兩株素食者腸道菌產(chǎn)β-葡萄糖苷酶考察及產(chǎn)酶條件優(yōu)化
2016-11-14 08:38:54李笑梅馬慧玲
食品科學(xué) 2016年7期
關(guān)鍵詞:大豆
李笑梅,馬慧玲
(哈爾濱商業(yè)大學(xué) 黑龍江省高校食品科學(xué)與工程重點(diǎn)實(shí)驗(yàn)室,黑龍江 哈爾濱 150076)
兩株素食者腸道菌產(chǎn)β-葡萄糖苷酶考察及產(chǎn)酶條件優(yōu)化
李笑梅,馬慧玲
(哈爾濱商業(yè)大學(xué) 黑龍江省高校食品科學(xué)與工程重點(diǎn)實(shí)驗(yàn)室,黑龍江 哈爾濱 150076)
采用對(duì)硝基苯基-β-D-葡萄糖苷法,與黑曲霉相比較考察素食者腸道菌LJ-G1、LJ-Q2產(chǎn)β-葡萄糖苷酶能力,再經(jīng)單因素、響應(yīng)面試驗(yàn)對(duì)菌種的產(chǎn)酶條件進(jìn)行優(yōu)化。結(jié)果表明:LJ-G1、LJ-Q2菌株均能產(chǎn)β-葡萄糖苷酶,且LJ-Q2產(chǎn)酶能力高于LJ-G1,與黑曲霉相比較,在發(fā)酵時(shí)間短于64 h時(shí)LJ-G1、LJ-Q2產(chǎn)酶能力高于黑曲霉;LJ-Q2菌種的最優(yōu)產(chǎn)酶條件為:培養(yǎng)基pH 8.0、培養(yǎng)溫度38 ℃、培養(yǎng)時(shí)間38 h。在最優(yōu)條件下進(jìn)行驗(yàn)證實(shí)驗(yàn),酶活力達(dá)到1.70 IU/mL。加入大豆異黃酮原料進(jìn)行發(fā)酵培養(yǎng),得到游離大豆異黃酮轉(zhuǎn)化率為39.4%。
素食者;腸道菌;β-葡萄糖苷酶;產(chǎn)酶條件
β-葡萄糖苷酶(β-glucosidase)又叫β-D-葡萄糖苷水解酶,可以將結(jié)合型的大豆異黃酮水解為糖基和游離型的大豆苷元[1-2]。在人體內(nèi)糖苷型的異黃酮不能直接被吸收,而是先被小腸的β-葡萄糖苷酶水解后脫去糖基進(jìn)入血液中,才能發(fā)揮各種生理作用[3]。腸道細(xì)菌在異黃酮的代謝中起到了關(guān)鍵的作用[4],部分益生菌還可以將大豆苷元進(jìn)一步代謝為雌馬酚,與苷元相比它與雌激素受體的結(jié)合力更強(qiáng)[5]。
現(xiàn)今產(chǎn)β-葡萄糖苷酶的菌種研究主要集中在酵母[6]、曲霉、木霉以及細(xì)菌內(nèi)[7]。其中曲霉被普遍認(rèn)為是產(chǎn)β-葡萄糖苷酶的優(yōu)良菌種,尤以黑曲霉的效率最高[5]。但是黑曲霉發(fā)酵到達(dá)產(chǎn)酶高峰期要96~144 h[8-9],工業(yè)化生產(chǎn)周期較長(zhǎng)成本較大。近幾年益生菌產(chǎn)β-葡萄糖苷酶發(fā)酵大豆異黃酮呈現(xiàn)研究熱,有些益生菌[10-11]不僅可以轉(zhuǎn)化糖苷型大豆異黃酮,還有維持腸道、生殖系統(tǒng)、皮膚等正常菌群,抗感染、助降低膽固醇、抑制癌細(xì)胞和增強(qiáng)免疫功能等[12]。LJ-G1、LJ-Q2兩株菌是從素食者糞便中分離出來(lái)的具有代謝大豆異黃酮產(chǎn)雌馬酚的能力[13-14],大豆苷元是此代謝過(guò)程的中間產(chǎn)物之一[1],所以推測(cè)兩種菌株可能有產(chǎn)β-葡萄糖苷酶的能力,而且兩種腸道菌的生長(zhǎng)曲線(xiàn)表明[15],指數(shù)生長(zhǎng)期6~24 h、穩(wěn)定期24~39 h活菌數(shù)達(dá)到最大值。據(jù)此研究探討LJ-G1、LJ-Q2腸道菌產(chǎn)β-葡萄糖苷酶特性及產(chǎn)酶條件優(yōu)化,旨在為豐富產(chǎn)β-葡萄糖苷酶菌種理論知識(shí),也為后續(xù)開(kāi)發(fā)以復(fù)合生物發(fā)酵法制備大豆苷元提供適宜的菌株資源和工藝技術(shù)參數(shù),具有實(shí)際應(yīng)用意義。
1 材料與方法
1.1 菌株與試劑
對(duì)照菌株:黑曲霉(Aspergillus niger)由哈爾濱商業(yè)大學(xué)食品工程學(xué)院微生物實(shí)驗(yàn)室提供;菌株LJ-G1、LJ-Q2從素食者糞便中分離獲得。
對(duì)硝基苯酚、檸檬酸 天津市光復(fù)精細(xì)化工研究所;對(duì)硝基苯基-β-D-吡喃葡萄糖苷 上海金穗生物科技有限公司;所有試劑均為分析純。
1.2 培養(yǎng)基
腦心浸出液培養(yǎng)基(brain heart influsion broth,BHI)青島海博生物技術(shù)有限公司。
麩皮培養(yǎng)基:麩皮0.5%、(NH4)2SO40.4%、K2HPO40.3%、MgSO40.05%、CaCl20.05%、FeSO40.005%、ZnCl20.000 14%、吐溫-80 20 μL,pH 4.8。
1.3 儀器與設(shè)備
Agilent 1200LC高效液相色譜 美國(guó)安捷倫公司;722型光柵分光光度計(jì) 上海精密科學(xué)儀器有限公司;LDZX-30KBS立式壓力蒸汽滅菌器 上海申安醫(yī)療器械廠(chǎng);超凈操作臺(tái) 上海凈化設(shè)備有限公司;YQX-II型培養(yǎng)箱 上海新苗醫(yī)療機(jī)械有限公司。
1.4 方法
1.4.1 產(chǎn)β-葡萄糖苷酶菌株的初篩方法
采用顯色反應(yīng)[16],判斷是否產(chǎn)β-葡萄糖苷酶。用0.1%的剛果紅對(duì)形成單菌落的平板染色處理10 min,再用1 mol/L的NaCl溶液脫色20 min,如果菌落周?chē)a(chǎn)生透明圈說(shuō)明該菌有產(chǎn)β-葡萄糖苷酶的能力。
1.4.2 3 種菌產(chǎn)β-葡萄糖苷酶酶活力比較
采用以對(duì)硝基苯基-β-D-吡喃葡萄糖苷(4-nitrophenylβ-D-glucopyranoside,pNPG)為底物測(cè)定酶活力[17-19]。
1.4.2.1 對(duì)硝基苯酚標(biāo)準(zhǔn)曲線(xiàn)的繪制
用pH 5.0的磷酸氫二鈉-檸檬酸緩沖溶液配制濃度為0、10、20、30、40、50、60 μmol/L的對(duì)硝基苯酚溶液,分別取2 mL上述稀釋后的溶液至10 mL比色管中,于45 ℃水浴鍋中水浴30 min后,加入4 mL 0.5 mol/L的碳酸鈉溶液,混合均勻,于試管架中冷卻至室溫。在400 nm波長(zhǎng)處測(cè)定吸光度(A400nm),繪制標(biāo)準(zhǔn)曲線(xiàn)。
1.4.2.2 培養(yǎng)基中酶活力的測(cè)定
將所試菌種黑曲霉接種于麩皮液體培養(yǎng)基中,28 ℃培養(yǎng)24~144 h;LJ-G1、LJ-Q2腸道菌分別接種于BHI液體培養(yǎng)基中,37 ℃厭氧培養(yǎng)24~144 h,每12 h測(cè)一次酶活力。各培養(yǎng)時(shí)段的培養(yǎng)基經(jīng)3 000 r/min離心20 min,得3 種粗酶液,各吸取2 mL于比色管中,再分別加入10 mL pH 5.0的緩沖液,2 mL 50 mmol/L對(duì)硝基苯基-β-D-吡喃葡萄糖苷溶液。其余實(shí)驗(yàn)過(guò)程同1.4.2.1節(jié)。參比溶液:以2 mL未發(fā)酵培養(yǎng)基代替發(fā)酵培養(yǎng)基。
酶活力定義:每分鐘水解生成1 μmol/L對(duì)硝基苯酚所需要的酶量為一個(gè)酶活力單位。按公式(1)計(jì)算酶活力[20-23]。

式中:X為發(fā)酵培養(yǎng)基中粗酶液酶活力/(IU/mL);c為根據(jù)實(shí)際樣液的吸光度計(jì)算出的對(duì)硝基苯酚濃度/(μmol/L);N為試樣溶液反應(yīng)前的總稀釋倍數(shù);t為反應(yīng)時(shí)間/min;V為酶反應(yīng)液用量/mL。
1.4.3 產(chǎn)酶條件單因素試驗(yàn)
在篩選出產(chǎn)酶優(yōu)勢(shì)菌株的前提下,進(jìn)行產(chǎn)酶單因素試驗(yàn)。固定因素水平設(shè)定為培養(yǎng)基pH 7.0、接種量5%、培養(yǎng)溫度37 ℃、培養(yǎng)時(shí)間36 h,在此基礎(chǔ)條件下,以酶活力為評(píng)價(jià)指標(biāo),選出適宜因素水平范圍。
1.4.3.1 培養(yǎng)基初始pH值對(duì)菌種產(chǎn)酶酶活力的影響
在接種量5%、培養(yǎng)溫度37 ℃、培養(yǎng)時(shí)間36 h條件下,分別考察培養(yǎng)基初始pH 4.0、5.0、6.0、7.0、8.0、9.0、10.0對(duì)菌種產(chǎn)酶活力的影響。
1.4.3.2 培養(yǎng)時(shí)間對(duì)菌種產(chǎn)酶酶活力的影響
在培養(yǎng)基初始pH 7.0、接種量5%、培養(yǎng)溫度37 ℃條件下,分別考察培養(yǎng)時(shí)間在24、36、48、60、72 h時(shí)對(duì)菌種產(chǎn)酶活力的影響。
1.4.3.3 培養(yǎng)溫度對(duì)菌種產(chǎn)酶酶活力的影響
在培養(yǎng)基初始pH 7.0、接種量5%、培養(yǎng)時(shí)間36 h條件下,分別考察培養(yǎng)溫度為33、35、37、39、41 ℃時(shí)對(duì)菌種產(chǎn)酶活力的影響。
1.4.3.4 接種量對(duì)菌種產(chǎn)酶酶活力的影響
在培養(yǎng)基初始pH 7.0、培養(yǎng)溫度37 ℃、培養(yǎng)時(shí)間36 h條件下,分別考察1%、3%、5%、7%、9%接種量對(duì)菌種酶活力的影響。
1.4.4 響應(yīng)曲面優(yōu)化產(chǎn)酶條件
以單因素試驗(yàn)結(jié)果為依據(jù),設(shè)計(jì)培養(yǎng)基初始pH值、培養(yǎng)溫度、培養(yǎng)時(shí)間三因素做響應(yīng)曲面試驗(yàn)。以培養(yǎng)基中的酶活力為指標(biāo),確定最佳產(chǎn)酶條件。
1.4.5 最優(yōu)產(chǎn)酶條件下大豆苷元轉(zhuǎn)化率的測(cè)定
采用高效液相色譜法測(cè)定,固定相為Agilent1200 LC C18柱(250 mmh4.6 mm,5 μm),流動(dòng)相為甲醇-水,0 min 20∶80(V/V,下同),40 min 60∶40,進(jìn)行梯度洗脫,流速為1.2 mL/min,柱溫為40 ℃,檢測(cè)波長(zhǎng)為259 nm。將配制成的0.048、0.096、0.144、0.192、0.24 mg/mL系列質(zhì)量濃度大豆異黃酮混合標(biāo)準(zhǔn)溶液[18]依次進(jìn)樣。利用峰面積繪制標(biāo)準(zhǔn)曲線(xiàn)。根據(jù)公式(2)計(jì)算大豆異黃酮含量。
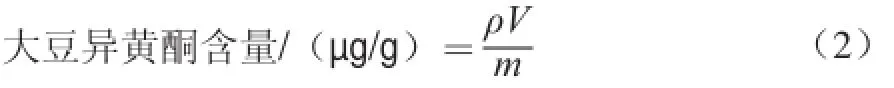
式中:ρ為由各類(lèi)型大豆異黃酮回歸方程得出其質(zhì)量濃度/(μg/mL);m為樣品質(zhì)量/g;V為定容體積/mL。
將菌液接種培養(yǎng)基中,再加入0.3 g大豆異黃酮粉,在最優(yōu)條件下發(fā)酵培養(yǎng),然后培養(yǎng)液經(jīng)3 000 r/min離心30 min,吸取上清液進(jìn)入高效液相色譜進(jìn)行檢測(cè),與未接菌培養(yǎng)基進(jìn)行比較,計(jì)算游離型大豆異黃酮轉(zhuǎn)化率。
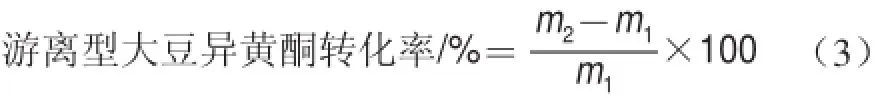
式中:m1為原料中游離型異黃酮含量/mg;m2為發(fā)酵后游離型異黃酮含量/mg。
2 結(jié)果與分析
2.1 產(chǎn)β-葡萄糖苷酶能力顯色反應(yīng)結(jié)果
LJ-G1桿菌、LJ-Q2球菌均產(chǎn)生透明圈,說(shuō)明兩種菌具有產(chǎn)β-葡萄糖苷酶的能力。
2.2 3 種菌產(chǎn)β-葡萄糖苷酶活力比較
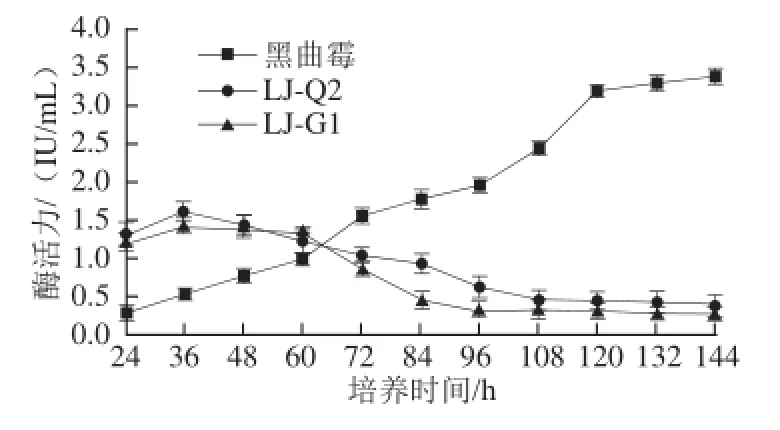
圖1 3 種菌產(chǎn)β-葡萄糖苷酶酶活力的比較Fig.1 Comparison of β-glucosidase activities from three strains
由圖1可知,3 種菌在培養(yǎng)36 h時(shí)均產(chǎn)酶,酶活力最高的是LJ-Q2,達(dá)到1.60 IU/mL。此時(shí)黑曲霉產(chǎn)酶能力最低,隨著培養(yǎng)時(shí)間的增加,黑曲霉產(chǎn)酶能力逐漸增加,而LJ-Q2、LJ-G1產(chǎn)酶能力逐漸降低,當(dāng)培養(yǎng)時(shí)間達(dá)到64 h時(shí),3 種菌產(chǎn)酶能力相同,之后產(chǎn)酶能力從大到小順序?yàn)楹谇梗綥J-Q2>LJ-G1。故以下優(yōu)化試驗(yàn)均用LJ-Q2菌種。
2.3 產(chǎn)酶條件單因素試驗(yàn)
2.3.1 培養(yǎng)基初始pH值對(duì)菌種產(chǎn)酶活力的影響
一銨國(guó)內(nèi)市場(chǎng)高位盤(pán)整,出廠(chǎng)報(bào)價(jià)受成本支撐,仍處高位,價(jià)格暫無(wú)下滑風(fēng)險(xiǎn)。二銨方面,秋季節(jié)市場(chǎng)需求釋放,出口市場(chǎng)較為穩(wěn)定,內(nèi)銷(xiāo)市場(chǎng)仍在加大,估計(jì)后期冬儲(chǔ)市場(chǎng)價(jià)格仍會(huì)有所上揚(yáng)。

圖2 培養(yǎng)基初始pH值對(duì)菌種產(chǎn)酶活力的影響Fig.2 Effect of initial medium pH on β-glucosidase activity
如圖2所示,當(dāng)培養(yǎng)基初始pH值為4.0時(shí)菌株產(chǎn)酶能力較低,當(dāng)pH值為8.0時(shí)產(chǎn)酶能力達(dá)到最高,之后酶活力開(kāi)始下降,表明LJ-Q2在培養(yǎng)基初始pH值為7.0~9.0時(shí)產(chǎn)酶能力相對(duì)較強(qiáng)。
2.3.2 培養(yǎng)時(shí)間對(duì)菌種產(chǎn)酶活力的影響

圖3 培養(yǎng)時(shí)間對(duì)菌種產(chǎn)酶活力的影響Fig.3 Effect of incubation time on β-glucosidase activity
如圖3所示,菌株在培養(yǎng)時(shí)間達(dá)到36 h時(shí)產(chǎn)酶能力達(dá)到最高,之后酶活力隨培養(yǎng)時(shí)間的延長(zhǎng)而下降,表明該菌的最適培養(yǎng)時(shí)間在24~48 h。
2.3.3 培養(yǎng)溫度對(duì)菌種產(chǎn)酶活力的影響
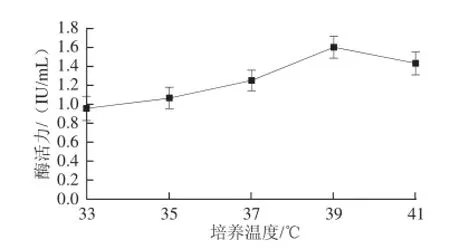
圖4 培養(yǎng)溫度對(duì)菌種產(chǎn)酶活力的影響Fig.4 Effect of culture temperature on β-glucosidase activity
如圖4所示,培養(yǎng)溫度為33 ℃時(shí),菌株產(chǎn)酶能力較低,當(dāng)培養(yǎng)溫度達(dá)到39 ℃時(shí)菌株產(chǎn)酶能力達(dá)到最高,此后隨著溫度升高酶活力下降,表明該菌最適培養(yǎng)溫度為37~41 ℃。
2.3.4 接種量對(duì)菌種產(chǎn)酶活力的影響

圖5 接種量對(duì)菌種產(chǎn)酶活力的影響Fig.5 Effect of inoculum amount on β-glucosidase activity
如圖5所示,接種量1%時(shí),菌株產(chǎn)酶能力較低,隨著接種量的增加,菌株產(chǎn)酶能力隨之增大,當(dāng)接種量達(dá)到5%時(shí),菌株產(chǎn)酶能力達(dá)到最高,此后隨著接種量升高酶活力變化不顯著(P>0.05),表明該菌種最適接種量為5%。
2.4 響應(yīng)面試驗(yàn)結(jié)果分析
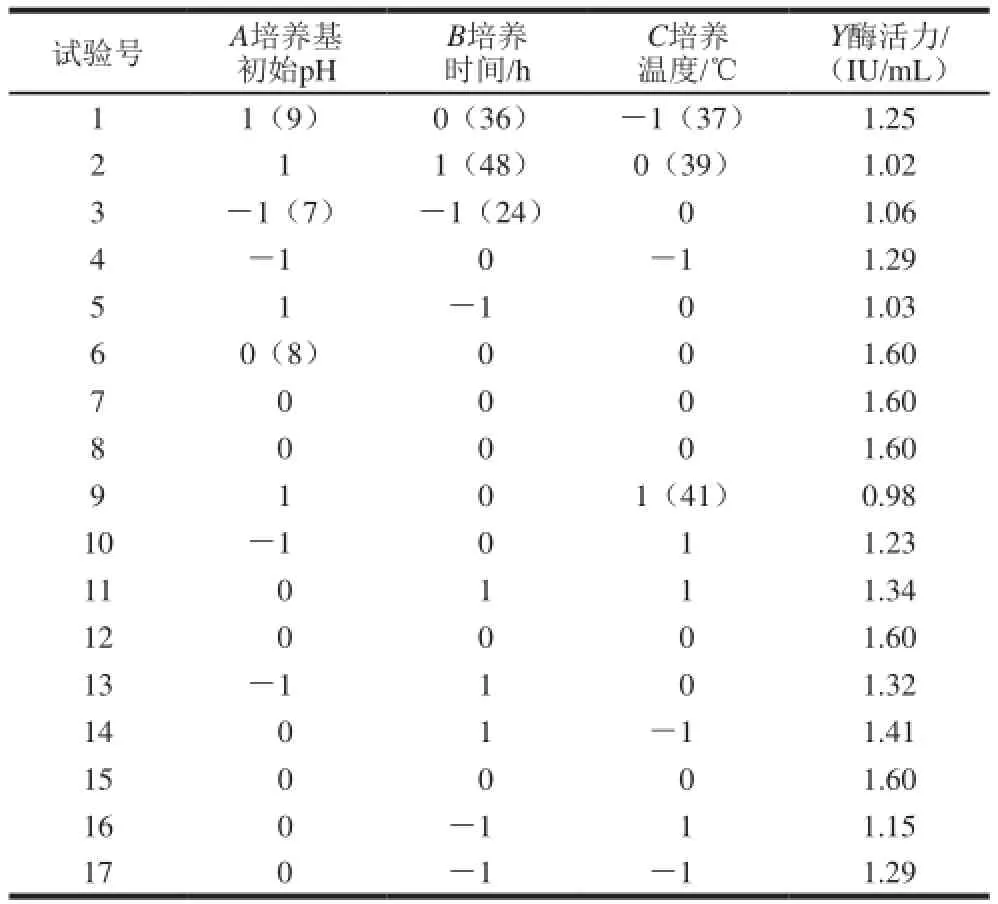
表1 培養(yǎng)條件響應(yīng)面試驗(yàn)方案設(shè)計(jì)及結(jié)果Table 1 Response surface design and results for the optimization of culture conditions
培養(yǎng)條件響應(yīng)面試驗(yàn)各因素和水平見(jiàn)表1,酶活力最高為1.60 IU/mL。產(chǎn)酶條件響應(yīng)面回歸方程及顯著性分析見(jiàn)表2。利用Design-Expert軟件對(duì)表2的響應(yīng)曲面法試驗(yàn)設(shè)計(jì)結(jié)果的數(shù)據(jù)進(jìn)行了多元回歸擬合分析,得到回歸方程:

表2 響應(yīng)面回歸方程系數(shù)及顯著性檢驗(yàn)Table 2 Significance test of all coefficients in the regression model
由表2可知,試驗(yàn)所選用的模型高度顯著(P<0.000 1),模型的校正決定系數(shù)=0.951 1,說(shuō)明該模型能解釋95.11%響應(yīng)值的變化,相關(guān)系數(shù)R=0.993 0,說(shuō)明該模型擬合程度良好,試驗(yàn)誤差小,該模型是合適的。從回歸方程系數(shù)顯著性檢驗(yàn)可知,模型一次項(xiàng)A、B極顯著(P<0.01),C顯著(P<0.05);二次項(xiàng)AB、AC極顯著(P<0.01),BC不顯著(P>0.05),各因素對(duì)酶活力的影響程度排序?yàn)锳>B>C。
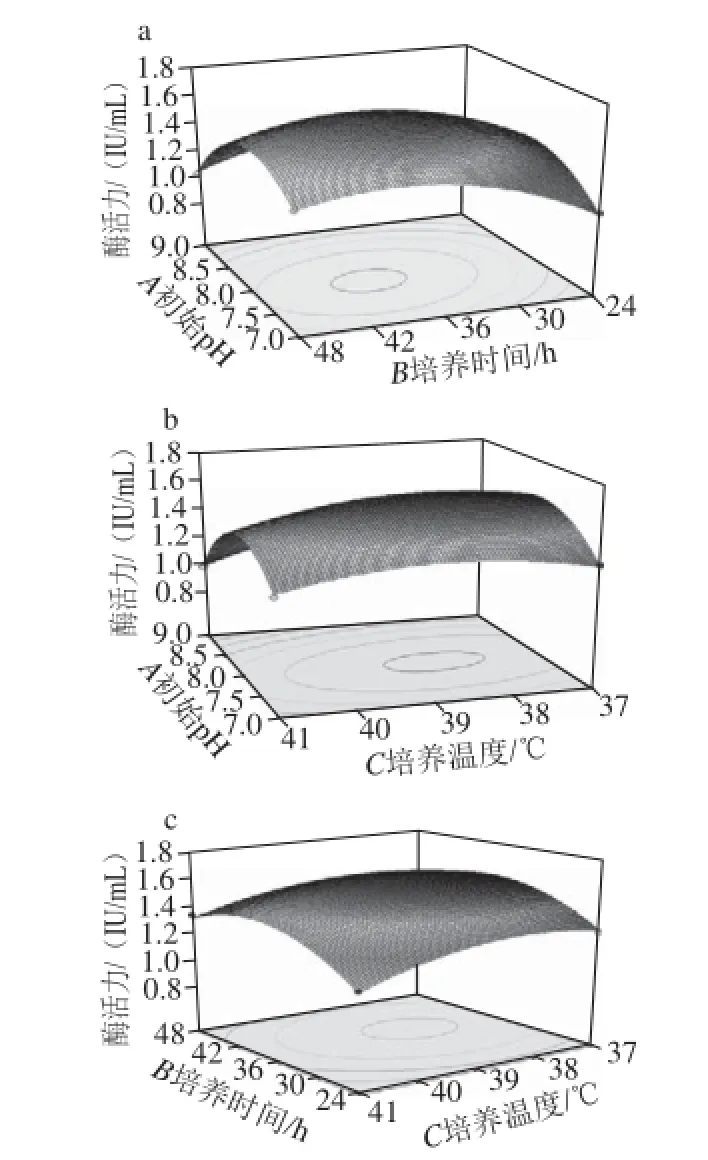
圖6 培養(yǎng)基初始pH值、培養(yǎng)時(shí)間、培養(yǎng)溫度間的交互作用對(duì)酶活力影響的響應(yīng)面圖Fig.6 Response surface plots for the effect of time, initial medium pH and temperature on β-glucosidase activity
由圖6a可知,當(dāng)培養(yǎng)溫度為39 ℃時(shí),隨培養(yǎng)時(shí)間的延長(zhǎng),酶活力逐漸增加,當(dāng)培養(yǎng)時(shí)間在38 h左右時(shí)酶活力達(dá)到最大,之后又慢慢下降。隨著培養(yǎng)基初始pH值的增加,酶活力增大,在培養(yǎng)基初始pH值為8左右時(shí),酶活力為最大,之后又呈下降趨勢(shì)。
由圖6b可知,當(dāng)培養(yǎng)時(shí)間為39 h時(shí),隨著培養(yǎng)基初始pH值的增大,酶活力逐漸增大,當(dāng)培養(yǎng)基初始pH值在8左右時(shí)酶活力達(dá)到最大,之后又慢慢下降。隨著培養(yǎng)溫度的增加,酶活力增大,在培養(yǎng)溫度為38 ℃左右時(shí),酶活力為最大,之后又呈下降趨勢(shì)。
由圖6c可知,當(dāng)培養(yǎng)基初始pH值為8時(shí),隨培養(yǎng)時(shí)間的延長(zhǎng),酶活力逐漸增大,當(dāng)培養(yǎng)時(shí)間在38 h左右時(shí)酶活力達(dá)到最大,之后又慢慢下降。隨著培養(yǎng)溫度的增加,酶活力增大,在培養(yǎng)溫度為38 ℃左右時(shí),酶活力為最大,之后又呈下降趨勢(shì)。
響應(yīng)面產(chǎn)酶條件優(yōu)化結(jié)果為:培養(yǎng)基初始pH 8.04、培養(yǎng)溫度38.31 ℃、培養(yǎng)時(shí)間38.21 h。根據(jù)實(shí)際情況校正值為:培養(yǎng)基pH 8.0、培養(yǎng)溫度38 ℃、培養(yǎng)時(shí)間38 h,在此條件下測(cè)得酶活力為1.7 IU/mL,比未優(yōu)化前酶活力1.60 IU/mL提高了6.2%。
2.5 發(fā)酵培養(yǎng)游離型大豆異黃酮苷元轉(zhuǎn)化率結(jié)果
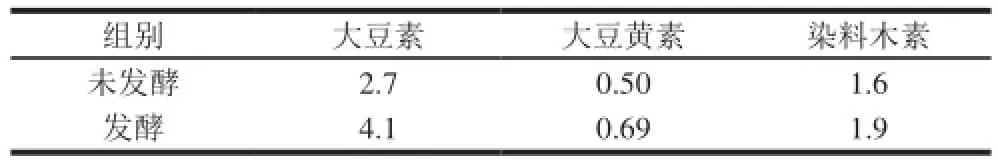
表3 3 種游離型大豆異黃酮含量Table 3 Contents of three soybean isoflavone aglycons before and after fermentation mg
由表3可知,發(fā)酵后3 種游離型大豆異黃酮含量比發(fā)酵前高,共增加了1.89 mg,根據(jù)公式(3)計(jì)算游離型大豆異黃酮轉(zhuǎn)化率為39.4%。
3 結(jié) 論
素食者糞便分離到的腸道菌LJ-G1和LJ-Q2有產(chǎn)β-葡萄糖苷酶的能力,在38 ℃培養(yǎng)36 h時(shí)、產(chǎn)酶能力比黑曲霉高。LJ-Q2菌株響應(yīng)面試驗(yàn)產(chǎn)酶條件優(yōu)化結(jié)果為:培養(yǎng)基初始pH 8.0、培養(yǎng)溫度38 ℃、培養(yǎng)時(shí)間38 h。在此條件下酶活力達(dá)到1.70 IU/mL,比未優(yōu)化前酶活力提高了6.2%。此條件應(yīng)用于發(fā)酵大豆異黃酮其轉(zhuǎn)化率為 39.4%。近幾年對(duì)腸道菌產(chǎn)β-葡萄糖苷酶多有研究。Raimondi等[24]研究了雙歧桿菌屬8 個(gè)菌種的22 個(gè)菌株轉(zhuǎn)化大豆苷的能力,發(fā)現(xiàn)在發(fā)酵7 d內(nèi),除Bifidobacteria longum MB220和Bifidobacteria longum MB224外,其他菌株都能水解大豆苷,其中Bifidobacteria adolescentis MB116在生長(zhǎng)穩(wěn)定期的早期就有最高的酶活力,酶活力達(dá)0.92 U/mg。也有研究表明鏈球菌[25]等細(xì)菌可轉(zhuǎn)化結(jié)合型大豆異黃酮。LJ-Q2腸道菌與黑曲霉相比雖然產(chǎn)酶活力低,但是達(dá)到峰值的時(shí)間短于黑曲霉108 h,以進(jìn)一步轉(zhuǎn)化成雌馬酚,因此,LJ-Q2腸道菌具有很好的應(yīng)用前景。接下來(lái)的工作將探討LJ-Q2腸道菌與其他產(chǎn)β-葡萄糖苷酶的腸道益生菌進(jìn)行復(fù)合,以提高產(chǎn)β-葡萄糖苷酶的能力,應(yīng)用于大豆苷元的制備,這對(duì)今后雌馬酚的轉(zhuǎn)化也有著重要的意義。
[1] 徐茂軍. β-葡萄糖苷酶對(duì)豆奶及豆奶粉中大豆異黃酮糖苷化合物的轉(zhuǎn)化作用研究[J]. 中國(guó)食品學(xué)報(bào), 2005, 5(4): 28-33. DOI:10.3969/ j.issn.1009-7848.2005.04.006.
[2] PYO Y H, LEE T C, LEE Y C. Enrichment of bioactive isoflavones in soymilk fermented with β-glucosidase-producing lactic acid bacteria[J]. Food Research International, 2005, 38: 551-559. DOI:10.1016/j.foodres.2004.11.008.
[3] 孫遜, 姚文, 朱偉云. 腸道大豆異黃酮降解菌研究進(jìn)展[J].世界華人消化雜志, 2006, 14(10): 236-241. DOI:10.3969/ j.issn.1009-3079.2006.10.008.
[4] TSUNCHIHASHI R, SAKAMOTO S, KODERA M, et al. Microbial metabolism of soy isoflavones by human intestinal bacterial strains[J]. Journal of Natural Medicines, 2008, 62: 456-460. DOI:10.1016/S0020-1693(01)00446-7.
[5] TSANGALIS D, WILCOX G, SHAH N P, et al. Urinary excretion of equol by postmenopausal women consuming soymilk fermented by probiotic bifidobacteria[J]. European Journal of Clinical Nutrition,2007, 61(3): 438-441. DOI:10.1038/sj.ejcn.1602530.
[6] 韓婷, 程鋼. β-葡萄糖苷酶以及益生菌生物轉(zhuǎn)化大豆異黃酮糖苷的研究進(jìn)展[J]. 食品科學(xué), 2010, 31(9): 333-337.
[7] MATEO J J, STEFANO R D. Description of the β-glucosidase activity of wine yeasts[J]. Food Microbiology, 1997, 14(6): 583-591. DOI:10.1006/fmic.1997.0122.
[8] 劉小杰. 黑曲霉ZJ1搖瓶發(fā)酵產(chǎn)β-葡萄糖苷酶的研究[J]. 中國(guó)食品添加劑, 2004(3): 27-31. DOI:10.3969/j.issn.1006-2513.2004.03.008.
[9] 魯瑋, 岳冬冬, 劉新育. 黑曲霉產(chǎn)β-葡萄糖苷酶的純化及對(duì)中藥糖苷類(lèi)成分的轉(zhuǎn)化[J]. 中國(guó)醫(yī)藥工業(yè)雜志, 2014, 45(3): 220-223.
[10] 楊守鳳, 徐建雄. 基于復(fù)合益生菌和枯草芽孢桿菌發(fā)酵轉(zhuǎn)化大豆異黃酮的比較研究[J]. 科學(xué)試驗(yàn)與研究, 2014(3): 1-4.
[11] 翟清燕, 趙龍玉, 趙鳳春, 等. 高產(chǎn)大豆異黃酮糖苷轉(zhuǎn)化酶乳酸菌的篩選及其發(fā)酵條件的優(yōu)化[J]. 食品工業(yè)科技, 2014, 35(12): 162-166. DOI:10.13386/j.issn1002-0306.2014.12.026.
[12] 張勇. 世界益生菌產(chǎn)品研究和發(fā)展趨勢(shì)[J]. 中國(guó)微生態(tài)學(xué)雜志, 2009, 21(2): 185-192.
[13] 李笑梅, 賈冰心. 素食者產(chǎn)雌馬酚腸道菌的分離與鑒定[D]. 哈爾濱:哈爾濱商業(yè)大學(xué), 2014: 50-59.
[14] 李笑梅, 賈姚. 兩株素食者腸道菌產(chǎn)雌馬酚特性的研究[D]. 哈爾濱:哈爾濱商業(yè)大學(xué), 2015: 15-39.
[15] 李笑梅, 賈姚. 素食者產(chǎn)雌馬酚腸道菌生長(zhǎng)條件優(yōu)化[J]. 食品科學(xué), 2014, 35(23): 199-203. DOI:10.7506/spkx1002-6630-201423039.
[16] 鄧媛. β-葡萄糖苷酶高產(chǎn)菌株的篩選及對(duì)大豆異黃酮糖苷轉(zhuǎn)化的應(yīng)用研究[D]. 西安: 西北大學(xué), 2011: 17-32.
[17] GUNATA Z, DUGELAY I, VALLIER M J, et al. Multiple forms of glycosidases in an enzyme preparation from Aspergillus niger: partial characterization of a β-glucosidase[J]. Enzyme and Microbial Technology, 1997, 21: 39-44. DOI:10.1016/S0141-0229(96)00221-9.
[18] GRIMALDI A, BARTOWAKY E, JIRANEK V. A survey of glycosidase activities of commercial wine strains of Oencoccusoeni[J]. International Journal of Food Microbiology, 2005, 105: 233-244. DOI:10.1016/j.ijfoodmicro.2005.04.011.
[19] BARBAGALLO R N, SPAGNA G, PALMERI R, et al. Assessment of β-glucosidase activity in selected wild strains of Oencoccusoeni for malolactic fermentation[J]. Enzyme and Microbial Technology, 2004, 34: 292-296. DOI:10.1016/j.enzmictec.2003.11.013.
[20] 孫艷梅. β-葡萄糖苷酶水解大豆異黃酮糖苷的研究[D]. 哈爾濱: 東北農(nóng)業(yè)大學(xué), 2003: 21-30. DOI:10.7666/d.y506393.
[21] GB/T 23788ü2009保健食品中大豆異黃酮的測(cè)定方法 高效液相色譜法[S].
[22] MATEO J J, STEFANO R D. Description of the β-glucosidase activity of wine yeasts[J]. Food Microbiology, 1997, 14(6): 583-591. DOI:10.1006/fmic.1997.0122.
[23] INCECCO N, BARTOWSKY B, KASSARA S, et al. Release of glycosidically bound flavour compounds of chardonnay by Oencoccusoeni during malolactic fermentation[J]. Food Microbiology, 2004, 21: 257-265. DOI:10.1016/j.fm.2003.09.003.
[24] RAIMONDI S, RONCAGLIA L, LUCIA D M, et al. Bioconversion of soy isoflavones daidzin and daidzein by Bifidobɑcterium strains[J]. Applied Microbiology and Biotechnology, 2009, 81(5): 943-950. DOI:10.1007/s00253-008-1719-4.
[25] CHUN J, KIM J S, KIM J H. Enrichment of isoflavone aglycones in soymil by fermentation with single and mixed cultures of Streptococcus infantarius 12 and Weissella sp.4[J]. Food Chemistry, 2008, 109(2): 277-278. DOI:10.1016/j.foodchem.2007.12.024.
Comparison of Two β-Glucosidase-Producing Strains from the Intestinal Tract of Vegetarians and Optimizaiton of Fermentation Conditions for β-Glucosidase Production
LI Xiaomei, MA Huiling
(Key Laboratory of Food Science and Engineering, Harbin University of Commerce, Harbin 150076, China)
The β-glucosidase-producing capacity of strains LJ-G1 and LJ-Q2 isolated from the feces of vegetarians was measured using 4-nitrophenol-β-D-glucoside (pNPG) as substrate and compared with that of Aspergillus niger. The fermentation conditions for β-glucosidase production by the selected strain were optimized using combination of single factor method and response surface methodology. The results showed that both intestinal strains were able to produce β-glucosidase, and LJ-Q2 had higher β-glucosidase-producing capacity than LJ-G1. LJ-G1 and LJ-Q2 had higher β-glucosidase-producing capacity than Aspergillus niger in 64 h of fermentation. The optimal fermentation conditions for β-glucosidase production were determined as follows: culture medium pH, 8.0; temperature, 38 ℃; and time, 38 h. The experimental value of β-glucosidase activity produced under the optimized conditions was 1.70 IU/mL. The fermentation process could convert 39.4% of soybean isoflavones into aglycones.
vegetarians; intestinal bacteria; β-glucosidase; fermentation conditions for β-glucosidase production
10.7506/spkx1002-6630-201607023
TS201.3
A
1002-6630(2016)07-0123-05
李笑梅, 馬慧玲. 兩株素食者腸道菌產(chǎn)β-葡萄糖苷酶考察及產(chǎn)酶條件優(yōu)化[J]. 食品科學(xué), 2016, 37(7): 123-127.
DOI:10.7506/spkx1002-6630-201607023. http://www.spkx.net.cn
LI Xiaomei, MA Huiling. Comparison of two β-glucosidase-producing strains from the intestinal tract of vegetarians and optimizaiton of fermentation conditions for β-glucosidase production[J]. Food Science, 2016, 37(7): 123-127. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201607023. http://www.spkx.net.cn
2015-06-18
黑龍江省科技廳應(yīng)用技術(shù)項(xiàng)目(G013B203);黑龍江省自然科學(xué)基金項(xiàng)目(C201201);黑龍江省高校科技創(chuàng)新團(tuán)隊(duì)建設(shè)計(jì)劃項(xiàng)目(2010td04)
李笑梅(1960—),女,教授,本科,研究方向?yàn)槭称房茖W(xué)。E-mail:lixm0451@163.com
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國(guó)化肥信息(2022年7期)2022-08-31 01:29:28
中國(guó)化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場(chǎng)周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫(huà)報(bào)·智力大王(2015年10期)2016-01-27 01:01:35
讀寫(xiě)算(中)(2015年10期)2015-11-07 07:24:12
