沉水植物在富營養化淺水湖泊修復中的生態機理
2016-11-02 09:57:31楊鳳娟蔣任飛饒偉民代曉炫肖許沐謝海旗
安徽農業科學 2016年26期
關鍵詞:生態
楊鳳娟, 蔣任飛, 饒偉民, 代曉炫, 肖許沐, 謝海旗*
(1.中水珠江規劃勘測設計有限公司,廣東廣州 510610;2.水利部珠江水利委員會水生態工程中心,廣東廣州 510610)
?
沉水植物在富營養化淺水湖泊修復中的生態機理
楊鳳娟1,2, 蔣任飛1,2, 饒偉民1,2, 代曉炫1,2, 肖許沐1,2, 謝海旗1,2*
(1.中水珠江規劃勘測設計有限公司,廣東廣州 510610;2.水利部珠江水利委員會水生態工程中心,廣東廣州 510610)
通過查閱近年來淺水湖泊沉水植物修復相關文獻報道,總結了沉水植物在富營養化湖泊治理中的生態機理,分析了影響沉水植物恢復的主要因子(藻類影響、魚類攝食及擾動等),綜合探討了沉水植物在富營養化淺水湖泊修復中的重要作用,為科學合理地開展富營養化淺水湖泊修復實踐活動提供理論指導與經驗借鑒。
沉水植物;富營養化湖泊;生態修復;生態機理
水體富營養化不僅影響水資源的利用、漁業的發展,而且會刺激有害藻類的暴發,使水域喪失生態功能和價值[1]。同時,水體富營養化還給人們的生活和經濟帶來極大的危害和損失[2]。相關調查表明,我國湖泊、水庫富營養化的比例分別為77.00%、30.80%[3-4],湖泊富營養化成為我國最嚴重的水環境問題之一。
沉水植物作為初級生產者,在水生態系統中起著重要作用,重建沉水植物群落結構被認為是修復富營養化水體的有效手段[5-6]。我國富營養化淺水湖泊成功修復的案例以惠州西湖和太湖五里湖最為典型。通過恢復沉水植物群落結構,惠州西湖子湖——南湖從以浮游植物為主導初級生產力的“藻型湖泊”轉變為以沉水植物為主要初級生產力的“草型湖泊”,形成了穩定的食物網結構,透明度由30 cm提高至150 cm,水體總磷、總氮含量分別降低54.50%和52.70%[7];太湖五里湖水體從藻類占優勢的濁水態向水生植物占優勢的清水態轉變,透明度由39 cm提高至70 cm,水體總磷、總氮含量分別降低23.80%和20.70%[8]。同時,沉水植物群落也受到諸多因素的影響,如藻類遮光、風浪擾動、營養鹽負荷及魚類攝食和擾動。因此,研究沉水植物在淺水湖泊修復過程中的生態機理及影響沉水植物群落結構重建的相關環境因子,對富營養化淺水湖泊的治理有著重要的理論價值和實踐指導意義。筆者綜述了沉水植物在富營養化湖泊治理中的生態機理,以期為科學合理地開展富營養化淺水湖泊修復實踐活動提供理論依據。
1 沉水植物對水質改善的作用機理
1.1對氮、磷營養鹽的控制氮、磷是植物生長的必需營養元素。長勢良好的水生植物可使水體中總氮、總磷含量明顯降低[9]。沉水植物除了通過根、葉吸收水體中的氮、磷及控制水體中營養鹽濃度外[10],還可在光合作用過程中產生氧氣擴散到根際并進入沉積物,影響沉積物氮、磷的循環[11-12]。相關研究表明,沉水植物覆蓋的區域中,氧氣可以滲透至沉積物表層20 mm處,而在無沉水植物覆蓋的區域,氧氣的滲透深度僅4 mm[13];沉積物中的Fe2+在沉水植物根際氧化作用下形成Fe3+,并與PO43--P形成鐵-磷復合體,抑制磷從沉積物釋放到水體中[14]。同時,沉水植物通過促進沉積物-水界面的反硝化作用,還原硝酸鹽,釋放出分子態氮(N2)或一氧化二氮(N2O),減少水體中的氮營養鹽。雷婷文等[15]研究表明,苦草(VallisneriaL.)、黑藻(HydrillaverticillataL.)、金魚藻(Ceratophyllumdemersum)、伊樂藻(Elodeanuttalli)、狐尾藻(MyriophyllumL.)、菹草(PotamogetoncrispusL.)均能有效去除水體中的氮、磷等營養鹽含量,其中以苦草效果最好。
Gao等[16]研究發現,金魚藻、伊樂藻、眼子菜(PotamogetonL.)、狐尾藻、苦草等沉水植物通過葉片吸收等方式,對水體中總磷具有較好的去除效果,尤其在春季和秋季,金魚藻對水體中總磷的去除率高達92.00%,且對總溶解性磷的去除率也高達90.93%。在太湖梅梁灣的生態修復工程區中,沉水植物的根系吸收作用和化學反應促進作用使沉積物總氮、總磷營養鹽分別由修復前(2003年)的7 043、1 370 mg/kg降低至2 929、352 mg/kg。
1.2提高水體透明度沉積物再懸浮會直接影響水體的透明度,而沉水植物可以減緩水流速度和風力,并且通過根部固定沉積物,抑制再懸浮。同時,沉水植物還可作為懸浮物的捕獲器,促進沉降,進而提高水體透明度[10]。Horppila等[17]進行為期83 d的研究發現,在沉水植物金魚藻和鈍葉眼子菜(Potamogetonobtusifolius)生長的區域,表層沉積物再懸浮量為793 g (DW)/m2,而在無沉水植物生長的區域,表層沉積物再懸浮量為1 701 g(DW)/m2。De等[18]對12個已修復的富營養城市人工湖進行了研究,發現沉水植物覆蓋率大于30%的湖泊能夠長期維持清水態,而沉水植物覆蓋率小于30%的湖泊則再度發展為濁水態湖泊。Horppila等[17]研究發現,在沉水植物生長的區域,通過沉積物再懸浮帶入水體中的磷元素含量為11.8 mg/(m2·d),無沉水植物生長區域的磷元素含量為24.5 mg/(m2·d)。由此可知,在缺乏沉水植物的富營養化湖泊中,沉積物再懸浮會導致水體磷營養鹽濃度增加。
2 沉水植物對藻類的控制機理
2.1營養鹽競爭沉水植物可以通過吸收氮、磷營養鹽、無機碳等競爭限制藻類的生長。離子態氮(NO3-、NH4+)、磷(PO43-)是初級生產者直接吸收利用的營養形態。不論沉水植物是否存在,藻類的生物量與湖水中氮、磷的含量呈正相關[19]。但藻類生命周期短,對水體中氮、磷營養鹽固定能力弱[14],而沉水植物具有過量吸收營養物質的特性,個體大,生活周期長,體內積累的氮、磷多,儲存較藻類穩定,能有效降低氮、磷循環速度、控制藻類的暴發[20]。
對于根系不發達的沉水植物如黑藻、伊樂藻、金魚藻等,主要依靠自身葉片吸收水體中的營養鹽,如NO3-、PO43-、CO2和HCO3-[21],進而與浮游植物形成競爭關系。研究表明,大量沉水植物可以導致水體中磷含量急劇下降,造成浮游植物缺乏營養鹽而難以在夏天形成水華[22]。對于根系發達的沉水植物如眼子菜、苦草、狐尾藻等,可通過根吸收和促進沉積物中化學反應等方式對內源污染進行控制,削減浮游植物營養鹽來源。2.2化感作用沉水植物分泌的化感物質可抑制水體中浮游植物的生長[23-24],減弱浮游植物或附著藻的遮陰效應,促進自身生長。化感物質可使藻類細胞的生理作用產生變化,如影響抗氧化酶活性、堿性磷酸酶活性,促進丙二醛等有毒物質積累,從而引起藻類生物量的減少。通過化感物質抑制藻類生長的沉水植物至少有37種[25],其中以狐尾藻、金魚藻、伊樂藻、茨藻、輪藻等的化感作用較明顯。
不同沉水植物分泌的化感物質對藻類的作用不同,從而影響浮游植物種類組成。Hilt等[26-27]研究發現,狐尾藻通過分泌化感物質抑制堿性磷酸酶活性,減少葉片表面磷的釋放量,降低附著藻的磷元素來源,限制附著藻的生長;水蘊草[Elodeadensa(Planch.)Casp.)和伊樂藻可以通過化感作用抑制附著藍藻的生長,進而限制附著藻生物膜的形成,促進葉片表面對光照和營養鹽的吸收[28]。
2.3其他作用浮游植物不具有主動運動能力,通常需要借助風浪、水流等外力作用,而沉水植物可以減弱風浪對水體的擾動,使浮游植物失去漂浮的助力,加速其沉降。此外,沉水植物還可以影響浮游植物對光照的吸收,限制其群落的發展。
3 沉水植物對浮游動物的影響
沉水植物群落可為浮游動物提供主要的棲居地和避難場所[29]。在淺水湖泊中,水平遷移(DielHorizontalMigration,DHM)是浮游動物常見的行為。浮游動物遷移至沿岸帶或沉水植物茂盛的水域中,躲避魚類等的捕食,維持種群數量、群落結構穩定發展,加強對浮游植物個體、種群甚至群落結構的控制作用[30-31]。
Canfield等[32]研究發現,水體中的浮游甲殼類動物生物量與沉水植物在水體中的占有體積(PlantVolumeInhabited,PVI)呈顯著正相關(R=0.47, p=0.000 1)。當缺乏沉水植物時,浮游動物的種群密度處于較低水平(<1ind/L),而浮游植物的密度則處于較高水平(38mm3/L);當沉水植物的PVI>15%~20%時,浮游甲殼類動物生物量處于較高水平,且體型較大的個體居多[33]。由此可見,沉水植物群落在一定程度上可保證浮游動物的種群數量,實現對浮游植物的控制,有利于維持湖泊的清水狀態。
4 沉水植物的生態效應機理
不同營養級的上行效應(Bottom-upEffect)和下行效應(Top-downEffect)相結合維持著生態系統群落結構的穩定。沉水植物對無機環境的影響以及與其他生物之間的相互作用(圖1)保證了生態系統中上行效應和下行效應及其他級聯效應的正常發揮[34-36]。
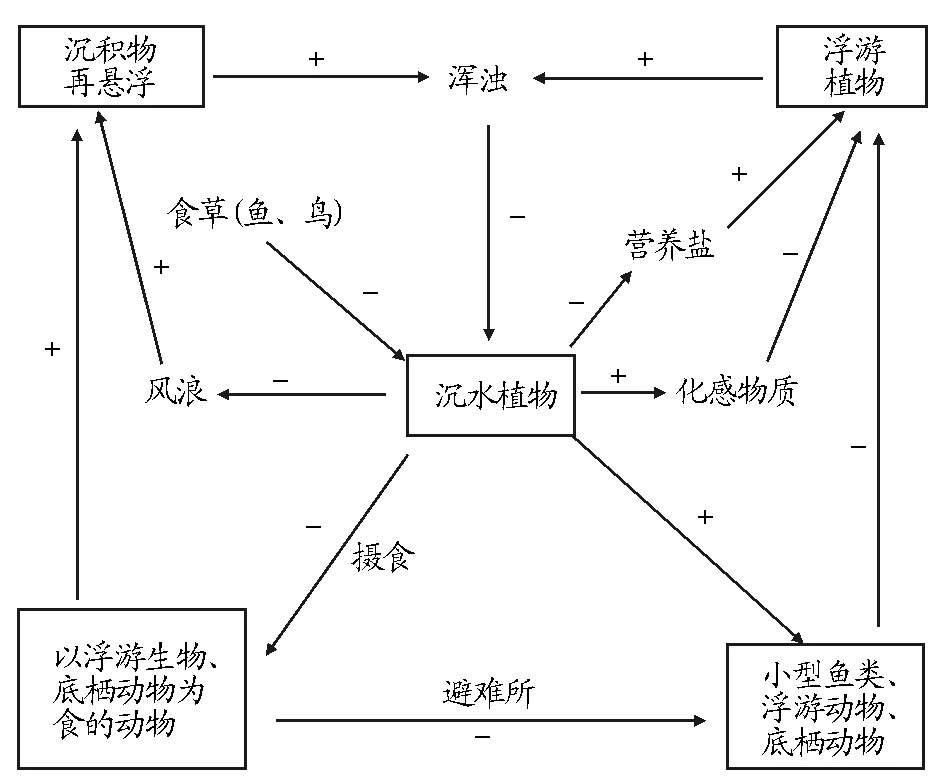
注:“+”表示正反饋;“-”表示負反饋。Note: “+” and “-” stand for positive and negative feedback respectively.圖1 淺水湖泊中沉水植物對湖泊水環境的影響Fig.1 Effects of submerged macrophytes on the water environment of shallow lakes
沉水植物對營養鹽的吸收和鈍化抑制上行效應對浮游植物的促進作用,降低水華暴發的可能性。另外,沉水植物群落結構促進肉食性魚類生長,保證生態系統中下行效應的發揮,即肉食性魚類捕食草食性魚類、浮游生物魚類,優化魚類種群結構,降低沉水植物、浮游動物被大量捕食的壓力,達到控制浮游植物的效果(圖2)[37]。
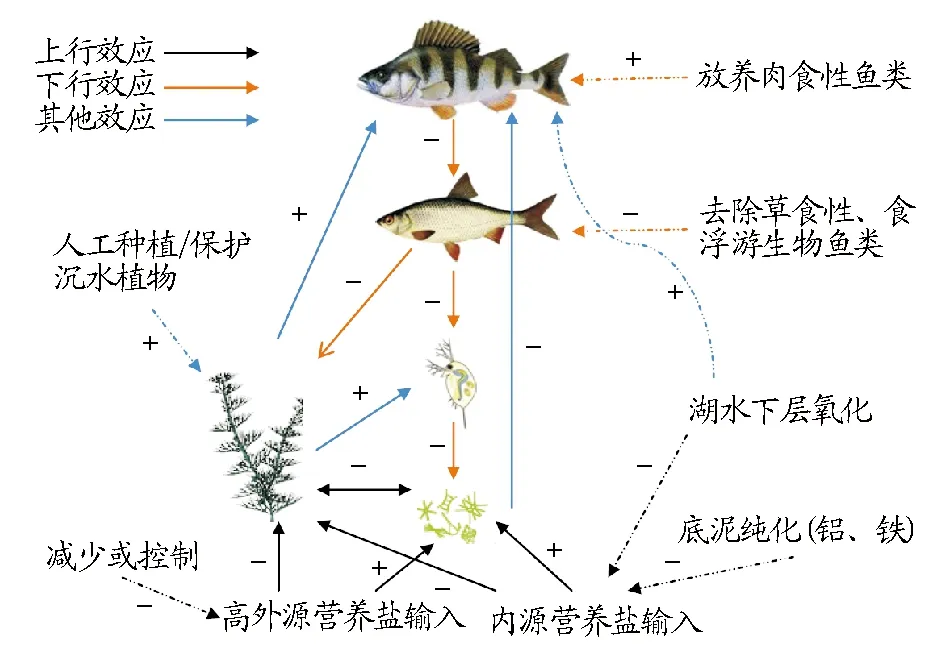
注:“+”表示正反饋;“-”表示負反饋。Note: “+” and “-” stand for positive and negative feedback respectively.圖2 湖泊生態系統上行效應(通過營養鹽)-下行效應(通過攝食)和其他效應模式Fig.2 Conceptual models of bottom-up (via nutrients), top-down (via consumption) and other effects of lake ecosystem
5 影響沉水植物恢復的因子
5.1藻類的限制富營養化湖泊的初級生產力通常以浮游植物為主,呈現濁水態。藻類的遮陰效應、營養鹽和無機鹽的競爭作用會阻礙沉水植物的生長以及在水域中的空間分布[38]。Sand-jensen等[38]研究發現,在富營養湖泊(氮0.93mg/L;磷0.33mg/L),附著藻對沉水植物的遮陰達到86%。當水體中磷濃度大于0.05mg/L時,藻類大量生長形成水華[18];藻類釋放的有色溶解有機顆粒物(ParticulateOrganicMatter,POM)增加光散射,導致光衰減(藍光,波長476~495nm)也隨之增加,從而影響沉水植物的光吸收效率[39],引起水體底層缺氧及植物根系腐爛等現象發生。水華暴發形成氣泡浮渣也會增加光的衰減,對沉水植物造成影響。藻類形成的附著生物膜不僅影響沉水植物葉片表面的光吸收,還阻礙葉片對無機碳的吸收,影響碳酸鹽在葉片表面的擴散[40]。
針對藻類引起的不良影響,通常可以采用生物控制與物理通氣、曝氣等方法[41-42]。物理通氣等方法可破壞藻類的垂直分布[41],增加水體CO2,增大葉片與CO2的接觸面積,解除CO2限制。浮游動物也可實現對藻類的控制[43-44]。在發生藻類水華時,可投放、接種浮游甲殼類動物如長刺溞(Daphnia longispiana)、大型溞(Daphnia magna)和方形網紋溞(Gerzoda daphnia gamaranaura)等進行控制[45-46]。
5.2魚類的影響草食性、雜食性魚類及底層魚類會對沉水植物造成影響[47]。由于草食性魚類食量大,每天攝食沉水植物的量甚至超過魚自身的重量[48],當攝食強度超過沉水植物的再生能力時,會導致沉水植物大規模減少甚至消失。而控制草食性魚類的生長,會促進沉水植物的生長[49-50]。Qiu等[51]研究發現,去除武漢東湖草食性魚類之后,微齒眼子菜(Potamogeton maackianus)自行恢復至6 000g(WW)/m2的生物量。魚類對底泥的擾動及排泄作用導致底泥中營養鹽釋放至水體中,刺激藻類生長,反過來抑制沉水植物生長。Wright等[52]對英國2個湖泊進行研究,發現當去除湖泊中的魚類(396kg/hm2),沉水植物覆蓋度由1%上升到93%,生物量由1.0g/m2上升到46.7g/m2;當再度引進魚類時,沉水植物生物量又由46.7g/m2下降至4.8g/m2。由此可見,魚類是沉水植物生長的重要限制因子。
6 建議
富營養化淺水湖泊修復過程中,控制營養鹽和構建健康的生態系統是主要目標。了解沉水植物在富營養化淺水湖泊治理中的生態機理,恢復沉水植物群落是富營養化淺水湖泊修復的重要方向。但是作為維持湖泊清水態的關鍵因子,沉水植物受到諸多因素的影響。因此,現階段通過改善基礎環境恢復沉水植物是實現富營養化淺水湖泊生態恢復的有效途徑。
在采用“恢復沉水植物群落”手段進行富營養化湖泊修復時,筆者提出了以下幾點建議:①針對不同地區(如不同氣候帶等)的修復對象,結合客觀實際,優先選用土著種、化感作用強、根際固定作用強的沉水植物用于控制藻類及沉積物內源污染;②在沉水植物群落構建初期,采取相應措施減少相關因素對沉水植物生長的影響,如通過添加絮凝劑等透明度提升工程、浮游動物控藻手段等,保證沉水植物在修復初期快速生長;③為促進沉水植物群落結構的快速形成,建議在初期去除水體中草食性魚類和底層魚類,以減少魚類對沉水植物的攝食及對沉積物的擾動;④當系統趨于穩定時,適當放養肉食性魚類(如鱖魚等),結合生物操縱原理,進行水生態系統構建,使生態系統中的上行效應和下行效應得以發揮,從而進一步加強系統的穩定性。
[1]SMITHVH,TILMANGD,NEKOLAJC.Eutrophication:Impactsofexcessnutrientinputsonfreshwater,marine,andterrestrialecosystems[J].Environmentalpollution, 1999, 100(1): 179-196.
[2]GUOL.DoingbattlewiththegreenmonsterofTaihuLake[J].Science, 2007, 317(5842): 1166.
[3] 金相燦,劉樹坤,章宗涉.中國湖泊環境[M].北京:海洋出版社,1995.
[4]LEC,ZHAY,LIY,etal.EutrophicationoflakewatersinChina:Cost,causes,andcontrol[J].Environmentalmanagement, 2010, 45(4): 662-668.
[5] 曹昀, 胡紅, 時強, 等.沉水植物恢復的透明度條件研究[J].安徽農業科學,2012, 40(3): 1710-1711.
[6]CHENGX,LIS,PUP.Ecologicalrestorationofurbaneutrophiclakes:AcasestudyonthephysicalandecologicalengineeringinLakeMochou,Nanjing[J].Journaloflakesciences, 2006, 18(3): 218-224.
[7] 劉從玉, 劉平平, 劉正文, 等.沉水植物在生態修復和水質改善中的作用[J].安徽農業科學, 2008, 36(7): 2908-2910.
[8] 陳開寧, 包先明, 史龍新, 等.太湖五里湖生態重建示范工程:大型圍隔試驗[J].湖泊科學,2006, 18(2): 139-149.
[9] 馬劍敏, 嚴國安, 任南, 等.東湖大型圍隔及圍欄內植物群落和水質的變化[J] .植物資源與環境, 1996, 5(3):35- 40.
[10]BARKOJW,JAMESWF.Effectsofsubmergedaquaticmacrophytesonnutrientdynamics,sedimentation,andresuspension[M]//Thestructuringroleofsubmergedmacrophytesinlakes.NewYork:Springer,1998: 197-214.
[11]OREMLANDRS,TAYLORBF.DiurnalfluctuationsofO2,N2,andCH4intherhizosphereofThalassiatestudinum[J].Limnologyandoceanography, 1977, 22(3): 566-570.
[12]SMITHRD,DENNISONWC,ALBERTERS.Roleofseagrassphotosynthesisinrootaerobicprocesses[J].Plantphysiology, 1984, 74(4): 1055-1058.
[13]WANGY,LIZ,ZHOUL,etal.Effectsofmacrophyte-associatednitrogencyclingbacteriaondenitrificationinthesedimentsoftheeutrophicGonghuBay,TaihuLake[J].Hydrobiologia, 2013, 700(1): 329-341.
[14]JAYNESML,CARPENTERSR.Effectsofvascularandnonvascularmacrophytesonsedimentredoxandsolutedynamics[J].Ecology, 1986: 875-882.
[15] 雷婷文, 魏小飛, 戴耀良, 等.6種常見沉水植物對水體的凈化作用研究[J].安徽農業科學, 2015, 43(36):160-161,196.
[16]GAOYX,ZHUGW,QINBQ,etal.EffectofEcologicalengineeringonthenutrientcontentofsurfacesedimentsinLakeTaihu,China[J].Ecologicalengineering, 2009, 35(11): 1624-1630.
[17]HORPPILAJ,NURMINENL.EffectsofsubmergedmacrophytesonsedimentresuspensionandinternalphosphorusloadinginLakeHiidenvesi(southernFinland)[J].Waterresearch, 2003, 37(18): 4468-4474.
[18]DEBACKERS,TEISSIERS,TRIESTL.Stabilizingtheclear-waterstateineutrophicpondsafterbiomanipulation:Submergedvegetationversusfishrecolonization[J].Hydrobiologia, 2012, 689(1): 161-176.
[19] 楊清心.富營養水體中沉水植物與浮游藻類相互競爭的研究[J].湖泊科學,1996(8):17-24.
[20]JEPPESENE,LAURIDSENTL,KAIRESALOT,etal.Impactofsubmergedmacrophytesonfish-zooplanktoninteractionsinlakes[M]//Thestructuringroleofsubmergedmacrophytesinlakes.NewYork:Springer, 1998: 91-114.
[21]VANDENBERGMS,SCHEFFERM,COOPSH,etal.Theroleofcharaceanalgaeinthemanagementofeutrophicshallowlakes[J].Journalofphycology, 1998, 34(5): 750-756.
[22]HILTS,HENSCHKEI,RüCKERJ,etal.Cansubmergedmacrophytesinfluenceturbidityandtrophicstateindeeplakes?Suggestionsfromacasestudy[J].Journalofenvironmentalquality, 2010, 39(2): 725-733.
[23]K?RNERS,NICKLISCHA.Allelopathicgrowthinhibitionofselectedphytoplanktonspeciesbysubmergedmacrophytes1[J].Journalofphycology, 2002, 38(5): 862-871.
[24]MULDERIJG,MOOIJWM,DONKE.AllelopathicgrowthinhibitionandcolonyformationofthegreenalgaScenedesmus obliquusbytheaquaticmacrophyteStratiotes aloides[J].Aquaticecology, 2005, 39(1): 11-21.
[25]MULDERIJG,VANNESEH,VANDONKE.Macrophyte-phytoplanktoninteractions:Therelativeimportanceofallelopathyversusotherfactors[J].Ecologicalmodelling, 2007, 204(1): 85-92.
[26]HILTS,GROSSEM.Canallelopathicallyactivesubmergedmacrophytesstabiliseclear-waterstatesinshallowlakes?[J].Basicandappliedecology, 2008, 9(4): 422-432.
[27]HILTS.Allelopathicinhibitionofepiphytesbysubmergedmacrophytes[J].Aquaticbotany, 2006, 85(3): 252-256.
[28]ERHARDD,GROSSEM.AllelopathicactivityofElodea canadensisandElodea nuttalliiagainstepiphytesandphytoplankton[J].Aquaticbotany, 2006, 85(3): 203-211.
[29]BURKSRL,LODGEDM,JEPPESENE,etal.Dielhorizontalmigrationofzooplankton:Costsandbenefitsofinhabitingthelittoral[J].Freshwaterbiology, 2002, 47(3): 343-365.
[30]BR?NMARKC,HANSSONLA.Chemicalcommunicationinaquaticsystems:Anintroduction[J].Oikos, 2000, 88(1): 103-109.
[31]JEPPESENE,SφNDERGAARDM,SφNDERGAARDM,etal.Cascadingtrophicinteractionsinthelittoralzone:AnenclosureexperimentinshallowLakeStigsholm,Denmark[J].Archivfürhydrobiologie, 2002, 153(4): 533-555.
[32]CANFIELDJRDE,SHIREMANJV,COLLEDE,etal.PredictionofchlorophyllaconcentrationsinFloridalakes:Importanceofaquaticmacrophytes[J].Canadianjournaloffisheriesandaquaticsciences, 1984, 41(3): 497-501.
[33]SCHRIVERPER,BφGESTRANDJ,JEPPESENE,etal.Impactofsubmergedmacrophytesonfish-zooplanlphytoplanktoninteractions:Large-scaleenclosureexperimentsinashalloweutrophiclake[J].Freshwaterbiology, 1995, 33(2): 255-270.
[34]JEPPESENE,JENSENJP,SφNDERGAARDM,etal.Top-downcontrolinfreshwaterlakes:Theroleofnutrientstate,submergedmacrophytesandwaterdepth[M]//ShallowLakes′ 95.Netherlands:Springer,1997: 151-164.
[35]PERROWMR,JOWITTAJD,STANSFIELDJH,etal.Thepracticalimportanceoftheinteractionsbetweenfish,zooplanktonandmacrophytesinshallowlakerestoration[J].Hydrobiologia, 1999, 395: 199-210.
[36]GULATIRD,VANDONKE.LakesintheNetherlands,theirorigin,eutrophicationandrestoration:State-of-the-artreview[M]//Ecologicalrestorationofaquaticandsemi-aquaticecosystemsintheNetherlands(NWEurope).Netherlands:Springer,2002: 73-106.
[37]JEPPESENE,SφNDERGAARDM,LAURIDSEMTL,etal.Biomanipulationasarestorationtooltocombateutrophication:Recentadvancesandfuturechallenges[J].Advancesinecologicalresearch, 2012, 47: 411-488.
[38]SAND-JENSENK,SφNDERGAARDM.Phytoplanktonandepiphytedevelopmentandtheirshadingeffectonsubmergedmacrophytesinlakesofdifferentnutrientstatus[J].InternationaleRevuedergesamtenHydrobiologieundHydrographie, 1981, 66(4): 529-552.
[39]KALFFJ.Limnology:Inlandwaterecosystems[M].NewJersey:PrenticeHall, 2002.
[40]SAND-JENSENKAJ.Effectofepiphytesoneelgrassphotosynthesis[J].Aquaticbotany, 1977, 3: 55-63.
[41] 郭匿春.浮游動物與藻類水華的控制[D].武漢:中國科學院研究生院(水生生物研究所),2007.
[42] 張麗彬,王金鑫,王啟山,等.浮游動物在生物操縱法除藻中的作用研究[J].生態環境,2007,16(6):1648-1653.
[43]CARPENTERSR,KITCHELLJF.Consumercontroloflakeproductivity[J].BioScience, 1988,38:764-769.
[44]JAROMIRS,JANK.Long-termbiomanipulationofRimovReservior(CzechRepubilc)[J].Hdrobiologia, 1997,345:95-108.
[45]POGOZHEVPI,GERASIMOVATN.Theeffectofzooplanktononmicroalgaebloomingandwatereutrophication[J].Waterresources, 2001,7/8: 420-427.
[46] 張喜勤,徐銳賢,許金玉.水溞凈化富營養化湖水試驗研究[J].水資源保護,1998,24(4):32-35.
[47]VANDONKE,OTTEA.Effectsofgrazingbyfishandwaterfowlonthebiomassandspeciescompositionofsubmergedmacrophytes[M]//Managementandecologyoffreshwaterplants.Netherlands:Springer,1996: 285-290.
[48]VENKATESHB,SHETTYHPC.StudiesonthegrowthrateofthegrasscarpCtenopharyngodoni della (Valenciennes)fedontwoaquaticweedsandaterrestrialgrass[J].Aquaculture, 1978, 13(1): 45-53.
[49] 陳洪達.杭州西湖水生植被恢復的途徑與水質凈化問題[J].水生生物學集刊,1984, 8(2): 237-244.
[50] 陳洪達.養魚對武漢東湖生態系統的影響[J].水生生物學報,1989, 13(4): 359-368.
[51]QIUD,WUZ,LIUB,etal.TherestorationofaquaticmacrophytesforimprovingwaterqualityinahypertrophicshallowlakeinHubeiProvince,China[J].Ecologicalengineering, 2001, 18(2): 147-156.
[52]WRIGHTRM,PHILLIPSVE.Changesintheaquaticvegetationoftwogravelpitlakesafterreducingthefishpopulationdensity[J].Aquaticbotany, 1992, 43(1): 43-49.
Ecological Mechanisms of Submerged Macrophytes inthe Restoration of Eutrophic Shallow Lakes
YANG Feng-juan1,2, JIANG Ren-fei1,2, RAO Wei-min1,2, XIE Hai-qi1,2*et al
1.China Water Resources Pearl River Planning Surveying & Designing Co.,Ltd., Guangzhou, Guangdong 510610; 2.Aquatic Ecological Engineering Center,Pearl River Water Resources Commission of the Ministry of Water Resources, Guangzhou, Guangdong 510610)
According to the application of submerged macrophytes in the restoration of eutrophic shallow lakes in recent years, ecological mechanisms of submerged macrophytes in control of eutrophic shallow lakes were summarized, and the main factors influencing the restoration of eutrophic shallow lakes by submerged macrophytes (such as algae, fish feeding, and disturbance) were analyzed.The important roles of submerged macrophytes in the restoration of eutrophic shallow lakes were discussed.The research can provide theoretical guidance and experiencefor the restoration of eutrophic shallow lakes.
Submerged macrophytes; Eutrophic lakes; Ecological restoration; Ecological mechanisms
水利部公益性行業科研專項(201401013)。
楊鳳娟(1984- ),女,山東鄄城人,工程師,碩士,從事生態研究。*通訊作者,教授級高級工程師,從事生態研究及環評工作。
2016-07-06
S 181.3
A
0517-6611(2016)26-0058-04
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
貴茶(2019年3期)2019-12-02 01:46:32
鄉村地理(2018年3期)2018-11-06 06:51:02
茶葉通訊(2017年2期)2017-07-18 11:38:40
河北城市研究(2015年4期)2015-08-23 11:53:06
浙江工商大學學報(2015年6期)2015-03-01 02:56:40
