生姜中Salmonella enterica的分離與宿主氧化應答
2016-11-14 08:38:57李泊寧劉奕清
食品科學 2016年7期
黃 科,李泊寧,羅 潔,龐 敏,劉奕清,鄒 勇,
(1.重慶文理學院林學與生命科學學院,重慶 402160;2.重慶文理學院特色植物研究所,重慶市特色種苗工程技術研究中心,重慶高校園林花卉工程研究中心,重慶 402160)
生姜中Salmonella enterica的分離與宿主氧化應答
黃 科1,2,李泊寧1,羅 潔1,2,龐 敏1,劉奕清1,2,鄒 勇1,2,*
(1.重慶文理學院林學與生命科學學院,重慶 402160;2.重慶文理學院特色植物研究所,重慶市特色種苗工程技術研究中心,重慶高校園林花卉工程研究中心,重慶 402160)
本研究對重慶榮昌采后發病姜塊進行病原菌分離鑒定,分離到一株新的生姜致病菌。通過細菌形態、生理生化、16S rDNA遺傳分析鑒定為沙門氏菌(Salmonella enterica)。將沙門氏菌回接姜苗、姜塊,發現姜苗葉片失綠,褶皺;姜塊表面水漬并有分泌物;動態污染研究表明:沙門氏菌能在姜塊中迅速繁殖,顯著提高姜塊超氧化物歧化酶(superoxide dismutase,SOD)、過氧化物酶(peroxidase,POD)、過氧化氫酶(catalase,CAT)的活性。該研究表明,沙門氏菌能以生姜為寄主存活。
生姜;分離;沙門氏菌;抗氧化應答
生姜是一種重要的藥食同源經濟作物,可鮮食,也可作為調味。但生姜在營養繁殖時體內積累大量病原微生物(細菌和真菌)導致生姜在生產和貯藏過程中發生塊莖腐爛、葉片枯萎,葉斑、干枯病癥[1]。研究認為生姜的主要病害包括:熒光假單胞桿菌(Pseudomonas fluorescens)引起的“姜溫病”[2];歐文氏桿菌(Erwinia chrysanthemi)引發的生姜莖腐病[3];腸桿菌(Enterobacter cloacae)引起的塊莖腐爛[4]。短小芽孢桿菌(Bacillus pumilus)也能引起生姜發病[5];真菌腐霉菌屬(Pythium myriotylum)引起生姜軟腐病[6];鐮刀菌屬(Fusarium oxysporum)可引起葉片變黃,幼芽腐化,長期感染,可致植株死亡[7]。
沙門氏菌包含乍得沙門氏菌(S. bongor)和腸桿沙門氏菌(S. enterica),兩者均能入侵宿主細胞,但只有S. enterica能引起致病[8]。S. enterica以水果和蔬菜蔬為寄主感染人后,引發胃腸疾病[9-11],如以番茄為寄主[12];S. enterica也能侵染植物引起發病:S. enterica Typhimuriumjish寄生于萵苣葉柄周圍,隨著葉片的發育,感染加劇[13],S. typhimuriumjish感染可引起水果和蔬菜軟化腐爛[14],S. enterica通過番茄種子傳播,引發番茄在生長過程中發病[15]。還未見S. enterica引起生姜發病的報道,本研究從重慶榮昌采后生姜中分離一株生姜致病菌,對其進行生理生化,16S rDNA遺傳分析鑒定,并將致病菌回接生姜,觀察致病菌在生姜中的增殖能力及感染癥狀。通過對生姜致病菌分離鑒定研究,以期為生姜病害及人類胃腸道疾病預防提供新的信息。
1 材料與方法
1.1 材料與試劑
生姜樣品 重慶市榮昌縣盤龍鎮四川竹根姜栽培主產區。
酶聯免疫吸附試劑盒 南京建成生物工程研究所;脫毒生姜營養袋苗 重慶市天沛農業科技有限公司饋贈。
1.2 培養基
營養肉湯瓊脂(nutrient agar,NA)培養基:NaCl 5 g、蛋白胨10 g、牛肉膏3 g、瓊脂15 g,加水1 000 mL,調節pH值至7.3;肉湯(nutrient broth,NB)培養基:NaCl 5 g、蛋白胨10 g、牛肉膏3 g,加水1 000 mL,調節pH值至7.3。
1.3 儀器與設備
S3400N掃描電子顯微鏡 日本Hitachi公司;酶標儀美國Bio-Rad公司;聚合酶鏈式反應(polymerase chain reaction,PCR)基因擴增儀 德國Eppendorf公司。
1.4 細菌的分離與純化
發病樣品用清水沖洗,去除表面污漬,然后用無菌水沖洗2 min,重復3 次,用0.6% NaClO浸泡3 min,用無菌水沖洗5 min,重復3 次。用無菌刀挑出生姜腐爛組織,無菌水中浸泡15 min。將浸泡液劃線接種于NA培養基,25 ℃無菌培養48 h后觀察菌落生長狀況。挑取單個菌落,NA培養基上連續傳代培養3 代,純化菌株。將純化后的菌株置于NB培養基25 ℃培養24 h,致病菌株用20%甘油于-80 ℃保存備用。
1.5 致病菌形態觀察
致病菌形態鑒定參照Akin等[16]方法,細菌經NB培養基25 ℃培養24 h。經12 000 r/min離心1 min收集菌體,樣品用2.5%戊二醛(pH 7.4磷酸鹽緩沖溶液(phosphate buffered solution,PBS))固定1 h,然后經30%、50%、70%、80%、90%、100%乙醇脫水、噴金,樣品用S3400N掃描電子顯微鏡在15 kV觀察。
1.6 細菌生理生化實驗
菌株的生理生化特征通過以下實驗來測定:革蘭氏染色、鞭毛染色、莢膜染色、明膠實驗、V.P.實驗、甲基紅實驗、吲哚反應、三糖三醇實驗、賴氨酸脫羧酶實驗、尿素酶實驗、三糖鐵瓊脂實驗[17]。其中,三糖三醇實驗將乳糖、麥芽糖、纖維二糖,甘露醇、山梨醇、甜醇配制為質量濃度10 g/100 mL溶液滅菌后分別加入培養基中,三糖三醇最終質量濃度為1 g/100 mL,培養基配方為NH4H2PO41 g、MgSO4g7H2O 0.2 g、KCl 0.2 g、酵母膏0.2 g、瓊脂5 g,質量濃度0.04 g/100 mL溴甲酚紫乙醇溶液20 mL,蒸餾水1 000 mL,pH 7.2,將菌株穿刺接入培養基中37 ℃培養24 h,觀察菌株生長情況,每個實驗設3 個重復。
1.7 致病菌16S rDNA遺傳分析
采用細菌基因組DNA小提試劑盒提取致病菌基因組DNA。用E. coli 16S rDNA保守序列,正向引物P1:5′-AGAGTTTGATCCTGGCTCAG-3′,反向引物P2:5′-ACGGCTACCTTGTTACGACTT-3′[18],進行致病菌的16S rDNA擴增。PCR反應體系為10hBuffer 5 μL,2.5 mmol/L dNTP 3 μL,P1引物1 μL,P2引物1 μL,2.5 mmol/L MgCl23.5 μL,Taq酶(5 U/μL)0.5 μL,總DNA 1 μL,加入ddH2O 35 μL,總反應體積為50 μL。PCR擴增產物進行電泳檢測、回收、克隆、測序鑒定。測序后16S rDNA在美國國立生物技術信息中心(National Center for Biotechnology Information,NCBI,http://blast. ncbi.nlm.nih.gov/Blast.cgi)中進行比對。使用MEGA 5.1軟件中的鄰接法(neighbor joining,NJ)[19],以E. coli為外群建立系統發育樹。
1.8 S. enterica回接生姜姜塊、姜苗
致病菌經NB培養基25 ℃培養12 h,測定OD600nm值約為1.0時,制備109CFU/mL的細胞懸液。將成熟健康生姜用清水洗凈,用70%乙醇對塊莖表面消毒5 min,用無菌水沖洗,晾干。用無菌刀片將塊莖塊切割為2.5 cmh1.5 cm的小塊。在姜塊上接種1 mL 109CFU/mL的細胞懸液,無菌水作對照,25 ℃培養箱中培養。脫毒生姜組培苗經穴盤移栽后[20]轉至10 cmh10 cm營養袋;穿刺接種0.5 mL 109CFU/mL的細胞懸液,無菌水作對照,溫室培養。
1.9 生姜動態污染
將成熟健康生姜用清水洗凈,用70%乙醇對塊莖表面消毒5 min,之后無菌水沖洗,晾干。用無菌刀片將塊莖切割為1.5 cmh1.5 cm的小塊,在塊莖上劃5 mm深,3 mm寬傷口,在傷口上接種10 μL 109CFU/mL的細胞懸液,S. enterica在生姜上污染動態測定實驗參照Liu Jia等[21]的研究報道。細胞懸液接種在生姜2 h為起始點,每隔24 h從傷口上切取1 cm2、1 cm深樣品,每次切取20 塊樣品,樣品經研磨,10 mL無菌水混合均勻后,200 μL涂布于NA培養基25 ℃,70%濕度培養2 d。測定每個傷口的菌落數,每個實驗重復3 次,每次實驗含20 個姜塊。
1.10 宿主抗氧化酶活性測定
細菌經NB培養基,25 ℃培養12 h后,OD600nm值約1.0時,制備109CFU/mL的細胞懸液。將成熟健康生姜用清水洗凈,用70%乙醇對塊莖表面消毒5 min,之后無菌水沖洗,晾干。用無菌刀將塊莖切割為2.5 cmh1.5 cm的小塊。在姜塊上接種1 mL 109CFU/mL的細胞懸液,每次接種60 塊生姜,實驗重復3 次。接種后于25.5 ℃、70%相對濕度連續培養6 d。每隔24 h取樣品一次,每次取10 塊生姜,-80 ℃保存。連續取樣6 d。分別取2 g材料于預冷的研缽中,加入8 mL預冷的50 mmol/L、pH 7.8 PBS(先加2 mL,在冰浴下研磨成勻漿后,將勻漿轉入10 mL離心管,再用6 mL沖洗),10 000 r/min離心15 min,取上清液定容至10 mL后于4 ℃保存。上清液用于過氧化氫酶(catalase,CAT)、超氧化物歧化酶(superoxide dismutase,SOD)及過氧化物酶(peroxidase,POD)活性測定。采用雙抗一步夾心法酶聯免疫吸附試劑盒測定氧化酶活性。
2 結果與分析
2.1 S. enterica的形態及生理生化特性
分離的S. enterica經NA培養基25 ℃培養2 d后的形態如圖1A所示,菌落為不規則圓形,表面光滑、凸起、白色。掃描電鏡結果(圖1B)表明,單個克隆直徑約1.5 μm,長約13 μm。致病菌生理生化研究表明S. enterica為G-,含鞭毛,無莢膜,V.P.實驗為陰性,不產生吲哚,甲基紅實驗為陽性,不能液化明膠,不分解尿素,不發酵乳糖,但能利用麥芽糖、纖維二糖、甘露醇、甜醇、山梨醇,三糖鐵瓊脂實驗為陽性,賴氨酸脫羧酶反應為陽性結果如表1所示,生姜來源致病菌與已有S. enterica生理生化特性研究報道一致[22]。

圖1S. enterica 單個克隆在LB培養基形態(A)和掃描電鏡形態(×30 000)(B)Fig.1 Morphology of S. enterica colonies on NA agar medium (A) and morphology of a single cell of S. enterica observed with SEM (×30 000) (B)
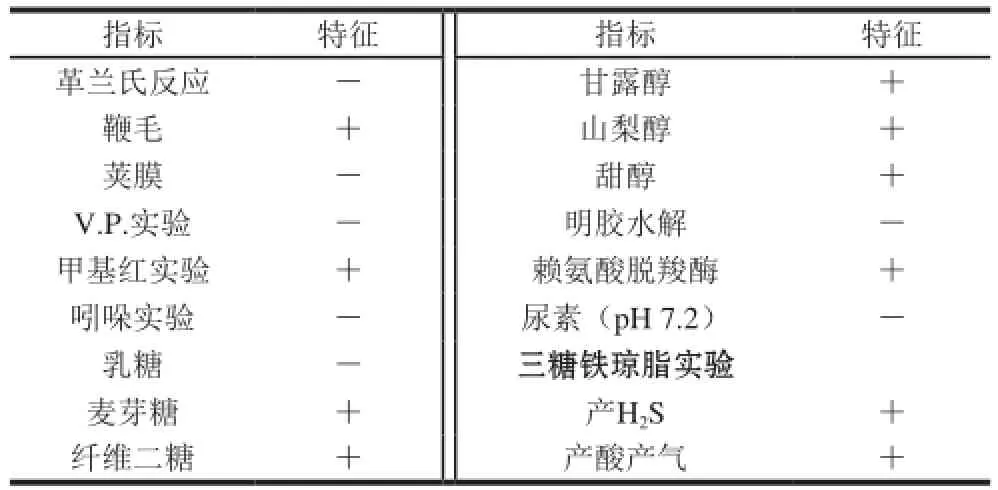
表1S. enterica 的生理生化鑒定結果Table 1 Physiological and biochemical characteristics of S. enterica
2.2 S. enterica的16S rDNA遺傳分析鑒定
以致病菌菌株的總DNA為模板,以16S通用引物進行PCR擴增,PCR產物經克隆、測序序列與GenBank中的核酸數據進行對比分析,結果表明致病菌序列與S. enterica屬細菌相似度達到99%,在此基礎上利用MEGA5.1的鄰近相接法構建S. enterica的系統發育樹,見圖2。對于構建的系統發育樹,采用Bootstrap方法500 個重復進行可靠性檢驗。它和S. enterica亞種具有較近的進化距離。
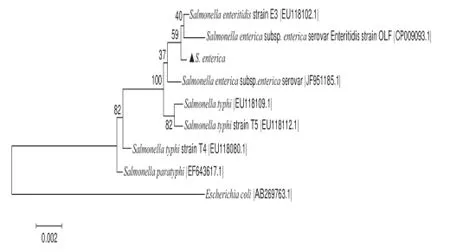
圖2 菌株16S rDNA系統進化樹分析Fig.2 Phylogenetic relationships of S. enterica based on partial 16S rDNA sequences
2.3 S. enterica對生姜的回接及侵染動態
S. enterica回接姜塊經25 ℃、70%濕度培養10 d以后,姜塊表面水漬,并有分泌物滲出,而對照無上述現象(圖3A)。S. enterica經穿刺回接生姜苗3 d后,約30%生姜幼苗葉片發黃,葉片褶皺(圖3B)。S. enterica在回接生姜后,在25 ℃培養時增殖較快(圖4),從接種后到第2天細菌增殖是接種后2 h的3 倍,第3天是第2天的3 倍,然后進入平臺期。這一結果表明S. enterica能以生姜為寄主,并迅速繁殖。
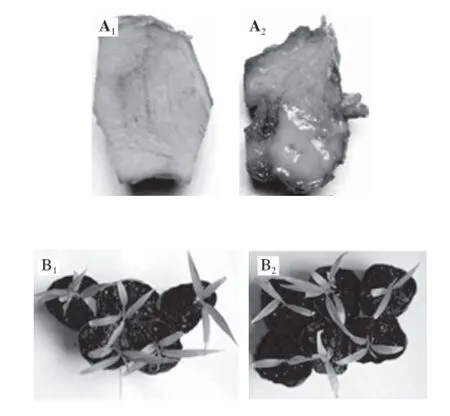
圖3S. enterica回接生姜姜塊(A)和幼苗(B)Fig.3 S. enterica inoculated on ginger rhizome (A) and plant seedlings (B)
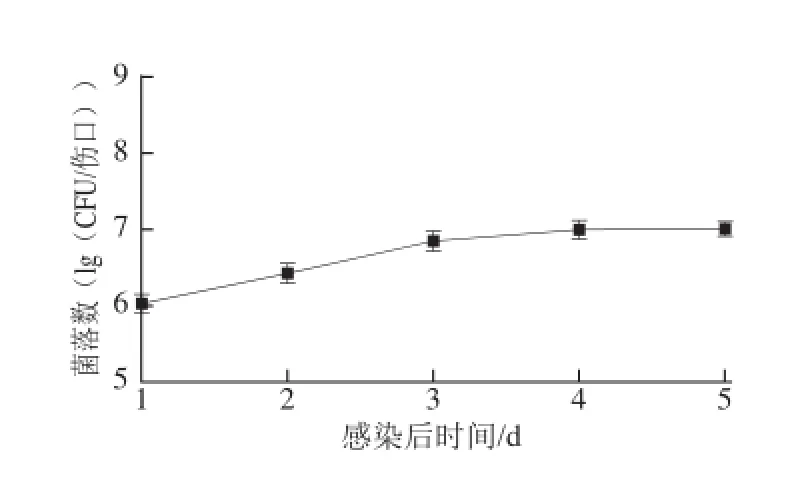
圖4S. enterica對姜塊的動態污染Fig.4 Population dynamics of S. enterica in ginger rhizomes that were wounded, inoculated, and stored at 25 ℃
2.4 S. enterica侵染生姜后對其產生的抗氧化活性
抗氧化酶在激發宿主主動防御系統中起主要作用,由圖5可知,S. enterica侵染宿主24 h后,宿主抗氧化酶SOD、POD、CAT活性均顯著升高,SOD活性升高約3 倍,POD、CAT活性升高約1 倍,隨著侵染時間的增加,SOD、POD活性呈上升趨勢,而CAT活性經1、2、3 d平臺期后再升高。
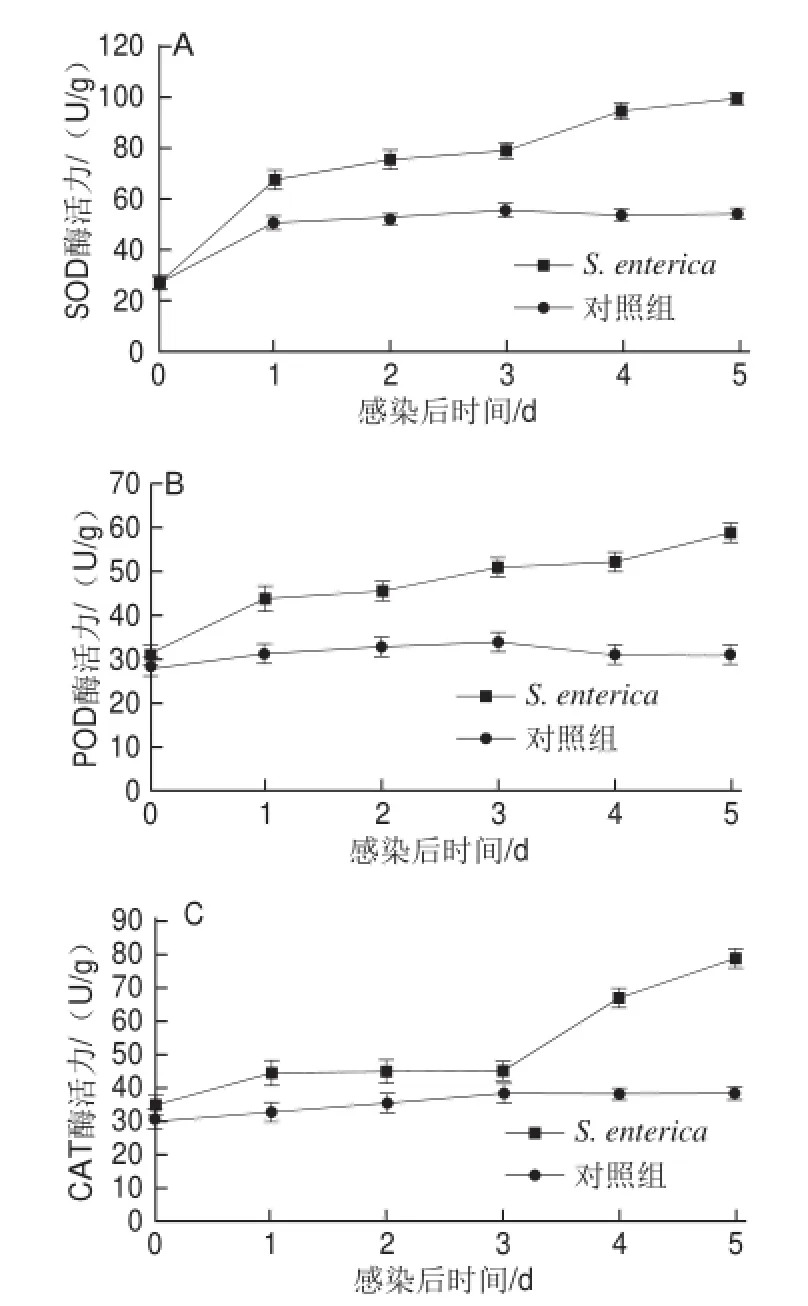
圖5侵染莖塊5 d中SOD(A)、POD(B)和CAT(C)酶活性Fig.5 SOD (A), POD (B) and CAT (C) enzyme activities in ginger rhizomes wounded and inoculated with either S. enterica or sterile,distilled water S. enterica
3 結論與討論
本研究從發病姜塊中分離出S. enterica。人類食物中毒主要是因食入受污染的食物或水,而S. enterica則是引起食物中毒的主要原因。許多研究表明,沙門氏菌可以感染許多植物,主要包括:卷心菜、萵苣、擬南芥、番茄、辣椒、苜蓿、大麥、土豆[11]。但未見沙門氏菌侵染生姜的研究報道。本研究從發病生姜中分離S. enterica,沙門氏菌易在環境中存活,在水和食品中能存活幾月,生姜栽培和生長發育中需大量水和肥,特別是在春季栽培時,畝施用大量農家肥,沙門氏菌可能是在田間施肥、施水時污染姜田,再加上適宜的溫度、濕度加速了沙門氏菌的繁殖,并污染生姜;同時,本研究表明S. enterica回接生姜和姜苗后,姜苗3 d后出現黃葉、褶皺;姜塊回接10 d后表面腐水漬,有分泌物。已有研究報道認為,假單胞菌、腸桿菌、歐文氏桿菌感染生姜塊莖后均能引起發病生姜出現水漬、葉片萎黃變色、塊腐爛等癥狀[2-4]。生姜在生產和貯藏過程中易感染真菌、細菌而發病,特別是在西南地區長江流域高溫高濕天氣,特別是在大雨后,土壤濕度飽和,溫度在25~30 ℃,這種微環境最易使生姜染病[2,23]。而濕度是影響S. enterica繁殖侵染宿主的重要因素,濕度越大,S. montevideo[24]和S. enterica[25]的繁殖速率越快。本研究結果表明S. enterica在姜塊繁殖速率較快,接種1 d后可繁殖3 倍。此外,致病菌侵染宿主通常引起宿主抗氧化酶活性升高,抗氧化酶在激發宿主主動防御系統中起主要作用[26],S. enterica侵染生姜以后,宿主抗氧化酶活力明顯升高。因此,綜合以上結果,本研究從發病生姜中分離S. enterica,這一研究結果將為我國西南地區,特別是重慶地區高溫、高濕的竹根姜產地對生姜病害防治提供了新的信息。此外,S. enterica能引發胃腸疾病,因此本研究也為人類疾病的預防提供了新的信息。
[1] PRABHAKARAN NAIR K P. The agronomy and economy of turmeric and ginger: the invaluable medicinal spice crops. Chapter 21: the diseases of ginger[M]. Boston, Massachusetts, United States: Elsevier, 2013: 409-426. DOI:10.1016/B978-0-12-394801-4.00028-4.
[2] TRUJILLO E E. Diseases of ginger (Zingiber officinɑle) in Hawaii[M]. Hawaii, United States: University of Hawaii, 1964: 1-14.
[3] STIRLING A. Erwinia chrysanthemi, the cause of soft rot in ginger (Zingiber officinɑle) in Australia[J]. Plant Pathology, 2002, 31(4): 419-420. DOI:10.1071/AP02046.
[4] JOLAD S D, LANTZ R C, CHEN G J, et al. Commercially processed dry ginger (Zingiber officinale): composition and effects on LPS-stimulated PGE production[J]. Phytochemistry, 2005, 66: 1614-1635. DOI:10.1016/j.phytochem.2005.05.007.
[5] PENG Q, YUAN Y, GAO M. Bacillus pumilus, a novel ginger rhizome rot pathogen in China[J]. Plant Disease, 2013, 97(10): 1308-1315. DOI:10.1094/PDIS-12-12-1178-RE.
[6] STIRLING G, TURAGANIVALU U, STIRLING A, et al. Rhizome rot of ginger (Zingiber officinɑle) caused by Pythium myriotylum in Fiji and Australia[J]. Australiasian Plant Pathology, 2009, 38(5): 453-460. DOI:10.1071/AP09023.
[7] NISHIJIMA K, ALVAREZ A, HEPPERLY P, et al. Association of Enterobacter cloacae with rhizome rot of edible ginger in Hawaii[J]. Plant Disease, 2004, 88(12): 1318-1327. DOI:10.1094/ PDIS.2004.88.12.1318.
[8] HENSEL M. Salmonella pathogenicity island 2[J]. Molecular Microbiology, 2000, 36(5): 1015-1023. DOI:10.1046/j.1365-2958.2000.01935.x.
[9] BERGER C N, SODHA S V, SHAW R K, et al. Fresh fruit and vegetables as vehicles for the transmission of human pathogens[J]. Environmental Microbiology, 2010, 12(9): 2385-2397. DOI:10.1111/ j.1462-2920.2010.02297.x.
[10] HEATON J, JONES K. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: a review[J]. Journal of Applied Microbiology, 2008, 104(3): 613-626. DOI:10.1111/j.1365-2672.2007.03587.x.
[11] SCHIKORA A, GARCIA A V, HIRT H. Plants as alternative hosts for Salmonella[J]. Trends in Plant Science, 2012, 17(5): 245-249. DOI:10.1016/j.tplants.2012.03.007.
[12] CUMMINGS K, BARRETT E, MOHLE-BOETANI J C, et al. A multistate outbreak of Salmonella enterica serotype Baildon associated with domestic raw tomatoes[J]. Emerging Infectious Diseases, 2001, 7(6): 1046-1048. DOI:10.3201/eid0706.010625.
[13] KROUPITSKI Y, PINTO R, BELAUSOV E, et al. Distribution of Salmonella typhimurium in romaine lettuce leaves[J]. Food Microbiology, 2011, 28(5): 990-997. DOI:10.1016/j.fm.2011.01.007.
[14] WELLS J, BUTTERFIELD J. Salmonella contamination associated with bacterial soft rot of fresh fruits and vegetables in the marketplace[J]. Plant Disease, 1997, 81(8): 867-872. DOI:10.1094/ PDIS.1997.81.8.867.
[15] BARAK J D, LIANG A S. Role of soil, crop debris, and a plant pathogen in Salmonella enterica contamination of tomato plants[J]. Vaccine, 2008, 3(2): 665-671. DOI:10.1016/j.vaccine.2004.11.029.
[16] AKIN D E, BURDICK D, MICHAELS G E. Rumen bacterial interrelationships with plant tissue during degradation revealed by transmission electron microscopy[J]. Applied and Environmental Microbiology, 1974, 27(6): 1149-1156.
[17] 布坎南R E, 吉本斯N E. 伯杰細菌鑒定手冊[M]. 中國科學院微生物研究所《伯杰細菌鑒定手冊》翻譯組, 譯. 8版. 北京: 科學出版社, 1984: 792-812.
[18] SONG J. Phylogenetic analysis of Streptomyces spp. isolated from potato scab lesions in Korea on the basis of 16S rRNA gene and 16S-23S rDNA internally transcribed spacer sequences[J]. International Journal of Systematic & Evolutionary Microbiology, 2004, 54(1): 203-209. DOI:10.1099/ijs.0.02624-0.
[19] TAMURA K, PETERSON D, PETERSON N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology & Evolution, 2011, 28(10): 2731-2739. DOI:10.1093/molbev/ msr121.
[20] 劉奕清, 陳澤雄, 吳中軍. 生姜脫毒種苗移栽基質篩選及肥水調控研究[J]. 北方園藝, 2010(2): 36-37.
[21] LIU J, WISNIEWSKI M, DROBY S, et al. Effect of heat shock treatment on stress tolerance and biocontrol efficacy of Metschnikowia fructicola[J]. FEMS Microbiology Ecology, 2011, 76(1): 145-155. DOI:10.1111/j.1574- 6941.2010.01037.x.
[22] 張河戰. 沙門氏菌的分類、命名及中國沙門氏菌菌型分布[J].微生物學免疫學進展, 2002, 30(2): 74-76. DOI:10.3969/ j.issn.1005-5673.2002.02.018.
[23] STIRLING A M. The causes of poor establishment of ginger (Zingiber officinɑle) in Queensland, Australia[J]. Australasian Plant Pathology, 2004, 33(2): 203-210. DOI:10.1071/AP04003.
[24] ITURRIAGA M H, TAMPLIN M L, ESCARTIN E F. Colonization of tomatoes by Salmonella montevideo is affected by relative humidity and storage temperature[J]. Journal of Food Protection, 2007, 70(1): 30-34.
[25] KINSELLA K J, PRENDERGAST D M, McCANN M S, et al. The survival of Salmonella enterica serovar Typhimurium DT104 and total viable counts on beef surfaces at different relative humidities and temperatures[J]. Journal of Applied Microbiology, 2009, 106(1): 171-180. DOI:10.1111/j.1365-2672.2008.03989.x.
[26] SOTO M J, SANJUáN J, OLIVARES J. Rhizobia and plantpathogenic bacteria: common infection weapons[J]. Microbiology, 2006, 152(6): 3167-3174. DOI:10.1099/mic.0.29112-0.
Salmonella enterica Causes Bacterial Disease in Ginger and Induces Host Antioxidant Response
HUANG Ke1,2, LI Boning1, LUO Jie1,2, PANG Min1, LIU Yiqing1,2, ZOU Yong1,2,*
(1. College of Forestry and Life Sciences, Chongqing University of Arts and Sciences, Chongqing 402160, China; 2. Chongqing Engineering Research Center for Special Plant Seedlings, Garden Flower Engineering Research Center of Chongqing Colleges, Research Institute of Special Plant, Chongqing University of Arts and Sciences, Chongqing 402160, China)
Ginger is an important economic crop with high medicinal and nutritional value. Diseases caused by pathogenic bacteria in ginger have negative impacts on its growth and storage postharvest quality, and often result in significant profit reduction. In the present study, a novel bacterial pathogen, Salmonella enterica, was isolated from postharvest rhizomes in Rongchang, Chongiqng city. The bacterium was identified by 16S rDNA sequence analysis, and morphological, physiological and biochemical tests. After being inoculated with S. enterica, ginger leaves were found to suffer from chlorosis and wrinkle and water and secretion appeared on the surface of rhizomes. Analysis of the population dynamics of S. enterica indicated that the bacterium colonized and grew well on ginger rhizomes. Biochemical analyses indicated that S. enterica activated antioxidant enzyme activities including superoxide dismutase, peroxidase and catalase in host tissues. These results reveal that S. enterica has the capacity to parasitize ginger.
ginger; isolate; Salmonella enterica; antioxidant response
10.7506/spkx1002-6630-201607028
Q93.331
A
1002-6630(2016)07-0152-05
黃科, 李泊寧, 羅潔, 等. 生姜中Salmonella enterica的分離與宿主氧化應答[J]. 食品科學, 2016, 37(7): 152-156. DOI:10.7506/spkx1002-6630-201607028. http://www.spkx.net.cn
HUANG Ke, LI Boning, LUO Jie, et al. Salmonella enterica causes bacterial disease in ginger and induces host antioxidant response[J]. Food Science, 2016, 37(7): 152-156. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201607028. http://www.spkx.net.cn
2015-06-08
永川區自然科學基金項目(Ycstc2013nc8009);重慶文理學院引進人才項目(Z2011RCY02);國家星火計劃項目(2013GA81102)
黃科(1980—),男,講師,博士,研究方向為病原微生物學。E-mail:shanbnm@126.com
*通信作者:鄒勇(1982—),男,講師,博士,研究方向為病原微生物學。E-mail:nevernever107@126.com
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
發明與創新(2016年38期)2016-08-22 03:02:52
