTTGCCTTAATCTG;LAMC-1-wtUTR-bottom:AATTCAGATTAAGGCAAACAGCT-AAAGTGAAGGCACATATAAAAGGTCCACAGTG; LA-MC1-mutUTR-top:GATCCACTGTGGACCTTTTATAT-CAGGGATCACTTTAGCTGTTCACGAAAATCTG; LA-MC1-mutUTR-bottom:AATTCAGATTTTCGTGAACA-GCTAAAGTGATCCCTGATATAAAAGGTCCACAGTG。反應(yīng)條件:95 ℃預(yù)變性3 min,95 ℃ 5 s、56 ℃ 30 s、72 ℃ 30 s,擴(kuò)增40個(gè)循環(huán)后進(jìn)行熔解曲線分析,以2-ΔΔCt法進(jìn)行計(jì)算。試驗(yàn)重復(fù)3次,取平均值。
1.3.5 LAMC1蛋白表達(dá) 采用Western blotting法。收集各組轉(zhuǎn)染后細(xì)胞,用RIPA裂解液裂解,提取細(xì)胞蛋白。蛋白樣品經(jīng)SDS-PAGE分離并轉(zhuǎn)移至硝酸纖維素膜。以兔抗人LAMC1和GAPDH為一抗,以HRP標(biāo)記的羊抗兔IgG Fc抗體為二抗,進(jìn)行Western blotting檢測(cè)。硝酸纖維素膜經(jīng)ECL顯影液孵育后,應(yīng)用LabWorksTM凝膠成像系統(tǒng)采集信號(hào)。圖像經(jīng)AlPhaView分析軟件進(jìn)行灰度值測(cè)定。通過計(jì)算每組樣品灰度值(目的蛋白條帶亮度值-背景值)與對(duì)應(yīng)蛋白灰度值(內(nèi)參蛋白GAPDH條帶亮度值-背景值)的比值,得到校正后目的蛋白的相對(duì)表達(dá)量。以對(duì)照組目的蛋白的表達(dá)量為1,計(jì)算各組校正后目的蛋白的相對(duì)表達(dá)量。試驗(yàn)重復(fù)3次,取平均值。
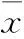
2 結(jié)果
2.1 各組miR-506 mRNA相對(duì)表達(dá)量及細(xì)胞活性、集落形成能力比較 見表1。
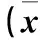
表1 各組miR-506 mRNA相對(duì)表達(dá)量及細(xì)胞活性、集落形成能力比較±s)
注:僅同載體間進(jìn)行比較;與pcDNA3組比較,*P<0.01。
2.2 miR-506的靶基因預(yù)測(cè) 生物信息學(xué)軟件預(yù)測(cè)發(fā)現(xiàn),LAMC1可能為miR-506的潛在靶基因。
2.3 各組pcDNA3/EGFP-LAMC1-wtUTR、pcDNA3/EGFP-LAMC1-mutUTR相對(duì)表達(dá)量比較 見表2。

表2 各組pcDNA3/EGFP-LAMC1-wtUTR、pcDNA3/EGFP-LAMC1-mutUTR相對(duì)表達(dá)量比較
注:僅同載體間進(jìn)行比較;與pcDNA3組比較,*P<0.01。
2.4 各組LAMC1 mRNA及其蛋白表達(dá)比較 見表3。
表3 各組LAMC1 mRNA及其蛋白相對(duì)表達(dá)量比較
注:僅同載體間進(jìn)行比較;與pcDNA3組比較,*P<0.05。
3 討論
目前結(jié)腸癌發(fā)生、發(fā)展的分子機(jī)制尚不完全清楚。近年研究發(fā)現(xiàn),miRNAs表達(dá)失調(diào)對(duì)結(jié)腸癌細(xì)胞的侵襲和遷移能力具有一定影響[5~10]。由此推測(cè),miRNAs可能參與了結(jié)腸癌的發(fā)生、發(fā)展。
miR-506是miRNAs的一員,其在不同腫瘤細(xì)胞中表達(dá)不同。Sun等[11]研究發(fā)現(xiàn),與正常乳腺上皮細(xì)胞比較,miR-506在乳腺癌細(xì)胞系(T47D、MDA-MB-231、SK-BR-3、HCC1937)中顯著低表達(dá)。進(jìn)一步研究發(fā)現(xiàn),miR-506可通過抑制IQGAP1/ERK信號(hào)傳導(dǎo)通路來抑制乳腺癌細(xì)胞的增殖、侵襲。Arora等[12]研究證實(shí),miR-506能抑制乳腺癌細(xì)胞系MDA-MB-231的上皮間質(zhì)轉(zhuǎn)化及癌細(xì)胞的黏附、侵襲和遷移。Yin等[13]研究發(fā)現(xiàn),miR-506通過抑制NF-κB p65信號(hào)傳導(dǎo)通路,增加活性氧的產(chǎn)生,激活p53信號(hào)通路,繼而導(dǎo)致肺癌細(xì)胞的凋亡、抑制肺癌細(xì)胞的活性,但在正常肺組織細(xì)胞中卻無此作用。說明miR-506可選擇性殺傷肺癌細(xì)胞,有可能成為治療肺癌的潛在靶點(diǎn)。Wang等[14]研究發(fā)現(xiàn),miR-506在原發(fā)性肝癌組織中低表達(dá),在肝癌細(xì)胞系HepG2和H7402中,miR-506通過靶定yes相關(guān)蛋白(YAP)mRNA的3′UTR來抑制癌細(xì)胞的增殖。有研究報(bào)道,80%宮頸癌組織miR-506低表達(dá),且與Ki-67的表達(dá)呈負(fù)相關(guān)。表明miR-506可抑制宮頸癌細(xì)胞的增殖。但也有研究報(bào)道,在黑色素瘤組織中miR-506表達(dá)升高,并可促進(jìn)黑色素瘤的發(fā)生、發(fā)展[15]。以上研究表明,miR-506可能具有抑癌和促癌雙重作用。
本研究發(fā)現(xiàn),與pcDNA3組比較,miR-506組細(xì)胞活性、集落形成能力明顯減弱;而pSIH1組與TuD-506組細(xì)胞活性、集落形成能力比較差異均無統(tǒng)計(jì)學(xué)意義。因此,尚不能證實(shí)抑制miR-506對(duì)結(jié)腸癌細(xì)胞的增殖有影響。其原因可能是結(jié)腸癌細(xì)胞SW620中miR-506低表達(dá),抑制其表達(dá)對(duì)其產(chǎn)生的影響微乎其微。其具體原因尚需進(jìn)一步研究。
層黏連蛋白是由α、β和γ鏈組成的三聚體化合物,是細(xì)胞外基質(zhì)的重要調(diào)節(jié)因子,對(duì)細(xì)胞的分化、遷移和黏附有一定影響[16]。層黏連蛋白表達(dá)升高可促進(jìn)癌細(xì)胞的遷移和侵襲[17,18]。LAMC1是層黏連蛋白家族中的一員,Ke等[19]報(bào)道,在黑色素瘤細(xì)胞中LAMC1表達(dá)升高,其表達(dá)變化與腫瘤短期內(nèi)復(fù)發(fā)及無瘤生存期縮短有關(guān)。Nishikawa等[20]研究證實(shí),LAMC1在前列腺癌中高表達(dá)。進(jìn)一步研究發(fā)現(xiàn),miR-29s通過直接靶定LAMC1的信號(hào)傳導(dǎo)通路而抑制癌細(xì)胞的活性。生物信息學(xué)軟件預(yù)測(cè)發(fā)現(xiàn),LAMC1可能是miR-506的下游靶點(diǎn)。本研究結(jié)果顯示,對(duì)于帶有野生型結(jié)合位點(diǎn)LAMC1 3′UTR的熒光報(bào)告載體,過表達(dá)miR-506能夠抑制GFP的表達(dá)強(qiáng)度;而對(duì)于帶有突變型結(jié)合位點(diǎn)LAMC1 3′UTR的熒光報(bào)告載體,miR-506不能影響報(bào)告基因的GFP表達(dá)強(qiáng)度。證實(shí)miR-506能夠直接作用于LAMC1 3′UTR的特定序列,并對(duì)結(jié)合位點(diǎn)上游熒光報(bào)告基因的表達(dá)起負(fù)性調(diào)控作用。本研究還發(fā)現(xiàn),miR-506組LAMC1 mRNA及其蛋白的相對(duì)表達(dá)量降低,進(jìn)一步證實(shí)LAMC1是miR-506的下游靶點(diǎn)。
綜上所述,miR-506能抑制結(jié)腸癌的細(xì)胞活性和集落形成能力,其作用的靶基因可能是LAMC1。但miR-506是否還調(diào)控其他靶基因來影響結(jié)腸癌細(xì)胞的生物活性尚需進(jìn)一步研究。
[1] Bartel DP. MicroRNAs: target recognition and regulatory functions[J]. Cell, 2009,136(2):215-233.
[2] Inui M, Martello G, Piccolo S. MicroRNA control of signal transduction[J]. Nat Rev Mol Cell Biol, 2010,11(4):252-263.
[3] Zhang B, Pan X, Cobb GP, et al. MicroRNAs as oncogenes and tumor suppressors[J]. Dev Biol, 2007,302(1):1-12.
[4] Nelson KM, Weiss GJ. MicroRNAs and cancer: past, present, and potential future[J]. Mol Cancer Ther, 2009,7(12):3655-3660.
[5] Zhang L, Dong Y, Zhu N, et al. microRNA-139-5p exerts tumor suppressor function by targeting NOTCH1 in colorectal cancer[J]. Mol Cancer, 2014(13):491-500.
[6] Liu X, Zhang Z, Sun L, et al. MicroRNA-499-5p promotes cellular invasion and tumor metastasis in colorectal cancer by targeting FOXO4 and PDCD4[J]. Carcinogenesis, 2011,32(12):1798-1805.
[7] Wu W, Yang J, Feng X, et al. MicroRNA-32 (miR-32) regulates phosphatase and tensin homologue (PTEN) expression and promotes growth, migration, and invasion in colorectal carcinoma cells[J]. Mol Cancer, 2013(12):30.
[8] Zhang Y, Wang Z, Chen M, et al. MicroRNA-143 targets MACC1 to inhibit cell invasion and migration in colorectal cancer[J]. Mol Cancer, 2012(11):23.
[9] Panza A, Votino C, Gentile A, et al. Peroxisome proliferator-activated receptor γ-mediated induction of microRNA-145 opposes tumor phenotype in colorectal cancer[J]. Biochim Biophys Acta, 2014,1843(6):1225-1236.
[10] Zhang GJ, Xiao HX, Tian HP, et al. Upregulation of microRNA-155 promotes the migration and invasion of colorectal cancer cells through the regulation of claudin-1 expression[J]. Int J Mol Med, 2013,31(6):1375-1380.
[11] Sun G, Liu Y, Wang K, et al. miR-506 regulates breast cancer cell metastasis by targeting IQGAP1[J]. Int J Oncol, 2015,47(5):1963-1970.
[12] Arora H, Qureshi R, Park WY. miR-506 regulates epithelial mesenchymal transition in breast cancer cell lines[J]. PLoS One, 2013,8(5):e64273.
[13] Yin M, Ren X, Zhang X, et al. Selective killing of lung cancer cells by miRNA-506 molecule through inhibiting NF-κB p65 to evoke reactive oxygen species generation and p53 activation[J]. Oncogene, 2015,34(6):691-703.
[14] Wang Y, Cui M, Sun BD, et al. MiR-506 suppresses proliferation of hepatoma cells through targeting YAP mRNA 3′UTR[J]. Acta Pharmacol Sin, 2014,35(9):1207-1214.
[15] Streicher KL, Zhu W, Lehmann KP, et al. A novel oncogenic role for the miRNA-506-514 cluster in initiating melanocyte transformation and promoting melanoma growth[J]. Oncogene, 2012,31(12):1558-1570.
[16] Simon-Assmann P. The laminin family: founding members of the basement membrane[J]. Cell Adh Migr, 2013,7(1):44-47.
[17] Tzu J, Marinkovich MP. Bridging structure with function: structural, regulatory, and developmental role of laminins[J]. Int J Biochem Cell Biol, 2008,40(2):199-214.
[18] Yamaguchi M, Ebihara N, Shima N, et al. Adhesion, migration, and proliferation of cultured human corneal endothelial cells by laminin-5[J]. Invest Ophthalmol Vis Sci, 2011,52(2):679-684.
[19] Ke HL, Ke RH, Li B, et al. Association between laminin γ1 expression and meningioma grade, recurrence, and progression-free survival[J]. Acta Neurochir (Wien), 2013,155(1):165-171.
[20] Nishikawa R, Goto Y, Kojima S, et al. Tumor-suppressive microRNA-29s inhibit cancer cell migration and invasion via targeting LAMC1 in prostate cancer[J]. Int J Oncol, 2014,45(1):401-410.
Effects of miR-506 on proliferation of colorectal carcinoma cells and its mechanism
ZUCaihua1,ZHANGGuoliang,LIUTao
(1FirstCentralClinicalCollegeofTianjinMedicalUniversity,Tianjin300192,China)
Objective To investigate the effects of microRNA-506 (miR-506) on proliferation of colorectal carcinoma cells and to identify the target gene for miR-506. Methods The colorectal carcinoma SW620 cells were respectively divided into pcDNA3 group, miR-506 group, pSIH1 group and TuD-506 group according to different tranfection plasmids. The real-time fluorescent quantitative PCR was used to detect the expression of miR-506. Cell viability and colony-forming ability was measured by CCK8 and colony formation assays, respectively. Furthermore, bioinformatics method was applied to predict potential target genes of miR-506.Green fluorescent protein (GFP) reporter assays were used to verify the direct regulation of miR-506 on target mRNA in SW620 cells. The LAMC1 mRNA and protein levels were detected by quantitative reverse transcription polymerase chain reaction (qRT-PCR) and Western blotting, respectively. Results Compared with the pcDNA3 group, the relative expression of miR-506 in the miR-506 group was significantly higher, while the activity and colony formation ability of the miR-506 group was significantly decreased (allP<0.01). No statistically significant difference was found in the relative expression of miR-506, cell viability and colony forming ability between the TuD-506 group and pSIH1 group (allP>0.05). LAMC1 was predicted to be a potential target gene of miR-506, and GFP reporter assays verified miR-506 directly targeted LAMC1 mRNA and negatively regulated its expression. The expression of miR-506 in the miR-506 group was significantly lower than that of the pcDNA3 group (allP<0.05). No statistically significant difference was found in the LAMC1 mRNA and protein expression between the TuD-506 group and pSIH1group (allP>0.05). Conclusion miR-506 plays a tumor suppressor role in SW620 cells by inhibiting cell viability and colony formation, LAMC1 is a direct target gene for miR-506, and miR-506 inhibits proliferation of colorectal cancer cells by down-regulating the expression of LAMC1.
colorectal carcinoma; miR-506; cell proliferation; laminin γ1
國(guó)家自然科學(xué)基金資助項(xiàng)目(81402322);2013年度天津市企業(yè)博士后創(chuàng)新項(xiàng)目擇優(yōu)資助計(jì)劃。
祖彩華(1989-),女,碩士在讀,研究方向?yàn)橄到y(tǒng)疾病。E-mail: caihuazu@163.com
簡(jiǎn)介:劉濤(1981-),男,主管技師,博士,研究方向?yàn)榉蔷幋aRNA對(duì)腫瘤細(xì)胞的調(diào)控。E-mail: 13821217761@163.com
10.3969/j.issn.1002-266X.2016.40.002
R735.3
A
1002-266X(2016)40-0005-04
2016-03-25)
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
西南國(guó)防醫(yī)藥(2016年7期)2016-12-01 06:01:15
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年1期)2016-01-14 03:41:26
河南醫(yī)學(xué)研究(2014年3期)2014-02-27 14:51:48
沈陽(yáng)醫(yī)學(xué)院學(xué)報(bào)(2014年1期)2014-02-16 06:19:24

- 山東醫(yī)藥的其它文章
- 急性非ST段抬高型心肌梗死患者血清BNP水平與病情及預(yù)后的關(guān)系
- 鼻咽癌組織Ezrin、E-cadherin表達(dá)及其與腫瘤侵襲、轉(zhuǎn)移及患者預(yù)后的關(guān)系
- 212例頭頸部惡性腫瘤患者HTR3B基因多態(tài)性與化療后出現(xiàn)惡心嘔吐的關(guān)系
- 中文修訂版疾病認(rèn)知問卷在維持性血液透析患者中的信效度評(píng)價(jià)
- 先天性巨結(jié)腸病變腸段CAD、S-100蛋白表達(dá)及其臨床意義
- 特利加壓素輔助治療中重度肝硬化腹水的臨床效果及其對(duì)肝腎功能、門脈系統(tǒng)內(nèi)徑的影響