卵白蛋白-油酸復合物對人結腸癌細胞的HAMLET樣生長抑制作用研究
2016-11-08 09:30:27邢效娟劉建壘
食品工業科技 2016年18期
邢效娟,劉建壘,景 浩
(中國農業大學食品科學與營養工程學院,北京 100083)
?
卵白蛋白-油酸復合物對人結腸癌細胞的HAMLET樣生長抑制作用研究
邢效娟,劉建壘,景浩*
(中國農業大學食品科學與營養工程學院,北京 100083)
乳白蛋白與油酸形成的復合物具有癌細胞生長抑制作用(Human alpha-lactalbumin made lethal to tumor cells,HAMLET)。本文制備了卵白蛋白(Ovalbumin,OVA)和油酸(Oleic acid,OA)的復合物(OVA-OA),通過檢測細胞生長抑制、細胞膜通透性、細胞生長周期和凋亡的變化,分析了OVA-OA對人結腸癌細胞(Caco-2)的生長抑制作用以及其中OA對生長抑制作用的貢獻。結果表明,當OVA-OA中OA的濃度達到182 μmol/L時,OVA-OA對Caco-2細胞有顯著(p<0.05)生長抑制作用,273 μmol/L時生長抑制作用增強,且隨著作用時間的延長(1~3 h)而增強。在182 μmol/L時,OA對Caco-2細胞也有生長抑制作用,其劑量效應關系與OVA-OA一致。但當OA濃度為273 μmol/L時,其生長抑制作用弱于OVA-OA的。OVA-OA和OA均會促使乳酸脫氫酶(Lactate dehydrogenase,LDH)釋放,增大晚期凋亡和壞死的細胞百分比,阻滯細胞周期在G0/G1期和S期。由此可知,OVA-OA對結腸癌細胞具有HAMLET樣生長抑制作用,其中OA起主要作用。OVA-OA對人結腸癌細胞的生長抑制作用是通過損傷細胞膜、阻滯細胞生長周期和促進細胞凋亡來實現的。
卵白蛋白,油酸,細胞生長抑制,乳酸脫氫酶,細胞凋亡,細胞周期
Hakansson和Svanborg等[1]發現母乳中含有一種能特異性促使人肺癌細胞凋亡的物質,用蛋白質電泳確定其分子量后發現這種物質的分子量與α-乳白蛋白(α-Lactalbumin,α-LA)相近,但經驗證發現α-LA本身并不能引起肺癌細胞的凋亡,因此當時稱這種物質為乳白蛋白多聚體(multimericα-LA)。經過5年的研究,Svensson等[2]證明這種聚合物是α-LA與油酸(Oleic acid,OA)的復合物,并將其命名為HAMLET。Tolin等[3]通過胃蛋白酶將α-LA進行酶解,再用α-LA肽段與OA混合制備復合物,并通過細胞實驗發現此復合物也有HAMLET樣的細胞生長抑制作用。隨著研究的進行,發現其他類型的蛋白質,比如馬溶菌酶[4]、β-乳球蛋白[5]、乳鐵蛋白[6]也可以與OA形成有HAMLET樣細胞生長抑制作用的復合物,而相對應的蛋白質沒有這種作用。
卵白蛋白(Ovalbumin,OVA)約占蛋清蛋白含量的54%,屬于含磷糖的糖蛋白,分子量為45 ku,等電點為4.5[7]。目前只有一些OVA與不飽和脂肪酸相互作用的報道,例如OA的結合使得OVA的熒光強度降低,二級結構變得更加松散[8],加熱后的OVA可以與亞油酸(Linoleic acid,LA)通過疏水鍵相互作用[9]。但對結合后的OVA-OA的生物活性,尤其是是否具有HAMLET樣的癌細胞生長抑制作用還未見報道。
一般認為α-LA-OA的癌細胞生長抑制作用不是由OA引起的,而是復合物特有的性質。Kamijima等[10]研究α-LA-OA對小鼠淋巴細胞白血病細胞株L1210細胞生長影響時發現即使當OA濃度達到2.5 mmol/L時,也沒有細胞生長抑制作用,而經α-LA-OA作用后細胞生長抑制率為90%左右。Fang等[11]研究α-LA-OA對人結腸癌細胞(Caco-2)和人早幼粒白血病細胞(HL-60)等細胞生長抑制作用時發現OA濃度低于100 μmol/L時,對細胞沒有生長抑制作用。但近年來有些研究者報道,HAMLET中起生長抑制作用的成分為OA,例如α-LA-OA可以抑制HL-60細胞和人宮頸癌細胞(Hela)的生長,但相應濃度的OA(0~100 μmol/L)也有細胞生長抑制作用[12-13]。因此,對于OA以及OA與蛋白質復合物的物化特性和細胞生長抑制作用還需進一步研究。
在本研究中,以人結腸癌細胞(Caco-2)為模型,分析研究OVA-OA的HAMLET樣細胞生長抑制作用以及OA對OVA-OA的細胞生長抑制作用的貢獻。通過LDH法、流式細胞技術分析OVA-OA對細胞膜、細胞生長周期和細胞凋亡的影響。
1 材料與方法
1.1材料與儀器
卵白蛋白(純度≥99%,Ca. No. 1021845)和油酸(純度≥99.5%,Cat. No.112-80-1)均購于成都格雷西亞化學技術有限公司;DMEM(Dulbeco’s modified eagle medium)購于美國Invitrogen公司;胎牛血清(Fetal bovine serum,FBS)購于北京鼎國生物技術有限公司;胰蛋白酶、噻唑藍(Thiazolyl blue tetrazolium bromide,MTT)、不含DNA的RNA酶、碘化丙啶(Propidium iodide,PI)、輔酶I(β-Nicotinamide adenine dinucleotide,NADH)均購于美國Sigma公司;青霉素/鏈霉素溶液購于碧云天生物技術研究所;Annexin V-FITC/PI凋亡試劑盒購于上海貝博生物;其他試劑為國產分析純;實驗用水為高純去離子水(deionized water,18 Ω;dH2O);人結腸癌細胞(Caco-2)購于中國科學院上海生命科學研究院細胞資源中心;細胞培養皿、培養板均購于無錫耐思生物科技有限公司。
Model 680型酶標儀美國BIO-RAD公司;FACSCalibur II型流式細胞儀美國BD公司;MCO-15AC型二氧化碳培養箱日本Sanyo(三洋)公司;BSC-1500 II A2-X型生物安全柜濟南市鑫貝西生物技術有限公司;pHs-3C+型酸度計成都世紀方舟科技有限公司;XB220A型電子天平瑞士Precisa Gravimetrics AG公司;Vortex-Genie 2漩渦振蕩器美國Scientific Industries公司;FM200型實驗室分散乳化機上海弗魯克液體機械制造有限公司。
1.2實驗方法
1.2.1OVA-OA的制備用pH8.0的磷酸鹽緩沖溶液(Phosphate buffer solution,PBS)配制10 mg/mL的OVA儲備液,離心(1460×g,10 min)后取上清4 ℃儲存備用。用無水乙醇溶解OA配制成1 mol/L的儲備液,4 ℃儲存備用;使用前用PBS將OA稀釋為1 mmol/L的工作液,再用乳化機均質(8000~10000 r/min,1 min),制成OA混懸液。將不同體積的OA混懸液加入到含有2 mL,OVA儲備液(10 mg/mL)的離心管中,再加入不同體積的PBS至10 mL,漩渦振蕩均勻,配制成不同摩爾比的OA與OVA的混合溶液,其中OVA濃度恒定為2 mg/mL。再將其置于70 ℃水浴加熱20 min,水浴結束后置于冰水中迅速冷卻,待測。同時將OVA,OA加熱,研究加熱對兩者細胞生長抑制活性的影響。
OVA-OA組中OVA的濃度為2 mg/mL(45.5 μmol/L),OA的濃度分別為0、91、182、273 μmol/L(OA與OVA的摩爾比分別為0∶1、2∶1、4∶1、6∶1),乙醇濃度低于0.5%;OA組濃度分別為0、91、182、273 μmol/L。
1.2.2細胞生長抑制檢測將Caco-2細胞以1×105個/mL的密度接種于96孔板,每孔加入100 μL細胞懸液,置于37 ℃,5% CO2培養箱中培養至細胞覆蓋孔底面積的70%后棄去培養基,每孔加入100 μL的樣品溶液,置于培養箱孵育1、2、3 h后棄去樣品,加入100 μL的DMEM10(含有10%胎牛血清的細胞培養基),于CO2培養箱繼續培養24 h后棄去培養基,每孔加入70 μL含0.5 mg/mL MTT的DMEM,避光孵育3 h后小心棄去上清,加入70 μL鹽酸-異丙醇,在vortex上振蕩(800 r/min,10 min)。測定其在570 nm處吸光值[14-15]。根據公式(1)計算經不同樣品作用后的細胞存活率。
細胞存活率(%)=A樣品570 nm/A對照570 nm×100
式(1)
1.2.3乳酸脫氫酶釋放率檢測Tris-pyruvate-NADH反應液配制:使用前,取100 mL Tris-EDTA緩沖液(pH7.4),分別加入1 mL丙酮酸鈉儲備液(135 mmol/L)和1 mL NADH儲備液(17 mmol/L),混勻,并在37 ℃水浴中孵育15 min以上,備用。
將Caco-2細胞以1×106個/mL的密度接種于直徑為100 mm的細胞培養皿中,每皿加入10 mL的細胞懸液,置于37 ℃,5% CO2培養箱中培養至細胞覆蓋孔底面積的80%~90%后棄去培養基,每孔加入3 mL的樣品溶液,置于培養箱孵育3 h后收集樣品溶液,離心(400×g,10 min)取上清,待測。取70 μL上清液與700 μL預熱(37 ℃)的Tris-pyruvate-NADH反應液(其中Tris,pyruvate,NADH的終濃度分別為50、1.22、0.154 mmol/L)快速混合,立即測定其在340 nm處吸光值,每隔30 s記錄一次,連續記錄3 min,以反應最初吸光值下降最快部分(如0~1 min或0~2 min)求線性回歸方程(時間為橫坐標,吸光值為縱坐標),其斜率絕對值即為每分鐘Abs340 nm的減少值(ΔAbs340 nm)。根據公式(2)計算LDH釋放率[14]。

表1 加熱對OVA與OA對Caco-2細胞的生長抑制作用的影響
注:樣品作用時間為3 h,不同大寫字母A~C表示同列不同均數差異顯著(p<0.05);不同小寫字母a~f表示同行不同均數差異顯著(p<0.05);表2、表3同。
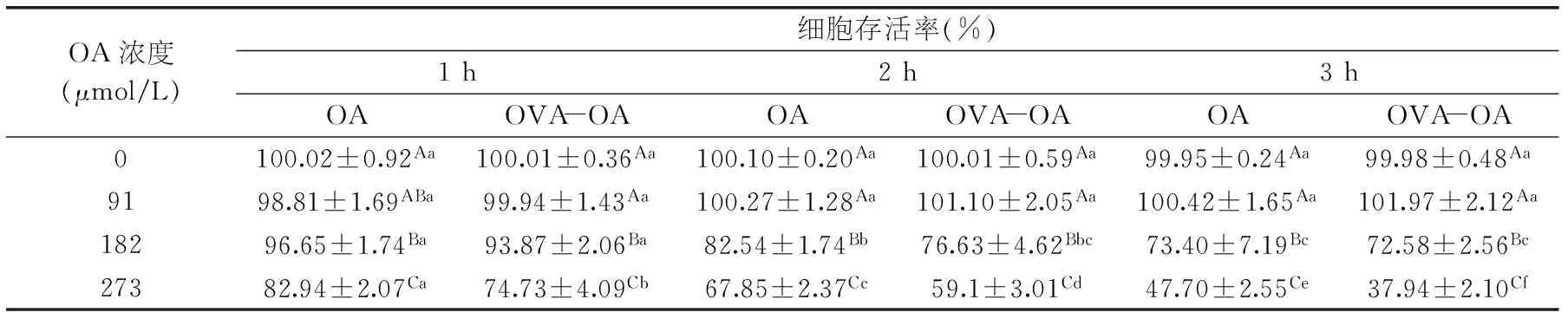
表2 在不同濃度和不同作用時間時OVA-OA及OA對Caco-2細胞的生長抑制作用
LDH釋放率(%)=ΔAbs340 nm,sample/ΔAbs340 nm,control×100
式(2)
式中ΔAbs340 nm為反應體系每分鐘Abs340 nm的減少值。
1.2.4細胞凋亡檢測按照1.2.3中的方法進行種盤和樣品處理,樣品處理結束后棄去樣品,每皿加入8 mL的DMEM繼續培養24 h后,消化,離心(100×g,5 min)收集細胞,用預冷(4 ℃)的PBS洗滌細胞兩次,洗凈胰酶和EDTA。用400 μL Annexin V結合液懸浮細胞,并轉移到流式管中,加入5 μL Annexin V-FITC染色液,輕輕混勻,于2~8 ℃避光孵育15 min;上機檢測前加入10 μL PI染液,輕輕混勻,于2~8 ℃避光條件下孵育5 min,上機檢測[16]。另用未染色的及Annexin V單染的對照組細胞對儀器進行校準。
流式細胞儀采用激發波長488 nm,發射波長530 nm檢測FITC和620 nm檢測PI,FITC的綠色熒光通過FITC通道(FL1)檢測,PI紅色熒光通過PI通道(FL2)檢測。用Cell Quest軟件分析得到不同凋亡期細胞的百分比。
1.2.5細胞周期檢測按照1.2.4中的方法處理細胞至預冷PBS洗凈胰酶和EDTA后,加入300 μL PBS并重懸細胞,緩慢加入700 μL預冷(-20 ℃)的無水乙醇(乙醇終濃度為70%),-20 ℃固定1 h;離心(100×g,5 min)棄上清,用5 mL預冷的PBS洗滌細胞兩次;加0.25 mL PBS重懸細胞,并轉移到流式管中,加入13 μL 10 mg/mL的RNA酶(終濃度0.5 mg/mL),37 ℃避光孵育1 h;加入14 μL 1 mg/mL的PI(終濃度50 μg/mL),4 ℃避光30 min左右后,用流式細胞儀收集細胞(激發波長488 nm,發射波長620 nm),并用Modfit軟件分析得到不同生長周期細胞的百分比[17]。
1.2.6統計學分析所有實驗均重復3次,數據以Mean±SD表示。采用Minitab 17.1.0軟件進行單因素方差分析(analysis of variance,ANOVA),進一步用Turkey多重比較確定各組數據間的顯著性差異,顯著水平為設定為p<0.05。
2 結果與分析
2.1OVA-OA對Caco-2細胞的生長抑制作用
采用不同摩爾比的OA與OVA(OVA 2 mg/mL;OA與OVA的摩爾比為0∶1、2∶1、4∶1、6∶1)制備OVA-OA復合物。首先研究加熱對OVA與OA對Caco-2細胞的細胞生長抑制作用的影響(表1);再同時用OVA-OA和與OVA-OA中相應濃度的OA(0、91、182、273 μmol/L)作用于Caco-2細胞,比較兩者的細胞生長抑制作用(表2)。
由表1可知,未加熱時,OVA對Caco-2細胞沒有生長抑制作用;濃度為91 μmol/L時,OA也沒有生長抑制作用;182 μmol/L時,對細胞有顯著的(p<0.05)生長抑制作用,且273 μmol/L時抑制作用增強。加熱后,OVA、OA對細胞的生長抑制作用均與未加熱時沒有顯著性差異。所以,加熱并不會影響OVA和OA對Caco-2細胞的生長抑制作用。
當OVA-OA中OA的濃度為91 μmol/L時,對Caco-2細胞無生長抑制作用(1~3 h);182 μmol/L時,對細胞有顯著的(p<0.05)生長抑制作用,273 μmol/L時抑制作用增強,且隨著作用時間的延長而增強。在濃度較低(91、182 μmol/L)時,OA與OVA-OA對細胞的生長抑制作用相當;但當OA濃度為273 μmol/L時,其生長抑制作用明顯弱于OVA-OA的;作用時間相同時,OA的細胞生長抑制率比OVA-OA低8%~10%。
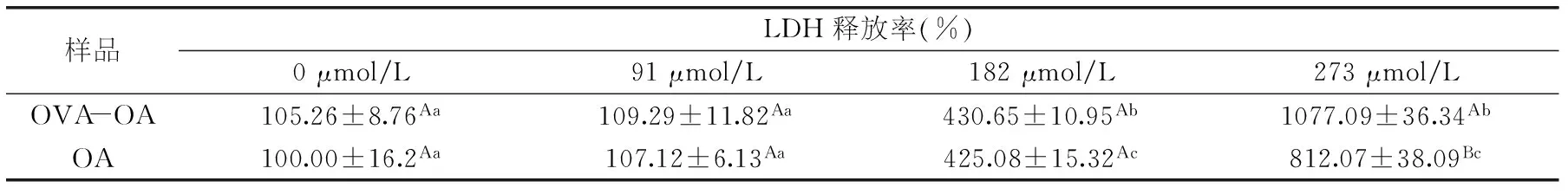
表3 不同濃度OVA-OA及OA對Caco-2細胞的LDH活性和釋放率的影響
研究發現α-LA-OA對肺、腎、結腸、膀胱、前列腺[1]、肝臟、乳腺[18]、喉[19]等來源的腫瘤細胞均有生長抑制作用。本研究顯示,OVA-OA對人結腸癌細胞(Caco-2)也有HAMLET樣生長抑制作用。Zhu等[20]研究OA對人類神經母瘤SH-SY5Y細胞的生長抑制作用,發現OA致SH-SY5Y細胞生長抑制與時間和濃度呈依賴性,當OA的濃度為200 μmol/L,作用3 h時,細胞的抑制率約為35%;作用6 h時,細胞的抑制率約為55%。Lima等[21]也發現OA對淋巴瘤細胞(Jurkat)和惡性淋巴瘤細胞(Raji)有生長抑制作用。在本實驗中,OA對Caco-2細胞的生長抑制也呈時間和濃度依賴,在273 μmol/L時作用3 h可以抑制細胞生長率至47.7%。
Brinkmann等[12]研究α-LA-OA對HL-60細胞的生長抑制作用,發現OA(濃度為0~100 μmol/L)與α-LA-OA對細胞表現出相似的細胞生長抑制作用,且α-LA-OA的生長抑制作用略高于OA。Permyakov等[19]研究發現α-LA-OA對人喉頭癌細胞(HEp-2)的半抑制率濃度為(IC50值)240 μmol/L(用OA的濃度表示),而OA單獨存在時IC50值為300 μmol/L,α-LA單獨存在時沒有細胞生長抑制作用。所以α-LA-OA中發揮細胞生長抑制作用的成分主要是OA,而α-LA可能主要起增加OA溶解度與運輸OA的作用。在本實驗中OVA本身沒有細胞生長抑制作用,OA具有細胞生長抑制作用,且濃度相同時OVA-OA生長抑制作用強于OA,所以在OVA-OA中發揮細胞生長抑制作用的成分是OA,OVA可能主要通過增加OA溶解度或運輸OA來提高OA自身的細胞生長抑制作用。
2.2OVA-OA對Caco-2細胞膜的破壞作用
根據表2可知樣品作用時間為3 h時,OVA-OA及OA對細胞的生長抑制作用最強,所以選擇作用3 h研究其對細胞膜的破壞作用(表3)。
當OVA-OA中的OA濃度為91 μmol/L時,細胞的LDH釋放率沒有顯著(p<0.05)變化;182 μmol/L時,細胞的LDH釋放率顯著增大,增大為對照組的4.5倍;273 μmol/L時LDH的釋放率進一步增大,為對照組的10.8倍。在濃度較低(91和182 μmol/L)時,OA與OVA-OA對細胞LDH釋放率的影響相當;但當OA濃度為273 μmol/L時,OA對細胞LDH的釋放率明顯低于OVA-OA,約低25%。
乳酸脫氫酶(lactate dehydrogenase,LDH)是存在于細胞內,參與糖酵解最后一步即丙酮酸和乳酸相互轉化的一種酶,當細胞膜受損傷時,釋放的LDH會增加,故細胞膜釋放的LDH增加通常反映細胞膜受損傷,膜通透性增大[14]。所以OA濃度為182,273 μmol/L時,OA與OVA-OA對Caco-2細胞的膜造成了損傷,致使LDH釋放,且273 μmol/L時OVA-OA對細胞膜的損傷程度更大。Mossberg等[21]用蛋黃磷脂酰膽堿或大豆卵磷脂組成單膜囊泡模仿熒光分子標記的α-LA-OA與細胞膜的相互作用,發現α-LA-OA能夠結合人造細胞膜,增大膜的流動性,同時會破壞細胞膜的完整性,導致細胞內的一些物質泄漏到細胞外,而未結合OA的α-LA并沒有對細胞膜產生破壞作用,與本研究的結果一致。Mossberg等[21]也研究了α-LA-OA與從肺癌細胞A549中提取的質膜的作用,發現α-LA-OA也可以與其結合,但與人造細胞膜不同的是α-LA-OA在質膜中的分布不均勻,這說明癌細胞的膜中可能有α-LA-OA特定的高親和位點。
2.3OVA-OA對Caco-2細胞凋亡的影響
根據表2可知樣品作用時間為3 h時,OVA-OA及OA對細胞的生長抑制作用最強,所以選擇作用3 h研究OVA-OA和OA對Caco-2細胞凋亡的影響,探討經樣品處理后不同時期凋亡的細胞百分比變化(圖1和表4)。
當OVA-OA復合物中OA的濃度為91 μmol/L時,細胞的凋亡沒有顯著(p<0.05)變化;182 μmol/L時,晚期凋亡和壞死的細胞百分比開始顯著增大,增大了約12%,而早期凋亡的細胞百分比沒有變化;273 μmol/L時晚期凋亡和壞死的細胞百分比進一步增大,增了約24%,早期凋亡細胞百分比也增大了3%左右。當OA濃度為91,182,273 μmol/L時,OA對細胞凋亡的影響均與OVA-OA一致。所以,OVA-OA和OA可以增大晚期凋亡和壞死的細胞百分比,且高劑量時(273 μmol/L)OVA-OA也可以使早期凋亡細胞百分比增多。
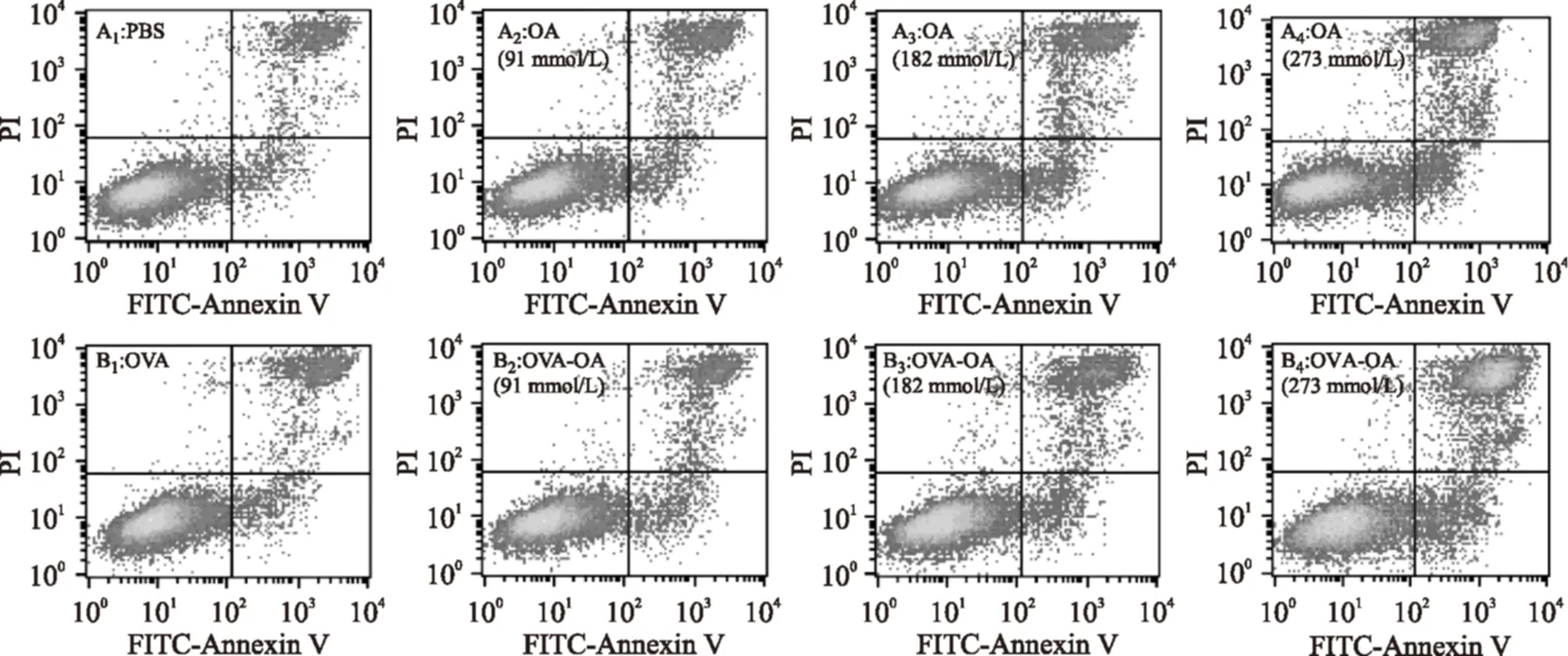
圖1 不同濃度OVA-OA及OA對Caco-2細胞凋亡的影響Fig.1 Cell apoptosis effects of OA and OVA-OA at different concentrations注:A:不同濃度的OA組;B:含不同濃度OA的OVA-OA組。左上象限顯示機械性損傷細胞(damaged),為Annexin V-/PI+;右上象限顯示凋亡晚期或者繼發性壞死細胞(late apoptosis/necrosis),為Annexin V+/PI+;左下象限顯示活細胞(viable),為Annexin V-/PI-;右下象限顯示早期凋亡細胞(early apoptosis),為Annexin V+/PI-。

表4 不同濃度OVA-OA及OA對Caco-2細胞凋亡的影響
注:不同字母小寫字母表示同列不同均數差異顯著(p<0.05);表5同。
OVA-OA和OA可以使晚期凋亡和壞死的細胞數增多,而晚期凋亡和壞死細胞的膜遭到破壞[22],說明OVA-OA和OA可以破壞Caco-2的細胞膜,與LDH實驗的結果一致。Fang等[11]用α-LA-OA作用Caco-2細胞24 h后,細胞的凋亡率約為50%,其中晚期凋亡和壞死的細胞數占39.54%。Brinkmann等[12]研究α-LA-OA作用3 h后對人急性單核細胞白血病細胞(THP-1)凋亡的影響,也得出了相似的結論。所以蛋白質和油酸的復合物可以增大晚期凋亡和壞死的細胞百分比。

表5 不同濃度OVA-OA及OA對Caco-2細胞生長周期的影響
2.4OVA-OA對Caco-2細胞生長周期的影響
根據表2可知,樣品作用時間為3 h時,OVA-OA及OA對細胞的生長抑制作用最強,所以選擇作用3 h研究OVA-OA和OA對Caco-2細胞周期的影響,探討經樣品處理后處于細胞周期不同階段的細胞百分比變化(圖2和表5)。

圖2 不同濃度OVA-OA及OA對Caco-2細胞生長周期的影響Fig.2 Cell cycle distribution effects of OA and OVA-OA at different concentrations注:A:不同濃度的OA組;B:含不同濃度OA的OVA-OA組。
當OVA-OA復合物中OA濃度為91 μmol/L時,細胞周期分布沒有發生變化;而濃度為182,273 μmol/L時,G2期和M期的細胞百分比顯著(p<0.05)降低,降低了10%左右,G0和G1期、S期細胞百分比均有增加的趨勢。當OA濃度為91、182、273 μmol/L時,OA對細胞周期的影響均與OVA-OA一致。所以OVA-OA和OA主要通過減少G2期和M期的細胞百分比,使細胞周期阻滯在G0/G1期和S期來影響細胞周期分布。
細胞周期是指一次有絲分裂結束到下一次有絲分裂的結束,細胞由一個分裂為兩個子細胞的過程,包括休眠期/DNA合成前期(G0/G1期)、DNA合成期(S期)、DNA合成后期(G2期)和細胞分裂期(M期)[23-24]。OVA-OA及OA處理Caco-2細胞后,可以顯著減少G2期和M期的細胞百分比,說明經OVA-OA及OA的處理會影響細胞DNA的合成,使細胞難以進入DNA合成后期和分裂期,進而影響細胞分裂。方冰[16]研究α-LA-OA對HT-29細胞周期的影響,發現α-LA-OA會影響染色體的分裂,使細胞阻滯在G2/M期,與本實驗結果不同。
3 結論
卵白蛋白和油酸復合物(OVA-OA)對人結腸癌Caco-2細胞具有HAMLET樣的生長抑制作用,其中OA起主要作用。OVA-OA中的OA劑量越大、作用時間越長,則細胞生長抑制作用越強。OVA-OA的癌細胞生長抑制作用是通過損傷細胞膜導致LDH釋放率增大;使細胞周期阻滯在G0/G1期和S期從而阻滯細胞生長周期;使晚期凋亡和壞死的細胞百分比增多來實現的。
[1]Hakansson A,Zhivotovsky B,Orrenius S,et al. Apoptosis induced by a human milk protein[J]. Proceedings of the National Academy of Science,1995,92(17):8064-8068.
[2]Svensson M,Hakansson A,Mossberg AK,et al. Conversion ofα-lactalbumin to a protein inducing apoptosis[J]. Proceedings of the National Academy of Science of the United States of America,2000,97:4221-4226.
[3]Tolin S,Franceschi GD,Spolaore B,et al. The oleic acid complexes of proteolytic fragments ofα-lactalbumin display apoptotic activity[J]. The FEBS Journal,2010,277:163-173.
[4]Wilhelm K,Darinskas A,Noppe W,et al. Protein oligomerization induce by oleic acid at the solid-liquid interface-equine lysozyme cytotoxic complexes[J]. The FEBS Journal,2009,276:3875-3989.
[5]Liskova K,Auty MAE,Chaurin V,et al. Cytotoxic complexes of sodium oleate with beta-lactoglobulin[J]. European Journal of Lipid Science and Technology,2011,113(10):1207-1218.
[6]Fang B,Zhang M,Tian M,et al. Bovine lactoferrin binds oleic acid to form an anti-tumor complex similar to HAMLET[J]. Biochimica et Biophysica Acta,2014,1841:535-543.
[7]Huntington JA,Stein PE. Structure and properties of ovalbumin[J]. Journal of Chromatography B,2001,756(1-2):189-198.
[8]Kaspersen JD,Pedersen JN,Hansted JG,et al. Generic structure of cytotoxic liprotides:nano-sized complexes with oleic acid cores and shells of disordered proteins[J]. Chembiochem,2014,15(18):2693-2702.
[9]Sponton OE,Perez AA,Carrara CR,et al. Linoleic acid binding properties of ovalbumin nanoparticles[J]. Colloids and Surfaces B:Biointerfaces,2015,128:219-226.
[10]Kamijima T,Ohmura A,Sato T,et al. Heat-treatment method for producing fatty acid-bound alpha-lactalbumin that induces tumor cell death[J]. Biochemical and Biophysical Research Communications,2008,376:211-214.
[11]Fang B,Zhang M,Jiang L,et al. Influence of pH on the structure and oleic acid binding ability of bovineα-lactalbumin[J]. Protein Journal,2012,31:564-572.
[12]Brinkmann CR,Heegaard CW,Petersen TE,et al. The toxicity of alpha-lactalbumin made lethal to tumor cells is highly dependent on oleic acid and induces killing in cancer cell lines and noncancer-derived cells[J]. FEBS Journal,2011(278):1955-1967.
[13]Delgado Y,Cruz MM,Figueroa CM,et al. The cytotoxicity of BAMLET complexes is due to oleic acid and independent of theα-lactalbumin component[J]. FEBS Open Bio,2015,5:397-404.
[14]Popovich DG,Kitts DD. Structure-function relationship exists for ginsenosides in reducing cell proliferation and induce apoptosis in the human leukemia(THP-1)cell line[J]. Archives of Biochemistry and Biophysics,2002,406(1):1-8.
[15]Sargent J,Taylor C. Appraisal of the MTT assay as a rapidtest of chemosensitivity in acute myeloid leukaemia[J]. British Journal of Cancer,1989,60(2):206-210.
[16]Alday E,Valencia D,Carreno AL,et al. Apoptotic induction by pinobanksin and some of its ester derivatives from Sonoran propolis in a B-cell lymphoma cell line[J]. Chemico-Biological Interactions,2015,242:35-44.
[17]張永奎,李建民,王東隸,等. 大蒜素對體外人骨肉瘤細胞周期和凋亡的影響[J]. 腫瘤,2015,55(5):214-222.
[18]方冰. 乳白蛋白-油酸復合物的制備及其抗腫瘤機理研究[D]. 北京:中國農業大學,2013,53.
[19]Permyakovz SE,Knyazeva EL,Khasanova LM,et al. Oleic acid is a key cytotoxic component of HAMLET-like complexes[J]. Biological Chemistry,2012,393(1-2):85-92.
[20]Zhu Y,Schwarz S,Ahlemeyer B,et al. Oleic acid cause apoptosis and dephosphorylates bad[J]. Neurochemistry International,2005,46:127-135.
[21]Mossberg AK,Puchades M,Halskau O,et al. HAMLET interacts with lipid membranesand perturbs their structure and integrity[J]. Plos One,2010,5(2):e9384.
[22]Span LFR,Pennings AHM,Vierwinden G,et al. The dynamic process of apoptosis analyzed by flow cytometry using Annexin-V/propidium iodide and a modified in situ end labeling technique[J]. Cytometry,2002,47(1):24-31.
[23]高燕,林莉萍,丁健. 細胞周期調控的研究進展[J]. 生命科學,2005,17(4):318-322.
[24]Chiu A,Shi XL,Lee WKP,et al. Review of chromium(VI)apoptosis,cell-cycle-arrest,and carcinogenesis[J]. Journal of Environmental Science and Health,2010,28(3):188-230.
HAMLET-like cell-growth-inhibitory effect of ovalbumin-oleic acid complex on colon cancer cell
XING Xiao-juan,LIU Jian-lei,JING Hao*
(College of Food Science & Nutritional Engineering,China Agriculture University,Beijing 100083,China)
A complex ofα-lactalbumin and oleic acid has previously been shown to have growth-inhibitory effect on cancer cells(Human alpha-lactalbumin made lethal to tumor cells,HAMLET). Ovalbumin and oleic acid complex was prepared and its cell growth inhibition effect,along with contribution of oleic acid,were all evaluated on Caco-2 cell by using cell-growth inhibition,cellular membrane integrity,cell cycle and apoptosis assays. The result showed that OVA-OA significantly(p<0.05)inhibited cell growth at 182 μmol/L(OA),and the inhibitory effect was enhanced as the concentration was increased to 273 μmol/L. The inhibitory effect was also increased as incubation time was extended in a time-dependent manner. OA at 182 μmol/L also exhibited remarkable cell-growth-inhibitory effect in a similar pattern to OVA-OA,while its inhibitory effect was lower than that of OVA-OA at the same OA concentration of 273 μmol/L. Both OVA-OA and OA could increase LDH release rate,lead to cycle arrested at G0/G1and S phases,and induce apoptosis. In conclusion,the complex of OVA and OA has HAMLET-like cell-growth-inhibitory effect,which could be mainly contributed to OA. The mechanisms were associated with damage of cellular membrane integrity,arrest of cell cycle and induction of cell apoptosis.
ovalbumin;oleic acid;cell-growth inhibition;lactate dehydrogenase;cell apoptosis;cell cycle
2016-03-30
邢效娟(1992-),女,碩士研究生,研究方向:蛋白質與小分子相互作用,E-mail:x2010314024@163.com。
景浩(1957-),男,博士,教授,研究方向:分子營養與食品安全,E-mail:haojing@cau.edu.cn。
“十二五”農村領域國家科技計劃課題子課題(2012BAD28B08)。
TS201.4
A
1002-0306(2016)18-0137-07
10.13386/j.issn1002-0306.2016.18.017
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
