山西油松林分類學多樣性
2016-11-29 00:35:13秦曉娟趙小娜
生態學報 2016年20期
王 燁,秦 浩,董 剛,,秦曉娟,趙小娜,張 峰,,*
1 山西大學黃土高原研究所, 太原 030006 2 山西大學生命科學學院, 太原 030006
?
山西油松林分類學多樣性
王 燁1,秦 浩1,董 剛1,2,秦曉娟2,趙小娜1,張 峰1,2,*
1 山西大學黃土高原研究所, 太原 030006 2 山西大學生命科學學院, 太原 030006
基于物種分類系統關系的分類學多樣性不僅能反映群落內物種分類學之間的差異,而且可以間接反映生態系統或群落是否處于退化階段。與傳統的物種多樣性指數相比,分類學多樣性指數對不同的取樣方法和大小具有較強的穩健性。以山西油松林為研究對象,在山西省范圍內選取113個樣地進行植被調查,并結合歷史資料和相關文獻整理了植物名錄,野外共記錄植物357種,隸屬于3門4綱36目71科227屬。植物科內屬、種的組成差異較大,其中含屬、種最多的科是菊科,分別為31屬和50種,含種最多的屬為蒿屬(Artemisia11種)。選取平均分類學差異指數(Δ+)和分類學差異變異指數(Λ+)作為度量群落分類學多樣性的指標,同時分析了分類學多樣性指數與環境因子的關系,研究結果表明:113個樣地的Δ+值大都位于Δ+理論平均值上方(89.5%);Λ+值都位于Λ+理論平均值上方,這表明山西油松林物種組成的親緣關系較遠,分屬不同的分類單元;群落內物種分類單元的均勻度較差,即物種主要集中在幾個大的分類單元內,如菊科、薔薇科、豆科和禾本科等。對分類學多樣性指數影響最大的是群落結構因子(胸徑、喬木密度和群落物種數),其次是微地形因子(坡向和坡位)。土壤全氮和有機碳與Δ+呈極顯著正相關(P<0.01)。宏觀地形因子(緯度和海拔)與Δ+和Λ+相關性不顯著(P>0.05)。通過對山西油松林物種的Δ+和Λ+研究表明,山西油松林大多處于平穩的成長階段,物種組成相對穩定,存在一定的物種分類學差異,但主要集中幾個大的分類單元。
山西;油松林;平均分類學差異指數(Δ+);分類學差異變異指數(Λ+)
生物多樣性是評價物種數量、結構類型[1]、分布格局[2]、時空變化的有力工具[3]。傳統的生物多樣性指數大都基于物種個體數及相關信息,如Margalef豐富度指數、Pielou均勻度指數及Simpson多樣性指數等,這些指數對取樣的時間、地點、樣地大小與方法極其敏感[4],同時也不能直接反映種間的分類等級關系。
分類等級關系是群落物種多樣性的重要方面,當兩個群落物種豐富度相同時,分類關系較遠的群落比分類關系較近的有更大的多樣性[5]。為了研究群落分類學關系的變化,Clarke等提出了考慮物種間的分類關系的指數[6],主要包括分類多樣性指數Δ、分類差異指數Δ*、平均分類學差異指數Δ+和分類學差異變異指數Λ+[7]。平均分類學差異指數Δ+和分類學差異變異指數Λ+是基于物種間權重,根據分類等級的路徑長度,將群落分類學多樣性特征量化的方法[8-9]。其平均值不依賴于取樣性質和樣本大小,可用于大尺度的研究,避免取樣性質未知或不一致對取樣數據的影響[10],且易于計算[11]。Δ+反映群落種間親緣關系[12],Δ+值越高表示物種間親緣關系越遠,多樣性越大;Λ+可以反映群落物種分類關系的均勻程度[13],Λ+值越大表明物種分布均一性越差。分類學多樣性研究最初集中在動物區系和魚類群落[6,14]。如Leonard等[15]應用不同的分類指標評估英國海洋生物多樣性;Ellingsen等[16]研究了挪威大陸架底棲生物的分類學多樣性。近十余年來,在植被生態研究中分類學多樣性方法逐漸得到應用,如李景文等[17]對海南主要熱帶森林植物的分類學多樣性的研究;Da Silva等[18]對巴西中部稀樹草原的研究;秦曉娟等[3]檢驗了分類學多樣性對濕地植物群落的適用性;趙小娜等[19]對龐泉溝自然保護區植物群落分類學多樣性進行研究,并分析了分類學多樣性指數與環境因子的關系。
油松(Pinustabuliformis)是中國北方地區重要的用材和生態樹種,以油松為建群種形成的森林群落是華北山地優勢植被類型之一,山西是我國油松林分布的集中區之一,具有廣泛的代表性。本文在對山西油松群落進行野外調查的基礎上,運用分類學多樣性指數(Δ+和Λ+)對山西油松群落的分類學多樣性進行研究,探討山西油松林物種組成特征和分類學多樣性及與其有關的物種進化關系、物種共存和群落構建等問題,分析地形因子和群落因子與分類學多樣性指數的相關性,分析大尺度上影響物種分類學差異性的因子,為群落構建等問題提供一定的理論依據;同時,檢驗分類學多樣性分析方法在森林群落生態學中的應用效果,全面了解區域尺度上油松林的分類學多樣性,為山西油松林資源保護與管理提供科學依據。
1 研究區概況
油松林在山西北起恒山,至五臺山折向西,在岢嵐、興縣以南分布極為廣泛。以太行山的盂縣、平定、和順、左權、榆社、黎城、陵川,太岳山的沁源、霍州、介休、靈石、洪洞,呂梁山的交城、中陽、蒲縣、鄉寧,五臺山的繁峙、五臺、代縣,恒山,中條山的沁水、陽城、翼城[20-21],黑茶山的嵐縣、興縣、方山,管涔山的寧武、五寨、岢嵐等地分布較為集中。其中太岳山是我國油松林分布的代表性區域[22],而大同天然油松很少成林[23]。
野外調查的油松林(山西)分布范圍:110°45′—113°45′ E,34°48′—38°57′ N,海拔744—1867 m。研究區屬溫帶大陸性季風氣候,全年日照時數2808 h,無霜期120 d[24]。年平均氣溫為4—14 ℃,1月平均溫為-6.8 ℃,7月平均溫為23.6℃。年降水量為400—600 mm。
2 研究方法
2.1 野外調查
2012年6月—2014年9月,在山西共設置113個樣地進行植被調查(圖1),幾乎遍及山西省油松林分布區。樣地面積為20×30 m2,記錄喬木層樹種名、高度、冠幅、多度和郁閉度等;將樣地劃分為6個10×10 m2的樣方,選取對角線的2個樣方,記錄灌木層種名、多度、平均高和蓋度;在6個樣方中各選取1個1×1 m2的小樣方,記錄草本層種名、平均高和蓋度[25-26],采集每個樣方的表層土,密封保存,分析土壤屬性。記錄的環境信息包括經緯度、海拔、坡度、坡向、坡位和干擾情況等。
坡向采用等級制,即陽坡(1)、半陽坡(2)、半陰坡(3)和陰坡(4),數字越大,表示水分條件越好。坡位同樣采用等級制,即下坡位(1)、中下坡位(2)、中坡(3)、中上坡(4)和上坡位(5);干擾情況可分為無干擾(4)、輕微(3)、中度(2)和重度(1),數字越小,表示人為干擾相對越強。將調查的環境因子劃分為宏觀地形因子(經度、緯度和海拔)、微地形因子(坡度、坡向和坡位)、土壤屬性因子(全氮、全磷、pH和有機碳)和群落結構因子(胸徑、喬木密度和群落物種數),以便進行比較。
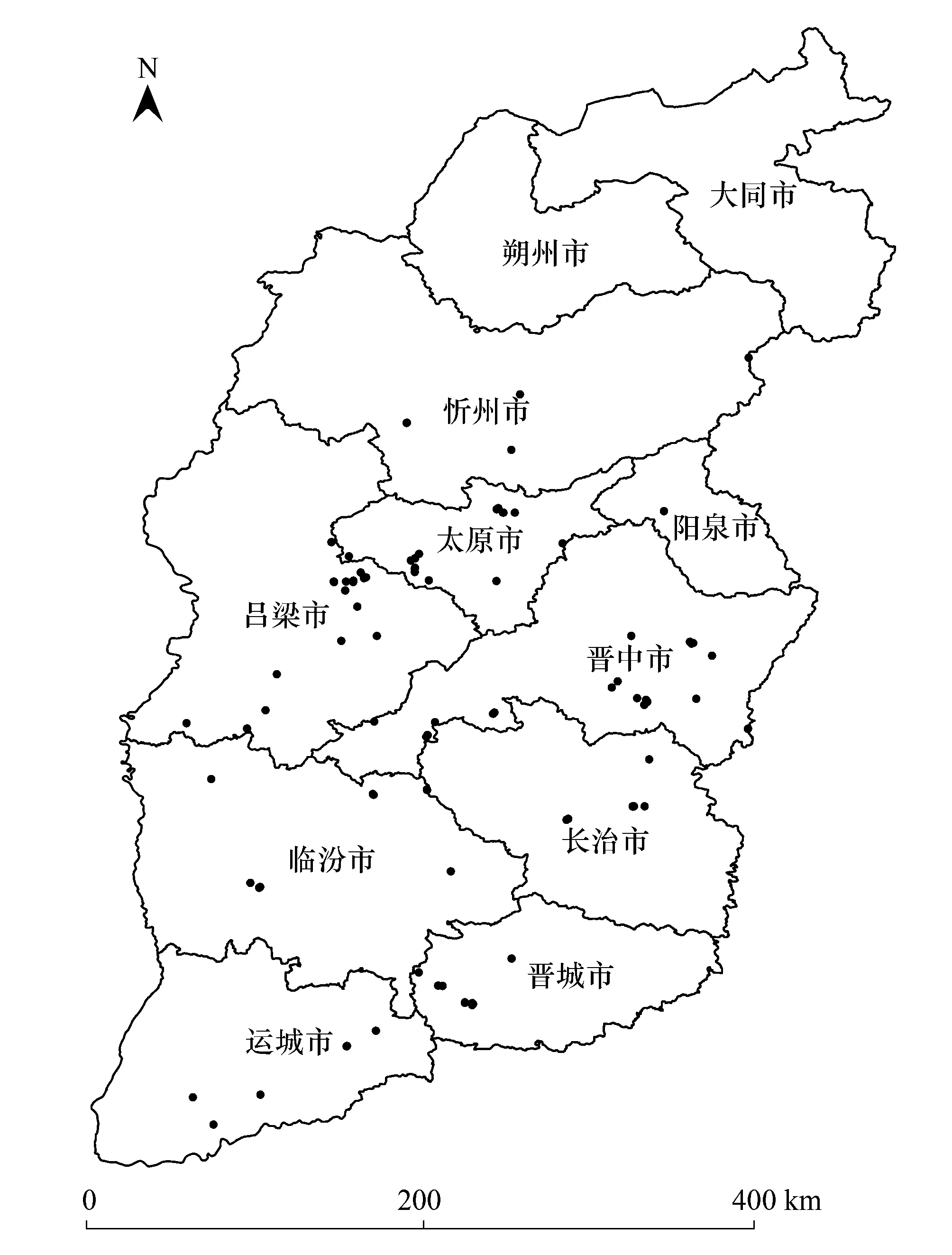
圖1 調查地點的基本信息 Fig.1 Dsitribution of stands of Pinus tabulaeformis in Shanxi
2.2 數據分析
綜合野外調查和歷史資料整理山西油松林物種總名錄。根據林奈分類系統,將物種分類等級確定為門、綱、目、科、屬、種6個等級。定義分類等級上最遠的2個種路徑長度為100(即最長路徑)[6],根據分類等級確定每一等級水平上的權重[27-28]:同屬的不同種ωij=16.67[3];同科不同屬的種ωij=33.33;同目不同科的種ωij=50.00;同綱不同目的種ωij=66.67;同門不同綱的種ωij=83.33;不同門的種ωij=100。
Clarke等[29]提出了計算分類學多樣性測度的方法,公式如下:
平均分類學差異指數
(1)
分類學差異變異指數
(2)
式中,ωij為第i和第j個物種在分類等級中的路徑長度,S為樣方中出現的種數。
對各樣地的多樣性指數值與理論平均值進行統計性檢驗,建立95%的置信漏斗曲線[5,30]。由PRIMER6軟件可求得Λ+與Δ+的值并完成置信漏斗的繪制。
用R軟件完成Λ+和Δ+與地形因子、土壤屬性和群落結構的Spearman秩相關分析。
3 結果與分析
3.1 群落結構與組成
山西北部油松林多為人工油松純林,林下灌木較少,草本植物優勢種為長芒草(Stipacapillata)等。中西部呂梁山、太岳山和太行山油松林成熟林較少,其中太岳山中幼齡林占優勢;關帝林區的油松多為中幼齡林,且多為純林,同時分布有少量的松、櫟類混交林;太行山區幼齡林約占2/3,其余大部分為中齡林,常與櫟類混交;灌木層主要有黃刺玫(Rosaxanthina)、美薔薇(Rosabella)、三裂繡線菊(Spiraeatrilobata)、土莊繡線菊(Spiraeapubescens)、虎榛子(Ostryopsisdavidiana)等;草本層以披針薹草(Carexlancifolia)為主。南部中條山幼齡林居多,中齡林約占l/3,常與白樺(Betulaplatyphylla)、遼東棟(Quercuswutaishanica)、元寶槭(Acertruncatum)、山楊(Populusdavidiana)等混交,灌木種類和數量較多,常見的有胡枝子(Lespedezabicolo)、黃刺玫(Rosaxanthina)、金銀忍冬(Loniceramaackii)、荊條(Vitexnegundvar.heterophylla)、西北栒子(Cotoneastermultiflorus)、甘肅山楂(Crataeguskansuensis)、土莊繡線菊、黃櫨(Cotinuscoggygriavar.coggygria)、暴馬丁香(Syringareticulata)等,草本層主要以羊胡子草(Carexrigescens)、華北米蒿(Artemisiagiraldii)等為主。
野外共記錄357種植物,隸屬于3門4綱36目71科227屬。3個門分別為被子植物門(包含2綱350種)、裸子植物門(包含1綱5種)和蕨類植物門(包含1綱2種)。被子植物中雙子葉植物綱(30目)占優勢地位。從目水平來看,管狀花目含科最多,有6科。含屬最多的為桔梗目(34屬),含種最多的為薔薇目(71種)。植物科內種、屬組成差異較大,其中菊科(31屬)、薔薇科(18屬)、豆科(15屬)禾本科(15屬)、唇形科(11屬)等含屬較多。菊科含種最多(50種),占總種數的14.01%,其次為豆科34種(9.52%)、薔薇科33種(9.24%)和禾本科21種(5.88%)。含種較多的屬為蒿屬(Artemisia11種)、堇菜屬(Viola9種)、委陵菜屬(Potentilla7種)、沙參屬(Adenophora7種)和薹草屬(Carex 5種)等。
由此可見,山西油松林群落大都屬于中幼齡林,林下的物種組成和分類單元相對比較豐富,但主要以幾個大的分類單元為主,如在科水平上集中于菊科、薔薇科、豆科和禾本科。
3.2 分類學多樣性指數
根據山西油松林的Δ+和Λ+理論平均值建立95%的置信漏斗曲線(圖2)。圖2中置信漏斗的上下置信限分別為相應種數在最大階元和最小階元的分類學多樣性理論值。圖2Δ+中粗線是Δ+的理論平均值(約為53),所有樣地Δ+理論平均值與種數的變化基本獨立。Δ+閾值在種數為4—7時明顯減小,之后隨著種數的增加,逐漸趨近于穩定,與Δ+理論平均值平行。
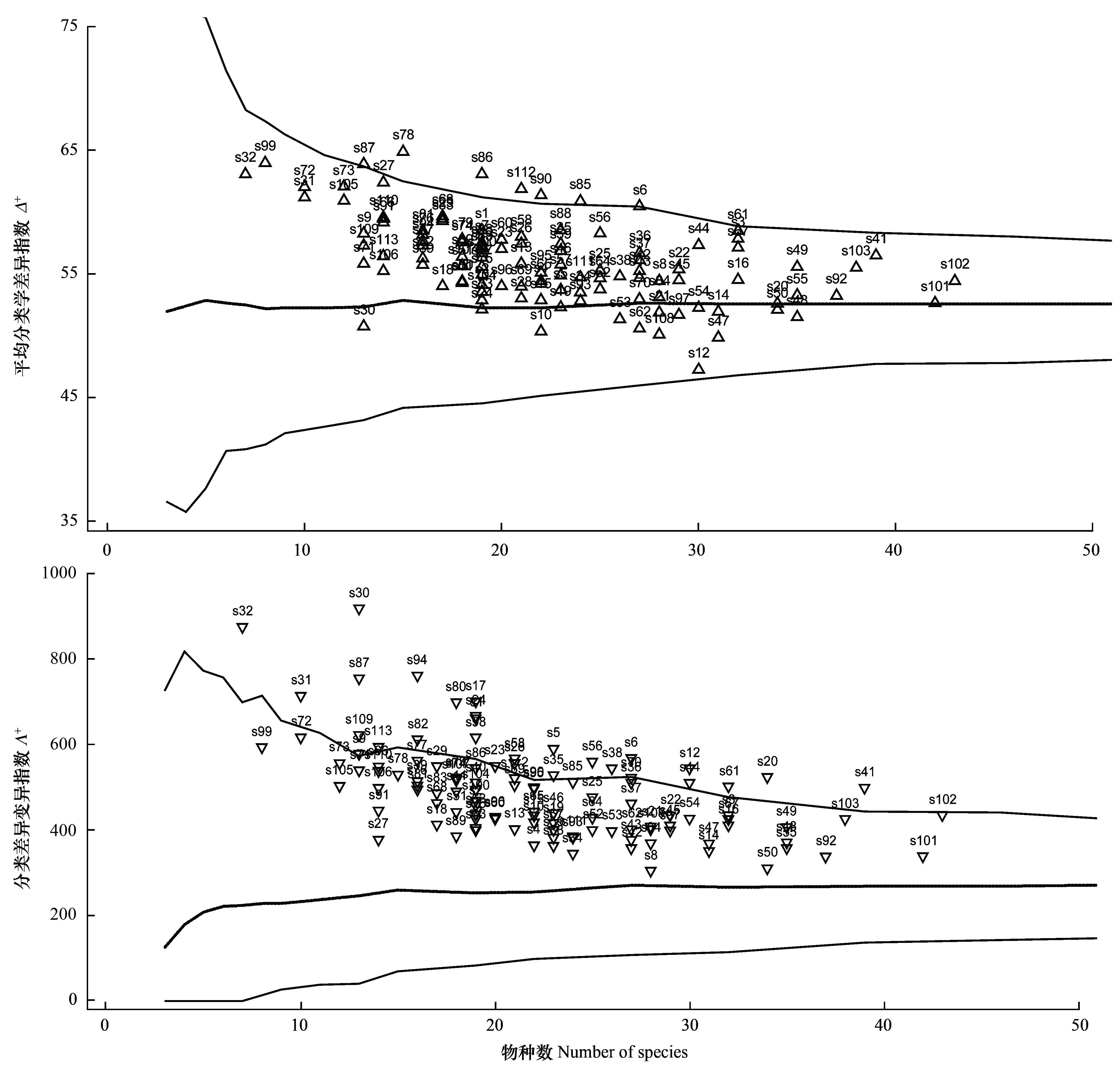
圖2 Δ+和Λ+ 95%的置信漏斗曲線及各調查點在曲線中的位置Fig.2 Δ+ and Λ+ for the plant communities in 113sites, plotted against the number of species on the 95% confidence funnel
Δ+值表示群落中所有種間路徑長度的理論平均值,置信漏斗范圍內樣地Δ+值越大,表示樣地內種間分類學多樣性越大,種間親緣關系越遠。Δ+值大都處于研究區Δ+的理論平均值之上,表明所調查的樣地內種間的分類學多樣性比較豐富。其中位于置信漏斗上包跡線上方的樣地占總樣地數的11.5%,包括樣地S6(永濟五老峰)、S78(晉源區天龍山)、S85(陽城莊溝)等,這些樣地的Δ+值顯著高于區域的Δ+理論平均值(P<0.05)。位于理論平均值下方的樣地數遠遠小于其上方樣地數,僅有樣地S10(運城鹽湖區)、S12(安澤草峪嶺)、S30(襄垣古韓)、S47(左權程家村)、S108(交城雙家寨)等10個;位于下包跡線之外的樣地并未出現。樣地S78平均分類差異指數Δ+值(64.86)高于其余樣地,即樣地S78種間親緣關系較其他樣地遠。樣地S12的Δ+值(47.23)最低,平均每科含3.33種,每屬含3種,表明其物種組成間的親緣關系較近。
圖2置信漏斗的上下置信限分別為Λ+值理論的最大曲線與最小曲線,Λ+值可表征物種分類單元分布的均一性水平。置信漏斗范圍內樣地Λ+值越小,表明油松群落間的分類地位關系程度越均勻。虛線是Λ+的理論平均值,反映區域內物種分類等級間的差異變異情況。由圖2可以看出,Λ+理論平均值也幾乎與種數變化無關,基本保持在280左右。只有當種數很少時(<5),Λ+的最大和最小曲線才會出現急劇變化;之后隨著種數的增加逐漸趨于穩定,且逐漸平行于Λ+理論平均值。
所有樣地的Λ+值均位于理論平均值上方,表明油松林種間的分類等級均勻性低于區域內的理論平均值。其中有25個樣地位于置信漏斗最大值曲線上方,分別為樣地S5(平陸東郭)、S6、S12、S17(永和芝河)、S20(鄉寧臺頭)等。這些樣地的Λ+值顯著高于區域內的理論平均值(P<0.05),表現出種間分類單元顯著的不均勻性。樣地S30分類差異變異指數Λ+(917.35)最高,表明其物種的等級分布最不均勻。樣地S30共有13種植物分屬2門3綱6目7科11屬,其中菊科含種最多(7種),其他皆為單屬單種的科。相反,樣地S8(垣曲毛家彎)分類差異變異指數Λ+(304.66)低于其余樣地,基本接近區域內的理論平均值,這說明S8物種等級分布相對均勻。樣地S8共有28種植物,隸屬2門3綱16目20科26屬,其中毛茛目、薔薇目、管狀花目和中央種子目各有兩個科,其他各目僅包含一個科。豆科、菊科、堇菜科含物種數最多,各有3種植物。
由Δ+和Λ+與種的分布可以看出,Δ+和Λ+隨著物種的增加基本呈現出下降的趨勢。樣地S102(寧武接官亭)種數最多,共43種,Δ+和Λ+值相對較低,分別為54.4和434.47。樣地S32種數最少,雖然僅有7種植物但它們分屬2門3綱5目5科6屬,平均分類差異指數(63.04)較高,分類差異變異指數(874.28)較高。
3.3 分類學多樣性指數與環境因子和群落結構的相關分析
由表1可知,宏觀地形因子與Δ+相關不顯著(P>0.05),與Λ+相關亦不顯著(P>0.05),其中緯度和海拔與Δ+和Λ+相關性大于經度,主要原因是山西緯度跨度大于經度和海拔梯度且變化明顯。微地形因子中,坡向與Δ+極顯著正相關(P<0.01),坡位與Δ+顯著正相關(P<0.05),與Λ+極顯著正相關(P<0.01)。
土壤屬性因子與Λ+相關不顯著(P>0.05);全氮和有機碳與Δ+呈極顯著正相關,pH和全磷與Δ+相關不顯著(P>0.05)。
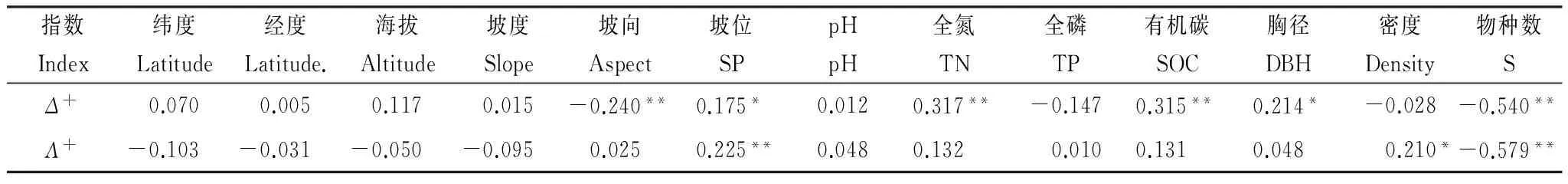
表1 Δ+、Λ+與環境因子和群落結構的Spearman秩相關關系
SP:坡位 Slope position;TN:全氮 Total nitrogen;TP:全磷 Total phosphorus;SOC:有機碳 Soil organic carbon;DBH:胸徑 Diameter at breast height;S:物種數 Number of species
群落結構因子中,物種數與Δ+呈極顯著負相關(P<0.01),與Λ+呈極顯著負相關(P<0.01);喬木密度與Δ+相關不顯著(P>0.05),與Λ+顯著正相關(P<0.05);胸徑與Δ+顯著正相關(P<0.05),與Λ+相關不顯著(P>0.05)。由此可以看出,影響油松林Δ+和Λ+分布的最直接的因子主要是微地形因子和群落結構因子。
4 討論
山西油松林的植物主要集中于菊科、豆科、禾本科、莎草科。多數樣地的Λ+值高于理論平均值,表明山西油松群落物種分布均一性較弱。Pienkowski等研究表明,Δ+值落入95%的置信漏斗最小值曲線以下時,表示該樣地的生境發生退化或受到嚴重污染[13,31]。山西油松林所有樣地Δ+值沒有位于置信漏斗最小值曲線下方的,表明山西油松林分布區域的生態環境整體情況良好,受到的各種干擾較少。這主要得益于1998年以來的退耕還林政策的實施和山西省生態文明建設各種管理措施的全面落實,包括停止對油松林的采伐、加強對盜伐油松林的處罰和監管等。五老峰和天龍山為旅游勝地,油松林處于中幼林階段,林隙間較為充足的光照有利于林下灌木和草本的生長,使得林下物種多樣性更豐富,群落分類學多樣性增高;同時,旅游對油松林有輕度干擾,加之一些植物繁殖體也可能隨游人進入松林,其中某些種可能在油松林定居并成功完成生活史,使得樣地S6和S78具有較高的分類學多樣性。樣地S85、S86、S87、S90、S112,屬于國有林場,油松林發育良好,群落結構完整,喬木層可分為兩個亞層,其中第一亞層為建群種——油松,第二亞層物種較為豐富,常見的有遼東櫟、山楊、白樺等,灌木層和草本層發育也較為完整,從而使得這些樣地具有較高的分類學多樣性。這一結果與其他研究者的結論一致[3],即結構復雜的群落具有更高的分類學多樣性指數。樣地S12位于安澤草峪嶺鄉道附近,墾荒、出行等人類日常活動嚴重干擾了林下和林緣植被的生長和種子的傳播;油松林處于幼林階段,且位于陽坡,水分條件較差,不利于林下植物的生長,分布的草本大多為菊科和禾本科等抗逆性強的種。因此,樣地S12的Δ+值最小。
相關性分析(表1)表明,盡管山西油松林分布的緯度和海拔變異明顯,但緯度和海拔并不是導致Δ+和Λ+分布的差異的決定性因子,亦即平均分類差異指數Δ+并沒有表現出明顯的緯度和海拔變化規律,與李永振等研究結果類似[11,32],這說明Δ+指數在區域尺度上反映群落的分類學多樣性具有較強的穩健性;同時,也表明山西是油松林分布相對適宜的區域,林下物種組成主要以菊科、禾本科等為主,它們具有較廣的生態幅和適應性。微地形因子中,坡向與Δ+極顯著正相關(P<0.01),即陰坡的油松林具有較高的分類學多樣性。坡向的差異會影響光照和水分的空間異質性[20]。在山西油松林分布區光照充足,限制植物生長和分布的主要是水分因子。在陽坡由于光照時間長、強度大,分布的物種往往以耐旱性較強的植物為主;由于土壤持水量較低,種間對水分的競爭強度相對較大,物種組成往往集中于某一科或某一屬。如樣地S30位于襄垣古韓鄉土橋村海拔1122 m的陽坡,充分的光照使得喜陽的油松長勢良好,喬木層郁閉度較大(0.85)不利于林下灌木和草本的生存;S30共有13種植物分屬2門3綱6目7科11屬,其中含種最多的科為菊科(7種),其他科皆為單屬單種,如展枝沙參(Adenophoradivaricata)、披針薹草、角盤蘭(Herminiummonorchis)、油松、茜草(Rubiacordifolia)、雞腿堇菜(Violaacuminata)。位于交城龐泉溝陰坡的樣地S90,由于水分條件好于陽坡,喬木層郁閉度較小(0.55),林下灌木層和草本層組成豐富多樣,共有22種植物分屬2門3綱13目16科21屬,包括側柏(Platycladusorientalis)、灰栒子(Cotoneasteracutifolius)、美薔薇、野青茅(Deyeuxiaarundinacea)、筋骨草(Ajugaciliata)、北柴胡(Bupleurumchinense)、異鱗薹草(Carexheterolepis)等不同生活型種,使得樣地分類多樣性指數較大。
坡位主要反映的是立地條件、水分狀況以及人為干擾等情況;坡位與Δ+顯著正相關(P<0.05),與Λ+極顯著正相關(P<0.01)。趙小娜等[19]研究結果表明坡位與Δ+正相關(P>0.05),與Λ+負相關(P>0.05),是由于研究區域位于龐泉溝自然保護區,海拔1600—2400 m,溫度變化明顯,且受人為活動干擾較少,Δ+和Λ+主要受海拔梯度的影響。本文研究的油松林海拔分布相對較低(744—1800 m),熱量條件相對較好,人為活動干擾較多,林下的物種組成相對簡單。群落物種數與Δ+和Λ+呈極顯著負相關(P<0.01);密度與Λ+顯著正相關(P<0.05);胸徑與Δ+顯著正相關(P<0.05)。如位于寧武接官亭林場的樣地S102物種數最多,共43種,屬2門3綱19目24科38屬,喬木層密度較小郁閉度低(0.60),林下種類豐富,灌木層優勢種為土莊繡線菊,草本優層優勢種為披針薹草和東方草莓(Fragariaorientalis),Δ+和Λ+值相對較低。位于武鄉縣石門鄉石門村的樣地S32物種數最少,僅出現油松、披針薹草、茜草、小紅菊(Dendranthemachanetii)、白蓮蒿(Artemisiasacrorum)、牛尾蒿(Artemisiadubia)等,植物但它們分屬2門3綱5目5科6屬,因此表現出較高的Δ+和Λ+值。
微地形因子和群落結構因子導致物種的微生境差異。已有研究表明,與微生境相比地形因子對分類學多樣性影響較小[11],本文的結果與此類似。群落結構因子對分類學多樣性影響最大可能是因為群落演替早期具有若干生態位相似或對環境有相似響應的近緣種;隨著演替的進行,物種對水分、光照等環境因子的需求增多,種間對相似資源的激烈競爭使近緣種逐漸減少,而使親緣關系較遠的物種得以實現穩定共存,群落趨向成熟,并最終達到頂級群落階段。
致謝:感謝山西大學黃土高原研究所蘇智嬌、趙蓉、張淼淼、李旭華、鄧永利、毛空、王潔等同學參與野外調查。
[1] Neeson T M, Van Rijn I, Mandelik Y. How taxonomic diversity, community structure, and sample size determine the reliability of higher taxon surrogates. Ecological Applications, 2013, 23(5): 1216- 1225.
[2] Jucker T, Carboni M, Acosta A T R. Going beyond taxonomic diversity: Deconstructing biodiversity patterns reveals the true cost of iceplant invasion. Diversity and Distributions, 2013, 19(12): 1566- 1577.
[3] 秦曉娟, 董剛, 鄧永利, 毛空, 李旭華, 張峰. 山西平陸黃河濕地植物分類學多樣性研究. 生態學報, 2015, 35(2): 409- 415.
[4] 蔣志剛, 紀力強. 鳥獸物種多樣性測度的G-F指數方法. 生物多樣性, 1999, 7(3): 220- 225.
[5] Moreno C E, Castillo-Campos G, Verdú J R. Taxonomic diversity as complementary information to assess plant species diversity in secondary vegetation and primary tropical deciduous forest. Journal of Vegetation Science, 2009, 20(5): 935- 943.
[6] Clarke K R, Warwick R M. A taxonomic distinctness index and its statistical properties. Journal of Applied Ecology, 1998, 35(4): 523- 531.
[7] Graham N A J, McClanahan T R, Letourneur Y, Galzin R. Anthropogenic stressors, inter-specific competition and ENSO effects on a Mauritian coral reef. Environmental Biology of Fishes, 2007, 78(1): 57- 69.
[8] Rogers S I, Clarke K R, Reynolds J D. The taxonomic distinctness of coastal bottom-dwelling fish communities of the North-east Atlantic. Journal of Animal Ecology, 1999, 68(4): 769- 782.
[9] Ceschia C, Falace A, Warwick R. Biodiversity evaluation of the macroalgal flora of the Gulf of Trieste (Northern Adriatic Sea) using taxonomic distinctness indices. Hydrobiologia, 2007, 580(1): 43- 56.
[10] 徐賓鐸, 金顯仕, 梁振林. 黃海魚類群落分類學多樣性的研究. 中國海洋大學學報: 自然科學版, 2005, 35(4): 629- 634.
[11] 李永振, 史赟榮, 艾紅, 董麗娜, 李娜娜, 李夏, 高天翔. 南海珊瑚礁海域魚類分類多樣性大尺度分布格局. 中國水產科學, 2011, 18(3): 619- 628.
[12] Richard M. The use of taxonomic distinctness to assess environmental disturbance of insect communities from running water. Freshwater Biology, 2007, 52(8): 1634- 1645.
[13] Mouillot D, Laune J, Tomasini J A, Aliaume C, Brehmer P, Dutrieux E, Do Chi T. Assessment of coastal lagoon quality with taxonomic diversity indices of fish, zoobenthos and macrophyte communities. Hydrobiologia, 2005, 550(1): 121- 130.
[14] 丁蘭平. 兩個新的群落多樣性指數. 海洋科學, 1999, 21(1): 25- 28.
[15] Leonard D R P, Clarke K R, Somerfield P J, Warwick R M. The application of an indicator based on taxonomic distinctness for UK marine biodiversity assessments. Journal of Environmental Management, 2006, 78(1): 52- 62.
[16] Ellingsen K E, Clarke K R, Somerfield P J, Warwick R M. Taxonomic distinctness as a measure of diversity applied over a large scale: the benthos of the Norwegian continental shelf. Journal of Animal Ecology, 2005, 74(6): 1069- 1079.
[17] 李景文, 李俊清, 任艷林. 海南主要熱帶森林生態系統類型自然保護區物種分類多樣性研究. 北京林業大學學報, 2002, 24(5/6): 108- 114.
[18] Da Silva I A, Batalha M A. Taxonomic distinctness and diversity of a hyperseasonal savanna in central Brazil. Diversity and Distributions, 2006, 12(6): 725- 730.
[19] 趙小娜, 秦曉娟, 董剛, 張峰. 龐泉溝自然保護區植物群落分類學多樣性. 應用生態學報, 2014, 25(12): 3437- 3442.
[20] 陳姣, 廉凱敏, 張峰, 王建軍, 史榮耀. 山西歷山保護區野生種子植物區系研究. 山西大學學報: 自然科學版, 2012, 35(1): 151- 157.
[21] 劉蕾, 張峰. 歷山白云洞景區植物群落物種多樣性研究. 山西大學學報: 自然科學版, 2010, 33(3): 468- 472.
[22] 狄曉艷. 油松遺傳多樣性與光合生理生態特性研究[D]. 太原: 山西大學, 2014.
[23] 張峰. 山西油松林分布區氣候因素的排序研究. 山西大學學報: 自然科學版, 1990, 13(3): 322- 327.
[24] 張新波, 李悅, 袁虎威, 李偉, 富裕華, 劉致遠, 朱松林. 山西油松天然林分21年子代生長性狀遺傳變異研究. 北京林業大學學報, 2014, 36(3): 104- 109.
[25] 方精云, 王襄平, 沈澤昊, 唐志堯, 賀金生, 于丹, 江源, 王志恒, 鄭成洋, 朱江玲, 郭兆迪. 植物群落清查的主要內容、方法和技術規范. 生物多樣性, 2009, 17(6): 533- 548.
[27] Clarke K R, Warwick R M. The taxonomic distinctness measure of biodiversity: weighting of step lengths between hierarchical levels. Marine Ecology Progress Series, 1999, 184: 21- 29.
[28] 曾燏. 嘉陵江干流魚類物種分類多樣性研究. 西華師范大學學報: 自然科學版, 2012, 33(3): 246- 260.
[29] Clarke K R, Warwick R M. A further biodiversity index applicable to species lists: variation in taxonomic distinctness. Marine Ecology Progress Series, 2001, 216(2): 265- 278.
[30] Ricotta C, Avena G C. An information-theoretical measure of taxonomic diversity. Acta Biotheoretica, 2003, 51(1): 35- 41.
[31] Pienkowski M W, Watkinson A R, Kerby G, Warwick R M, Clarke K R. Taxonomic distinctness and environmental assessment. Journal of Applied Ecology, 1998, 35(4): 532- 543.
[32] 李圣法. 東海大陸架魚類群落生態學研究-空間格局及其多樣性[D]. 上海: 華東師范大學, 2005.
Taxonomic diversity ofPinustabuliformiscommunities in Shanxi
WANG Ye1, QIN Hao1, DONG Gang1,2, QIN Xiaojuan2, ZHAO Xiaona1, ZHANG Feng1,2,*
1InstituteofLoessPlateau,ShanxiUniversity,Taiyuan030006,China2SchoolofLifeScience,ShanxiUniversity,Taiyuan030006,China
Taxonomic diversity based on the phylogenetic system directly reflects species diversity with evolutionary information, considering evolutionary and taxonomic hierarchies among species in a community. It can also indirectly indicate if community structure and components are affected by habitat and disturbance. Thus, it can indicate whether a community or ecosystem is degraded. Furthermore, taxonomic diversity is robust and minimally influenced by different sampling methods and sampling areas compared to the traditional species diversity indices (e.g., Shannon index).Pinustabulaeformisis an important specie of timbers and ecological trees in North China, and distributed widely in Shanxi. To explore the taxonomic diversity ofP.tabuliformiscommunities in Shanxi, we investigated 113 stands in Shanxi, in which 357 species belonging to 227 genera, 71 families, 36 orders, 4 classes, and 3 phyla were recorded. The compositions of genera and species within families exhibited differences. The dominant family including the most genera and species was Compositae (31 genera and 50 species). Most of the species belongs to the genusArtemisia(11 species). In addition, we selected the average taxonomic difference index (Δ+) and taxonomic difference variation index (Λ+) to measure taxonomic diversity, then discussed the relationships between taxonomic diversity indices and environmental factors. The results indicated that the expected values ofΔ+andΛ+were 53 and 280, respectively. Based on the distributions of 113 stands in the confidence funnel, theΔ+value of 89.5% and all theΛ+values were above the expected values ofΔ+andΛ+, which suggested that species composition of theP.tabuliformiscommunities had more distant relatives and belonged to different taxa. The evenness ofP.tabuliformiscommunities was low, given that most species belonged to a small number of families (e.g., Compositae, Rosaceae, Leguminosae, Gramineae). The community factors (DBH, density, and number of species) and microtopography factors (aspect and slope position) were the most important factors influencing the taxonomic diversity index, followed by macrotopography factors (latitude and altitude). DBH had a highly significant and positive correlation withΔ+andΛ+(P<0.01); the relationship between slope, aspect, andΔ+was highly significant (P<0.01). The slope position had a significant correlation withΔ+(P<0.05) and highly significant correlation withΛ+(P<0.01). The relationships betweenΔ+, total nitrogen, and soil organic carbon were highly significant and positive correlations (P<0.01). The correlation of macrotopography factors withΔ+andΛ+was not significant (P> 0.05). We concluded that mostP.tabuliformiscommunities in Shanxi were in the steady growth stage. The species composition was relatively stable, although there were taxonomic differences to some degree within species distributed mainly in a few of the larger taxa.
Shanxi;Pinustabuliformiscommunities; average taxonomic difference index (Δ+); Taxonomic difference variation index (Λ+)
山西省自然科學基金項目(2013011037- 1);山西省回國留學人員科研項目(20100012);科技部科技基礎性工作專項(2011FY110300)
2015- 02- 04;
日期:2016- 01- 22
10.5846/stxb201502040284
*通訊作者Corresponding author.E-mail: fzhang@sxu.edu.cn
王燁,秦浩,董剛,秦曉娟,趙小娜,張峰.山西油松林分類學多樣性.生態學報,2016,36(20):6520- 6527.
Wang Y, Qin H, Dong G, Qin X J, Zhao X N, Zhang F.Taxonomic diversity ofPinustabuliformiscommunities in Shanxi.Acta Ecologica Sinica,2016,36(20):6520- 6527.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
