青花菜CC-NBS-LRR抗病基因BoCNL1的克隆與分析
2016-10-31 03:19:45黃笑梅金建峰朱思眉朱柯柯趙羅鵬
浙江農業學報 2016年2期
關鍵詞:植物
黃笑梅,金建峰,張 雪,朱思眉,朱柯柯,趙羅鵬,蔣 明,*
(1.臺州學院 生命科學學院,浙江 椒江 318000;2.浙江大學 生命科學學院, 浙江 杭州 310058)
?
青花菜CC-NBS-LRR抗病基因BoCNL1的克隆與分析
黃笑梅1,金建峰2,張雪1,朱思眉1,朱柯柯1,趙羅鵬1,蔣明1,*
(1.臺州學院 生命科學學院,浙江 椒江 318000;2.浙江大學 生命科學學院, 浙江 杭州 310058)
根據已知序列設計PCR引物,從青花菜中克隆1個NBS-LRR(核苷酸結合位點-富含亮氨酸重復)抗病基因BoCNL1;在生物信息學分析的基礎上,利用RT-PCR研究該基因在不同器官中的表達模式。測序結果表明,BoCNL1基因的編碼區全長為2 550 bp,編碼849個氨基酸;編碼蛋白具CC(卷曲螺旋)、NBS和LRR結構域;進化分析結果表明,BoCNL1與不結球白菜的關系最近,在進化樹上處于同一分支,與醉蝶花的關系最遠;RT-PCR結果表明,BoCNL1在根、花莖、葉、花蕾、開放的花和嫩角果中均有表達,但表達量低。
青花菜;抗病基因;CC-NBS-LRR;基因克隆
植物在生長過程中通常會遭受真菌、細菌、線蟲、昆蟲和病毒等的侵襲,通過漫長的進化,植物已形成復雜的防衛機制以抵御各種生物脅迫[1]。病害是農業生產蒙受重大損失的主要生物脅迫之一,植物抗病能力取決于病原物無毒基因和寄主抗病基因產物的識別,以及隨后相關防衛途徑的激活,這一過程通常由抗病基因(resistance gene, R基因)控制[2]。R基因在植物中以基因家族的形式存在,根據編碼蛋白的特征,可分為NBS-LRR(nucleotide-binding site plus leucine-rich repeat, 核苷酸結合位點-富含亮氨酸重復)、eLRR-TM(extracellular leucine-rich repeat plus transmembrane receptor, 胞外富含亮氨酸重復跨膜受體)、eLRR-TM-pkinase(extracellular leucine-rich repeat transmembrane protein kinase, 胞外富亮氨酸重復跨膜蛋白激酶)和STK(serine-threorine kinase, 絲氨酸-蘇氨酸激酶)等類型[3]。
NBS-LRR在R基因中所占的比例最大,編碼蛋白的NBS結構域負責ATP的水解及信號的釋放,LRR則充當蛋白質相互作用的平臺和蛋白活化的元件[4]。NBS-LRR參與多種病原菌的防御,在抗病信號轉導和防衛反應激活過程中起著重要作用[5]。目前已從擬南芥(Arabidopsisthaliana)、甘蔗(Saccharumofficinarum)、森林草莓(Fragariavesca)、亞麻(Linumusitatissimum)、水稻(Oryzasativa)、蘋果(Malus×domestica)和番茄(Solanumlycopersicum)等植物中克隆或鑒定到大量NBS-LRR基因[6-12]。將八棱海棠(Malus×robusta)的CC-NBS-LRR基因導入蘋果,后者抗火疫病的能力顯著增加[13];水稻Pi64基因編碼一個新的CC-NBS-LRR蛋白,該基因與水稻葉瘟和穗瘟病抗性相關[14]。青花菜(Brassicaoleraceavar.italica)是一種深受人們喜愛的保健蔬菜,在生產過程中,受多種病菌的危害,造成產量和品質下降,開展抗病分子育種有著重要的意義,而有關NBS-LRR基因的克隆未見報道。本研究從青花菜中克隆一個NBS-LRR基因的基礎上,進行了序列比對和表達分析,為該基因的功能鑒定和分子育種利用奠定基礎。
1 材料與方法
1.1材料
青花菜材料Bo0112由實驗室栽植,該材料生育期為75 d左右,花球緊實,花蕾深綠色,具較強的霜霉病和灰霉病抗性。于花期采集根、葉、花莖、花蕾、開放的花和嫩角果,置于-80 ℃冰箱備用。從NCBI下載7條十字花科植物的同源序列用于比對,它們分別來自不結球白菜(B.rapa)(登錄號: XP_009147850.1)、大白菜(B.rapasubsp.pekinensis)(ACP30592.1)、甘藍型油菜(B.napus)(CDY22783.1)、高山南芥(Arabisalpina)(KFK34203.1)、擬南芥(Arabidopsisthaliana)(AED95001.1)、亞麻薺(Camelinasativa)(XP_010456005.1)和醉蝶花(Tarenayahassleriana)(XP_010554983.1)
1.2RNA的提取和cDNA合成
RNA提取用TRIzol法;cDNA的合成采用TaKaRa公司的試劑盒,第一鏈和第二鏈的合成根據其提供的說明書進行。
1.3BoCNL1基因的克隆
根據NCBI數據庫中的大白菜(登錄號:FJ842840.1)和不結球白菜(B.rapa)(XM_009149572.1)的NBS-LRR基因序列設計PCR引物,分別為CNL1:5′-ATGGGAGGCTGTGTATCACTAGA-3′和CNL2:5′-TTACTCCTGTTCGTGTCTCTGAAAC-3′。反應體系中含1×phusion HF緩沖液,phusion DNA聚合酶0.5 μL(NEB, USA),0.25 μmol·L-1的dNTPs(生工生物工程(上海)股份有限公司),各0.25 μmol·L-1上、下游引物,60 ng葉片cDNA模板,最后加無菌ddH2O至50 μL。PCR程序為:98 ℃預變性30 s;98 ℃變性10 s,57.5 ℃退火20 s,72 ℃延伸35 s,35個循環;72 ℃延伸10 min。
PCR產物經1.2%瓊脂糖凝膠電泳,割取含目的條帶的膠塊,利用碧云天生物技術研究所的DNA凝膠回收試劑盒回收純化,操作根據其提供的說明書進行。采用生工生物工程(上海)股份有限公司的平端DNA片段添dA試劑盒,根據其提供的操作手冊進行回收產物的平末端加A反應。取3 μL加A產物與pGEM-T easy載體(Promega, USA)連接,于42 ℃通過熱激法將連接產物轉入大腸桿菌DH5α感受態細胞,取4個陽性克隆用于測序。
1.4生物信息學分析
等電點和分子量利用在線工具http://web.expasy.org/compute_pi預測;序列比對用ClustalX 1.83軟件;MEGA 3.1用于系統發育樹的構建,建樹方法為鄰接法(neighbor joining method),自舉檢測次數為1 000。
1.5表達分析
根據測序結果,設計RT-PCR引物,分別為CNL3:5′-AGTTCGTGCGCATCTATTGATG-3′和CNL4:5′-TCGATAATCCAAAGGTGTTGAACACA-3′,預期PCR產物大小約為700 bp。反應體系中含1×PCR buffer,0.5 U的TaqDNA聚合酶(北京鼎國昌盛生物技術有限責任公司),0.3 μmol·L-1的dNTPs(生工生物工程(上海)股份有限公司),各0.2 μmol·L-1的CNL4和CNL5引物,60 ng根、葉、花莖、花蕾、開放的花或嫩角果cDNA,最后加無菌ddH2O至20 μL。PCR程序為:94 ℃預變性5 min;94 ℃變性30 s,56.5 ℃退火60 s,72 ℃延伸60 s,35個循環;最后于72 ℃延伸10 min。PCR產物經1.2%瓊脂糖凝膠電泳、拍照,RT-PCR實驗重復3次。以肌動蛋白基因(登錄號: AF044573.1)為內標,引物分別為5′-TCTCGATGGAAGAGCTGGTT-3′和5′-GATCCTTACCG-AGGGAGGTT-3′,PCR程序為:94 ℃預變性5 min;94 ℃變性30 s,55.6 ℃退火45s,72 ℃延伸90 s,共33個循環。
2 結果與分析
2.1BoCNL1基因及其編碼蛋白的特征
以CNL1/CNL2為引物對,從青花菜葉片cDNA中擴增到目的條帶。測序結果表明,BoCNL1基因的編碼區全長為2 550 bp,分別以ATG與TAA為起始和終止密碼子,GC值為39.73%。BoCNL1編碼849個氨基酸,等電點為7.20,分子量大小為96.5 kD。序列分析結果表明,BoCNL1在+26~+63位具一個CC(coiled coil, 螺旋卷曲)結構域;在+158~+438處具一個NBS結構域,另有一個LRR結構域位于+534~+594處(圖1)。
2.2BoCNL1的進化分析
為明確BoCNL1的進化地位,從NCBI數據庫中下載了7條十字花科植物的同源序列,并利用MEGA軟件構建系統發育樹(圖2)。結果表明,8個CNL序列之間的遺傳距離為0.199~0.731,甘藍型油菜與醉蝶花的遺傳距離最大,擬南芥和醉蝶花次之,為0.720,青花菜與不結球白菜之間遺傳距離最小。它們在系統發育樹上可分為3組,同為蕓薹屬的青花菜、大白菜、不結球白菜和甘藍型油菜聚為一組(Ⅰ);擬南芥、高山南芥和亞麻薺聚為一組(Ⅱ);而醉蝶花單獨處于一個分支(Ⅲ)。
2.3BoCNL1基因的表達分析
為研究BoCNL1在不同器官中的表達模式,分別以等量的根、葉、花莖、花蕾、開放的花或嫩角果cDNA為模板進行PCR擴增。RT-PCR結果表明,BoCNL1在各個部位均有表達,但表達量均處于較低的水平(圖3)。
3 討論
植物抗病基因以超家族的形式存在,大約有數百甚至數千個分布于基因組中,其中以NBS-LRR類型的R基因最為豐富,它們以基因簇的形式分布在染色體上[15]。在擬南芥(哥倫比亞生態型)中有207個R基因,其中149個為NBS-LRR基因[6];從亞麻基因組鑒定得到147個NBS-LRR,其中CC-NBS-LRR類型49個,TIR-NBS-LRR類型98個[9];番茄有252個NBS-LRR,分布于12條染色體上[12];蘋果和大豆(Glycinemax)中的NBS-LRR基因數量十分龐大,分別有505和319個[16-17]。最近,Zhang等[18]分別從琴葉擬南芥(A.lyrata)、不結球白菜(B.rapa)、薺菜(Capsellarubella)和鹽芥(Thellungiellasalsuginea)基因組中鑒定出198,204,127和88個NBS-LRR基因。本研究從青花菜中克隆到一個NBS-LRR基因,該基因的編碼區全長為2 550 bp,編碼849個氨基酸。
根據編碼蛋白N端的結構特征,NBS-LRR可分為TIR-NBS-LRR(TNL)和CC-NBS-LRR(CNL)兩種類型,TIR為果蠅Toll及哺乳動物白細胞介素1受體(DrosophilaToll and mammalian interleukin-1 receptors)的縮寫,CC則為卷曲螺旋(coiled coil)的簡稱[19]。TNL和CNL蛋白在序列和信號傳導途徑上存在顯著差異,但它們均參與病原菌的識別,在監控效應子(effector)誘導蛋白狀態方面起著重要作用[20-21]。本研究中,BoCNL1的N端具一個CC結構域,隨后為NBS和LRR結構域,它是一個典型的CC-NBS-LRR蛋白。
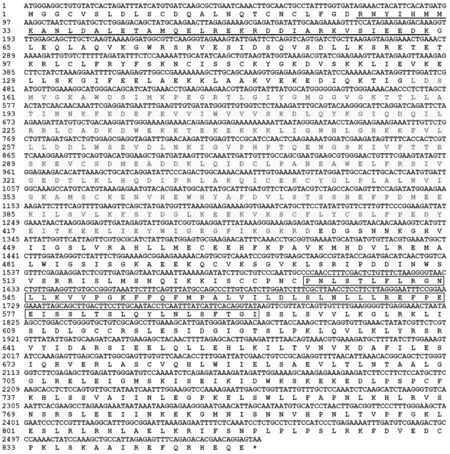
下劃線部分為螺旋卷曲結構域;陰影部分為NBS結構域;方框部分為LRR結構域。圖1 BoCNL1基因及其編碼蛋白序列Fig.1 Gene sequence of BoCNL1 and its deduced protein sequence

圖2 BoCNL1及其同源序列的系統發生樹Fig.2 Phylogenetic tree of BoCNL1 and its homologous sequences
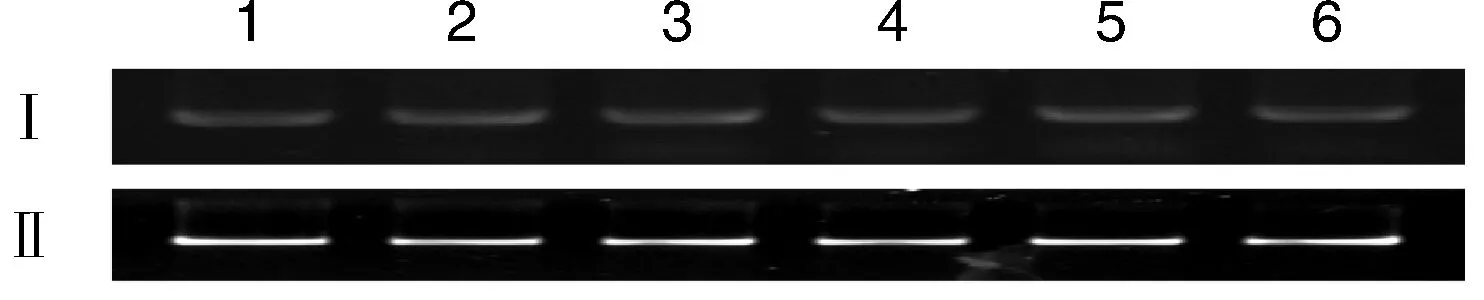
1~6: 根、花莖、葉、花蕾、開放花和嫩角果; Ⅰ: BoCNL1 在不同器官中的表達; Ⅱ: 肌動蛋白內標。圖3 BoCNL1基因的表達分析Fig.3 Expression analysis of BoCNL1
大部分NBS-LRR基因在未感染的健康植株中組成型表達,但表達量較低,僅少部分基因的表達具有組織特異性[21]。在特定病原菌的侵染下,NBS-LRR基因的表達上調,并誘導抗病相關基因的表達[22]。向日葵(Helianthusannuus)自交系QIR8 CC-NBS-LRR基因的表達受霜霉菌(Plasmoparahalstedii)誘導,并激發了一系列信號轉導相關基因的表達[23]。本研究中,BoCNL1在青花菜的根、花莖、葉、花蕾、開放花和嫩角果中均有表達,具有組成性表達的特點,但表達量較低。本課題組將對該基因開展進一步研究,以明確BoCNL1在不同病原菌侵染下的表達模式,解析其在抗病反應中的生物學功能。
[1]DANGL J L, JONES J D. Plant pathogens and integrated defense responses to infection[J].Nature, 2001, 411(6839): 826-833.
[2]XIAO S, ELLWOOD S, CALIS O, et al. Broad-spectrum mildew resistance inArabidopsisthalianamediated by RPW8[J].Science, 2001, 291(5501): 118-120.
[3]王友紅, 張鵬飛, 陳建群. 植物抗病基因及其作用機理[J]. 植物學通報, 2005, 22(1): 92-99.
[4]李峰, 張穎, 樊秀彩, 等. 植物NBS-LRR類抗病基因的研究進展[J]. 分子植物育種(網絡版),2011, 9: 1784-1790.
[6]MEYERS B C, KOZIK A, GRIEGO A, et al. Genome-wide analysis of NBS-LRR-encoding genes inArabidopsis[J].PlantCell, 2003, 15(4): 809-834.
[7]闕友雄, 許莉萍, 林劍偉, 等. 甘蔗NBS-LRR類抗病基因同源序列的分離與鑒定[J]. 作物學報, 2009, 35(4): 631-639.
[8]LI J, ZHANG Q Y, GAO Z H, et al. Genome-wide identification and comparative expression analysis of NBS-LRR-encoding genes uponColletotrichumgloeosporioidesinfection in two ecotypes ofFragariavesca[J].Gene, 2013, 527(1): 215-227.
[9]KALE S M, PARDESHI V C, BARVKAR V T, et al. Genome-wide identification and characterization of nucleotide binding site leucine-rich repeat genes in linseed reveal distinct patterns of gene structure[J].Genome, 2013, 56(2): 91-99.
[10]王世全, 張德春, 李平, 等. 水稻中一個NBS-LRR抗病同源基因家族的克隆和分析[J]. 遺傳學報, 2005, 32(7): 704-711.
[11]宋霄, 柏素花, 戴洪義. 蘋果NBS-LRR1基因的鑒定與表達分析[J]. 園藝學報, 2013, 40(7): 1233-1243.
[12]劉云飛, 萬紅建, 韋艷萍, 等. 番茄NBS-LRR抗病基因家族全基因組分析[J]. 核農學報, 2014, 28(5): 790-799.
[13]BROGGINI G A, W?HNER T, FAHRENTRAPP J, et al. Engineering fire blight resistance into the apple cultivar ‘Gala’ using the FB_MR5 CC-NBS-LRR resistance gene ofMalus×robusta5[J].PlantBiotechnologyJournal, 2014, 12(6): 728-733.
[14]MA J, LEI C, XU X, et al. Pi64, Encoding a novel CC-NBS-LRR protein, confers resistance to leaf and neck blast in rice[J].MolecularPlant-MicrobeInteractions, 2015, 28(5): 558-568.
[15]YOUNG N D. The genetic architecture of resistance[J].CurrentOpinioninPlantBiology, 2000, 3: 285-290.
[16]KANG Y J, KIM K H, SHIM S, et al. Genome-wide mapping of NBS-LRR genes and their association with disease resistance in soybean[J].BMCPlantBiology, 2012, 12: 139.
[17]PERAZZOLLI M, MALACARNE G, BALDO A, et al. Characterization of resistance gene analogues (RGAs) in apple (Malus×domesticaBorkh.) and their evolutionary history of the Rosaceae family[J].PLoSOne, 2014, 9(2): e83844.
[18]ZHANG Y M, SHAO Z Q, WANG Q, et al. Uncovering the dynamic evolution of nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes in Brassicaceae[J].JournalofIntegrativePlantBiology, 2015, doi: 10.1111/jipb.12365.
[19]李春來, 張懷渝. 植物抗病基因同源序列(RGA)研究進展[J]. 分子植物育種, 2004, 2(6): 853-860.
[20]PAN Q, WENDEL J, FLUHR R. Divergent evolution of plant NBS-LRR resistance gene homologues in dicot and cereal genomes[J].JournalofMolecularEvolution, 2000, 50(3): 203-213.
[21]MCHALE L, TAN X, KOEHL P, et al. Plant NBS-LRR proteins: adaptable guards[J].GenomeBiology, 2006, 7(4): 212.
[22]NAVARRO L, ZIPFEL C, ROWLAND O, et al. The transcriptional innate immune response to flg22. Interplay and overlap with Avr gene-dependent defense response and bacterial pathogenesis[J].PlantPhysiology, 2004, 135(2): 1113-1128.
[23]RADWAN O, MOUZEYAR S, NICOLAS P, et al. Induction of a sunflower CC-NBS-LRR resistance gene analogue during incompatible interaction withPlasmoporahalstedii[J].JournalofExperimentalBotany, 2015, 56(412): 567-575.
(責任編輯張韻)
Cloning and characterization of a CC-NBS-LRR disease resistance gene of BoCNL1 from Brassica oleracea var. italica
HUANG Xiao-mei1, JIN Jian-feng2, ZHANG Xue1, ZHU Si-mei1, ZHU Ke-ke1, ZHAO Luo-peng1, JIANG Ming1,*
(1.CollegeofLifeSciences,TaizhouUniversity,Jiaojiang318000,China; 2.CollegeofLifeSciences,ZhejiangUniversity,Hangzhou310058,China)
Primer pairs were designed according to known sequences, and a nucleotide-binding site plus leucine-rich repeat (NBS-LRR) disease resistance gene, designatedBoCNL1, was isolated from broccoli (Brassicaoleraceavar.italic). Bioinformatic analysis were performed, and RT-PCR was used to reveal expression patterns ofBoCNL1 in different organs. Results indicated that the complete coding sequence ofBoCNL1 was 2 550 bp in length, encoding 849 amino acids; and the deduced protein sequence contained coiled coil(CC), NBS, and LRR domains. Phylogenetic analysis results showedBoCNL1 was grouped with the homologous gene inB.rapa, indicating their closest relationship, and the longest genetic distance was observed betweenB.oleraceavar.italicaandTarenayahassleriana. RT-PCR results demonstrated thatBoCNL1 expressed with low levels of transcripts in roots, flower stalks, leaves, flower buds, flowers, as well as young siliques.
Brassicaoleraceavar.italica; disease resistance gene; CC-NBS-LRR; gene cloning
10.3969/j.issn.1004-1524.2016.02.13
2015-07-04
浙江省自然科學基金項目(LY13C150003);浙江省大學生科技創新活動計劃(新苗人才計劃)(2014R428011);浙江省本科院校中青年學科帶頭人學術攀登項目(pd2013420)
黃笑梅(1993—),女,浙江義烏人,本科生,從事植物分子生物學研究。E-mail: 369393248@qq.com
,蔣明,E-mail:jiangming@tzc.edu.cn
S635.3;Q78
A
1004-1524(2016)02-0259-05
黃笑梅,金建峰,張雪,等. 青花菜CC-NBS-LRR抗病基因BoCNL1的克隆與分析[J].浙江農業學報,2016,28(2): 259-263.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
