河南伏牛山區天然紅豆杉種群結構與動態研究
2016-10-28 07:49:30張志錄劉中華陳明輝張巧蓮楊風嶺
水土保持研究 2016年3期
關鍵詞:結構
張志錄, 劉中華, 陳明輝, 張巧蓮, 楊風嶺
(1.平頂山學院, 河南 平頂山 467000; 2.許昌職業技術學院, 河南 許昌 461000)
?
河南伏牛山區天然紅豆杉種群結構與動態研究
張志錄1, 劉中華1, 陳明輝1, 張巧蓮2, 楊風嶺1
(1.平頂山學院, 河南 平頂山 467000; 2.許昌職業技術學院, 河南 許昌 461000)
采用聚集度指標和編制靜態生命表的方法,對伏牛山區紅豆杉種群的空間格局及其演化動態進行了研究,結果顯示:(1) 伏牛山區紅豆杉種群的空間分布格局總體上呈現聚集分布狀態,聚集程度從幼苗階段到老齡階段的發育過程中,呈現先降后升的U型變化趨勢;(2) 不同齡級的紅豆杉樹存活量差別較大,種群存活曲線屬于Deevey-Ⅲ型;(3) 幼苗期和幼樹期的紅豆杉死亡率高,Ⅲ級(3 m高)以后死亡率明顯下降,種群數量趨于穩定。本研究表明,伏牛山自然保護區天然紅豆杉種群呈現明顯的衰退趨勢,嚴酷的環境篩致使紅豆杉幼苗死亡率高可能是該種群衰退的主要原因,但樹木資源缺乏保護而被濫伐或盜伐、生境被破壞等影響也不容忽視。施行嚴格的封育政策并輔以人工繁育措施,是保護和恢復該寶貴資源的當務之急。
瀕危植物; 生命表; 種群結構; 紅豆杉; 時間序列分析
紅豆杉(Taxuschinensis)是第四世紀孑遺植物,是世界瀕危植物,已在地球上生活250萬年,被稱為活化石。中國紅豆杉是中國特有種,屬國家一級保護植物,南北方地區均有分布。在華中地區主要分布于海拔1 000 m以上山地上部未被干擾的生境,在華南和西南地區多分布于1 500~3 000 m的山地落葉闊葉林中[1]。因受花期不遇、幼苗競爭力弱、種子捕食壓力和種子萌發條件等的限制,天然紅豆杉更新困難[2];自發現含有廣譜、強活性的抗癌藥物紫杉醇(Taxol)以來,紅豆杉資源遭到嚴重破壞而數量急劇下降,目前已處于嚴重的瀕危狀態[3],亟待保護與恢復。
種群結構及空間分布格局是種群最基本的特征,既可反映種群內大小不同的個體的分布和配置狀況,也能反映種群的數量動態及其未來發展與演變趨勢,對于種群及群落的更新復壯、生態系統的恢復具有重要意義[4-8]。目前,有關紅豆杉種群的研究報道,多見于南方紅豆杉,集中在種群結構[9-11]、種群分布格局[12-13]和種間關系[14-15]以及天然更新與種子庫[2,16]等方面。這些研究揭示了南方紅豆杉種群結構及其動態的影響因素、形成機制,對于南方紅豆杉瀕危種群的保護發揮了重要作用。然而,紅豆杉的分布存在著地域廣泛性和生境異質性,目前,有關華中地區紅豆杉種群結構與動態的研究比較缺乏。
伏牛山國家級自然保護區處于暖溫帶向亞熱帶的過渡區,是長江、黃河和淮河3大水系的分水嶺,是河南省天然林分布最大的區域,區內動植物種類繁多,分布有紅豆杉等多種珍稀植物種群[17]。目前,有關該區域紅豆杉種群的空間分布格局與種群數量結構及動態研究尚未見報道。2012—2014年對伏牛山自然保護區天然紅豆杉資源開展了專項調查,在此基礎上,本文就瀕危天然紅豆杉種群的空間分布格局與種群動態進行分析,以期為該地區瀕危植物紅豆杉種群的保護與恢復提供科學依據。
1 材料與方法
1.1研究地概況
伏牛山國家自然保護區位于河南省西南部,地理坐標為東經110°30′—113°05′,北緯32°45′—34°00′。伏牛山是秦嶺山脈向中原延伸的東端支脈,呈西北—東南走向,東西綿延約400 km,南北寬約40~70 km。海拔高度為500~2 200 m,平均海拔1 000 m。在我國地勢的三大階梯中,伏牛山正處在第二階梯向第三階梯過渡地帶,山體完整高大,山峰突兀聳立,地形、土壤復雜多樣;地理位置特殊,處在我國暖溫帶和北亞熱帶過渡區,也是華北、華中、西南植物的鑲嵌地帶,屬暖溫帶落葉闊葉林向北亞熱帶常綠落葉混交林的過渡區。由于高差大,區內氣候、土壤和自然植被的垂直帶譜明顯。伏牛山還是長江、黃河、淮河3大水系的分水嶺和一些支流的發源地,為重要的水源涵養林區。區內氣候溫暖濕潤,雨量充沛,年降水量800~1 100 mm,是著名的暴雨區;年最高氣溫26.5~28.5℃,最低氣溫-1.5~-2℃,年均溫12.1~15.1℃。復雜多樣的生態環境條件,區內生物資源多樣性豐富,僅珍稀、瀕危植物就有38科70種之多,中國特有屬37個,國家重點保護野生植物32種,是我國中部重要的植物資源寶庫,在研究植物區系、生態學等方面具有重要的科學價值[17]。
1.2樣地設置與調查
于2012—2014年,采用樣地調查法,對伏牛山自然保護區的天然紅豆杉種群進行調查。其中堯山泉水溝村:紅豆杉主要生長在縱長270 m山坡的中、上坡位,設置喬木調查樣方9個;寶天曼石鼓村:紅豆杉主要分布在縱長180 m山坡的下坡位,設置喬木調查樣方15個;玉皇山毛河村(玉皇山1):紅豆杉種群沿山谷呈狹長狀分布,主要集中在縱長約340 m山坡的中、上坡位,設置喬木調查樣方12個;柴家溝村(玉皇山2):紅豆杉主要分布在中、下坡位,設置喬木調查樣方14個。
喬木層每個樣方面積為20 m×20 m,對樣方內胸徑2 cm以上的喬木進行每木調查,記錄樹名、株數、樹高、胸徑、冠幅、蓋度等:用胸徑圍尺測量喬木胸徑;用測高器測量樹高;按南北和東西兩個方向,用皮尺測量樹冠在地面上的垂直投影的直徑,作為冠幅。在喬木樣方的四角和中心設置5個5 m×5 m的小樣方進行灌木層調查,記錄灌木種名、株數、高度、基徑、冠幅、蓋度等;用游標卡尺記錄灌木的基徑,用鋼卷尺測量株高和冠幅。隨機選取5個l m×1 m的小樣方進行草本層調查,記錄種名、株數、高度、蓋度、多度、干鮮重等。同時,對每個樣方的生境進行調查,記錄經緯坐標、海拔高度、地形起伏、坡向、坡位、坡度、土壤類型、土層厚度及剖面等。
1.3數據處理
1.3.1齡級結構分析本文采用空間序列替代時間變化的方法,應用大小結構分析法來研究種群年齡結構特征[18-19]。劉佳慶等[7]根據紅豆杉的生長規律和前人的研究結果,以樹高(H)結構反映種群年齡結構。參考此法,將伏牛山國家自然保護區的紅豆杉種群高度劃分為6個級別,以此代替該區域紅豆杉的年齡級(表1)。

表1 伏牛山國家自然保護區紅豆杉種群高度結構分級
注:Ⅰ為紅豆杉1級幼苗,包括Ⅰ1(1年生幼苗)和Ⅰ2(2~3年生幼苗);Ⅱ為紅豆杉幼樹;Ⅲ為紅豆杉小樹;Ⅳ為紅豆杉壯樹;Ⅴ為紅豆杉大樹;Ⅵ為紅豆杉老樹。
1.3.2種群空間分布格局分析采用聚集度指標,對天然紅豆杉種群的空間分布格局進行分析。聚集度指標包括方差均值比(DI)、負二項參數(K)、叢生指數(I)、平均擁擠指數(m*)、聚塊指數(Pai)和Green指數(GI),各指標計算公式及意義參考文獻[7,20—22]。
1.3.3種群結構與動態分析采用陳曉德[23]的量化方法,對紅豆杉的種群結構與動態進行分析。
(1) 種群內兩相鄰年齡級間個體數量變化動態(Vn):
(1)
式中:n——年齡級(高度級);Sn,Sn+1——第n,第n+1年齡級的個體數;Vn——種群個體數量的動態變化,其值為正值時表示種群內兩相鄰年齡級個體數量呈增長的動態趨勢,負值則表示衰退,零值表示穩定。
(2) 種群的年齡結構動態量值(P)由各年齡級間動態Vn對年齡級個體數(Sn)進行加權分配而得:
(2)
式中:k——年齡級數量。
(3) 考慮種群年齡級數量(K)及種群年齡級個體數(Sn)因素的種群年齡結構動態(E):
(3)
1.3.4種群靜態生命表和存活曲線采用靜態生命表編制方法[24]編制紅豆杉種群靜態生命表,選用一次性移動平均法對紅豆杉的種群齡級結構進行模擬和預測[25],公式如下:
(4)式中:n——需要預測的未來時間年限;Mt——未來n年時t齡級的種群大小;Xk——當前k齡級的種群大小。
2 結果與分析
2.1種群年齡-個體大小結構分析
由圖1可知,堯山泉水溝村紅豆杉種群共79棵,齡級結構不完整。幼苗數量少且全為根蘗苗,3年來的調查均未發現種子萌發苗;9 m以上的老齡樹缺失。總體上種群年齡結構呈正態分布,為穩定型種群。但因幼苗的嚴重缺失,未來可能會發生衰退演替。

圖1 伏牛山國家自然保護區紅豆杉種群年齡結構
寶天曼石鼓村紅豆杉更新種群653棵,齡級結構完整。有大量的更新幼苗,幼苗數占總數的67.69%;幼樹數量嚴重不足,僅占總數的2.14%。因Ⅱ級幼樹的嚴重不足和林齡偏大,未來將呈現衰退演替。
玉皇山毛河村(玉皇山1)紅豆杉更新種群276棵,種群齡級結構完整,結構形態近鐘形,呈現為齡級偏高的穩定型種群。其中Ⅲ齡級小樹個體數量的比例突然減少,僅為7.61%,將嚴重影響種群的未來發展。
玉皇山柴家溝村(玉皇山2)紅豆杉更新種群1 117棵,幼苗占總數的77.98%,老樹比例為1.61%,種群結構呈現基部寬頂部窄的金字塔型,屬增長型種群。
2.2種群空間分布格局
由表2可知,伏牛山各段的天然紅豆杉種群中各齡級的DI值都大于1,均符合聚集分布的特征。其中,堯山種群的Ⅰ,Ⅴ齡級的K值分別為0.15,0.40,數值較小,呈現較高的集群分布特征;而Ⅴ齡級的GI值為0.01,高度接近0,又符合隨機分布特征。寶天曼種群各齡級聚集度波動變化明顯:第Ⅰ齡級高度聚集,第Ⅱ齡級有所下降,第Ⅲ齡級則有所提高,而第Ⅳ,Ⅴ齡級明顯下降,第Ⅵ齡級又明顯提高。玉皇山南坡種群各齡級亦呈現由幼齡級到中齡級聚集度下降而由中齡級至大齡級又有所增加的趨勢;北坡種群從第Ⅰ齡級至第Ⅴ齡級均為集群分布,第Ⅵ齡級服從隨機分布。

表2 種群空間分布格局分析
注:SD為空間分布;C為集群分布;R為隨機分布;E為均勻分布;K為負二項參數;I為叢生指數;m*為平均擁擠指數;Pai為聚塊性指數;DI為方差均值比;GI為Green指數;t為T-test中的t值,查t值表所得。
總體上看,由幼齡級至中齡級,聚集程度下降而由中齡級至大齡級聚集程度增強,聚集度隨齡級增長而呈現U形動態變化。
2.3靜態生命表和存活曲線
由表3可知,伏牛山自然保護區內紅豆杉種群之間、種群內各齡級之間的消失率,有明顯的差異。堯山境內的天然紅豆杉種群,Ⅰ,Ⅱ齡級的消失率為負值,說明幼苗數量不足,該種群需要補充一定數量的幼苗來維持發展,否則將走向衰退;寶天曼種群在從Ⅰ齡級向Ⅱ齡級發展過程中的消失率極高,說明該齡級個體數量嚴重缺少,同時也說明該種群缺少種苗生長到幼苗的存活條件;玉皇山南坡種群中,Ⅲ,Ⅳ,Ⅴ齡級的消失率均為負值,說明種群個體數量波動明顯,中、幼齡級個體數量較少;玉皇山北坡種群,各齡級個體消失率低,且隨著齡級的升高而逐步減少,表明該種群更新狀態較為理想。
由圖2可以看出,伏牛山東段的堯山天然紅豆杉種群,存活曲線屬于Deevey-Ⅰ型,為衰退型的幼齡種群。伏牛山中部的寶天曼天然紅豆杉種群,存活曲線在Ⅰ,Ⅱ齡級間斜率較大,說明幼苗在向幼樹的生長過程中,死亡率很高;而在Ⅱ,Ⅲ,Ⅳ齡級間曲線平緩并逐級上升,而Ⅴ齡級向Ⅵ齡級生長的過程中再現較高的死亡率。總體來看,該種群存活曲線介于Deevey-Ⅰ,Deevey-Ⅱ型之間,但因Ⅱ齡級個體存活數量太少,未來將發生衰退演替。伏牛山西段的玉皇山內,毛河村(玉皇山1)天然紅豆杉種群的存活曲線屬于Deevey-Ⅱ型,而柴家溝(玉皇山2)種群的存活曲線屬于典型的Deevey-Ⅲ型,有良好的更新狀態。伏牛山3個不同地段的天然紅豆杉種群,存活曲線類型不同,表明其未來的發展態勢不同,東段種群將發生衰退演替,中段種群在未來一段時間內呈現穩定狀態,而西段種群將呈現增長態勢。分析原因,除種群或個體所處微環境的差異外,主要是伏牛山東段是低山丘陵區,人為活動較為頻繁,天然種群受之影響明顯。

表3 伏牛山區紅豆杉種群靜態生命表

圖2伏牛山紅豆杉種群存活曲線
2.4種群動態
從表4可以看出,堯山種群中第Ⅰ,Ⅱ齡級的Vn為負值,為衰退型結構,Ⅲ,Ⅳ級為增長型結構;寶天曼種群中,Ⅱ,Ⅲ,Ⅳ齡級為衰退型結構;Ⅰ,Ⅴ齡級為增長型結構。玉皇山南坡種群中,Ⅲ,Ⅳ,Ⅴ齡級為衰退型結構;玉皇山北坡種群各齡級均為增長型結構。從種群整體的P,E來看,所有種群均為正值,表明各個種群皆為增長型種群。其中玉皇山南坡種群E值0.001 4接近于0,說明該種群為穩定種群。
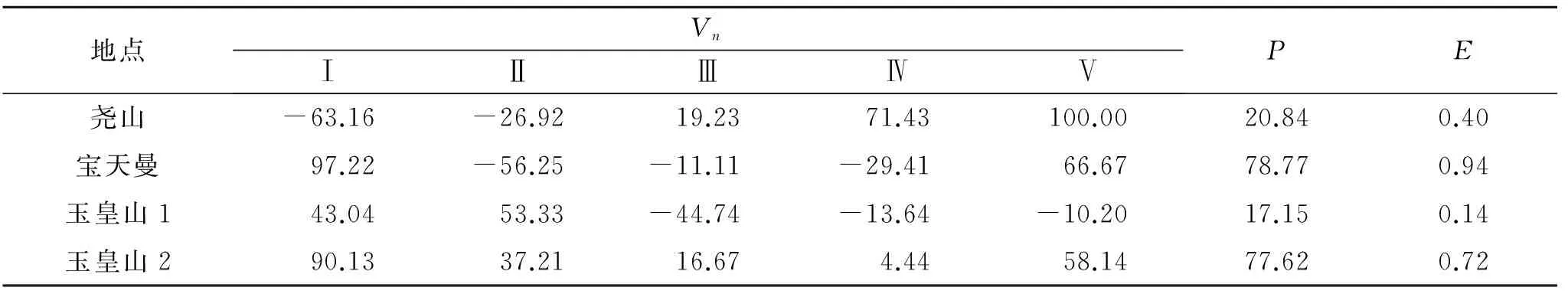
表4 伏牛山紅豆杉種群動態分析
由表5可以看出,不同生境的種群,動態變化規律有明顯差異。在未來2個齡級時間后,堯山紅豆杉種群Ⅱ,Ⅲ齡級的個體數量減少,Ⅳ,Ⅴ,Ⅵ齡級個體數量有少量增加;寶天曼紅豆杉種群Ⅱ齡級個體數量劇增,Ⅲ,Ⅳ,Ⅴ齡級個體數量均有所減少;玉皇山南坡種群Ⅱ,Ⅲ齡級個體數量微增,而Ⅳ,Ⅴ,Ⅵ齡級均微降;玉皇山北坡種群Ⅱ—Ⅵ齡級的個體數量均有所增加,Ⅱ齡級劇增。經過未來4個齡級時間后,堯山種群中Ⅳ齡級個體數量減少,而Ⅴ,Ⅵ齡級個體數量均呈現小幅度增加趨勢;寶天曼種群的Ⅵ齡級個體數量劇增,而Ⅴ齡級個體數量有所減少,Ⅵ齡級個體數量保持穩定;玉皇山南坡種群Ⅳ齡級個體數量增加,而Ⅴ,Ⅵ齡級個體數量均減少;玉皇山北坡種群Ⅳ,Ⅴ,Ⅵ齡級個體數量均增加。經過6個齡級時間后,各種群Ⅵ齡級個體數量,除了堯山種群穩定在上個時間外,其余3個地方的種群Ⅵ齡級個體數量均呈不同程度的增加。

表5 伏牛山紅豆杉種群動態時間序列分析
3 結論與討論
伏牛山東段的堯山紅豆杉種群和西段的玉皇山南坡種群存活曲線為Deevey-Ⅰ型,為衰退型種群;中段的寶天曼種群和西段的玉皇山北坡種群的存活曲線為Deevey-Ⅲ型,屬于增長型種群。在未來的2,4,6個齡級時間后,堯山種群和玉皇山南坡種群,由于幼苗數量不足,小樹數量呈現衰減趨勢;寶天曼種群的Ⅵ齡級個體數量呈現增長趨勢,表明該種群呈現增長動態;玉皇山北坡種群各齡級穩定增長,呈現良好的更新狀態。
種群動態反映了植物種群的未來命運。從種群的靜態生命表和存活曲線來看,在伏牛山紅豆杉種群的早期(如幼苗期、幼樹期)死亡率高,而中齡期以后則種群個體數量穩定。這說明紅豆杉幼苗和幼樹的生長過程中,存在嚴格的環境篩選,多數個體難以通過環境篩選而死亡。
處于伏牛山西段深山處的柴家溝村紅豆杉種群結構完整,而處于東段淺山區的泉水溝村的紅豆杉種群齡級結構不完整,種群的“大小級結構金字塔”并不呈現為三角形狀,這可能與結構齡級的劃分幅度有關,抑或是嚴酷的環境篩的作用結果。此外,頻繁的人為活動,干擾或破壞了紅豆杉的生境,致使紅豆杉數量減少,可能是造成某些“大小級結構”缺失的原因。
種群分布格局是物種與環境長期相互作用的結果。伏牛山各段的天然紅豆杉種群,均呈現聚集分布狀態;不同發育階段的種群,聚集程度不同,這與王磊等[13]的研究結果相一致。伏牛山自然保護區天然紅豆杉種群的空間分布格局呈現聚集分布,聚集程度隨齡級變化而呈U形動態變化,這種動態變化,客觀地反映了紅豆杉種群在生長發育過程中的生態策略和適應機制[22],因種子散落和母株發生萌蘗相對集中,導致幼苗期和幼樹期的紅豆杉呈現高度聚集分布;在后來的成長過程中,因個體所處的微環境不同,以及個體間對空間和養分的激烈競爭,導致幼苗和幼樹個體大批死亡而呈現聚集度有所下降。大樹以后階段的聚集度不降反升的生態機理,有待今后進一步研究。
特定時間生命表反映多個世代重疊的年齡動態歷程中的一個特定時間斷面,它是用同一時期收集到的種群所有個體的年齡數據編制而成,而不是對同生群的全部生活史追蹤。由于紅豆杉種群為天然林,研究方法是由“空間代時間”,所以調查所得的數據并不完全滿足編表的3個假設,在編制的生命表中會出現死亡率為負值的情況。對此,張文輝等[26]認為,這不僅客觀反映了種群波動真實情況,而且對種群數量預測有重要生態意義。天然植物種群各齡級株數波動是普遍現象,本文并未對數據作數據勻滑處理,似乎更有利于反映種群現實狀況。
由于紅豆杉屬國家一級重點保護植物,種群資源稀少,不能作伐木解析,而且用生長錐鉆木測年的方法也會對樹體造成一定的破壞。因此,本文采用了依據高度劃分齡級的方法。但生命表中年齡的劃分是以年為參照等距劃分的,依個體大小劃分時,年齡間距與個體大小等級并不是線形的。這樣編制生命表和繪制存活曲線,依據其線形判斷種群動態時,有時可能會有誤判。
本研究表明,伏牛山國家自然保護區紅豆杉種群前景堪憂,必須采取嚴格的封育措施對其和生境加以保護,嚴禁任何形式的破壞;建議輔以人工繁育措施,促進種群的更新與壯大。
[1]中國科學院植物研究所.中國高等植物圖鑒(第1冊)[M].北京:科學出版社,1972.
[2]鄧青姍,朱瓊瓊,魯長虎.南方紅豆杉的天然更新格局及食果鳥類對其種子的傳播[J].生態學雜志,2008,27(5):712-717.
[3]臧敏,卞新民.江西三清山珍稀瀕危植物考察研究[J].武漢植物學研究,2003,21(6):515-520.
[4]Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park, Chile[J]. Vegetatio,1992,98(1):13-22.
[5]Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercusgarryana) in British Columbia, Canada[J]. Forest Ecology & Management,2000,137(1):209-219.
[6]劉普幸.疏勒河中下游綠洲胡楊種群結構與動態研究[J].自然資源學報,2011,26(3):429-439.
[7]劉佳慶,李寧,熊天石,等.瀕危植物南方紅豆杉不同種群的結構和動態變化[J].熱帶亞熱帶植物學報,2014,22(5):479-485.
[8]張亞芳,李登武,王梅,等.黃土高原不同地區杜松種群結構與動態[J].林業科學,2015,51(2):1-10.
[9]李先琨,向悟生,蘇宗明.南方紅豆杉無性系種群結構和動態研究[J].應用生態學報,2004,15(2):177-180.
[10]李先琨.瀕危植物元寶山冷杉與南方紅豆杉種群生態學研究[M].北京:科學出版社,2006.
[11]李帥鋒,劉萬德,蘇建榮,等.云南蘭坪云南紅豆杉種群年齡結構與空間分布格局分析[J].西北植物學報,2013,33(4):792-799.
[12]李偉,王瑞雪,張光富,等.南方紅豆杉遷地保護種群的點格局分析[J].生態學雜志,2014,33(1):16-22.
[13]王磊,孫啟武,郝朝運,等.皖南山區南方紅豆杉種群不同齡級立木的點格局分析[J].應用生態學報,2010,21(2):272-278.
[14]茹文明,鐵軍,張桂萍,等.南方紅豆杉群落種間關系的數量分析[J].山西大學學報:自然科學版,2010,33(4):609-614.
[15]唐愽言,操國興,蔣萬平.武陵山大木山自然保護區紅豆杉群落優勢喬木種群生態位和種間聯結[J].植物分類與資源學報,2014,36(5):668-674.
[16]岳紅娟,仝川,朱錦懋,等.瀕危植物南方紅豆杉種子雨和土壤種子庫特征[J].生態學報,2010,30(16):4389-4400.
[17]王理順,申潔梅,李華堂,等.伏牛山自然保護區珍稀瀕危植物資源現狀與保護對策[J].河南林業科技,2003,23(3):42-44.
[18]Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed treeQuercusroburin two spruce-dominated forests[J]. Ecoscience,2000,7(1):38-44.
[19]李先琨,蘇宗明,向悟生,等.瀕危植物元寶山冷杉種群結構與分布格局[J].生態學報,2002,22(12):2246-2253.
[20]張金屯.植物種群空間分布的點格局分析[J].植物生態學報,1998,22(4):344-349.
[21]項小燕,吳甘霖,段仁燕,等.大別山五針松種內和種間競爭強度[J].生態學報,2015,35(2):389-395.
[22]李先琨,向悟生,歐祖蘭,等.瀕危植物南方紅豆杉種群克隆生長空間格局與動態[J].云南植物研究,2003,25(6):625-632.
[23]陳曉德.植物種群與群落結構動態量化分析方法研究[J].生態學報,1998,18(2):214-217.
[24]吳明開,沈志君,劉海,等.梵凈山自然保護區珙桐天然種群生命表與生存分析[J].生態學雜志,2012,31(6):1419-1424.
[25]韓路,王家強,王海珍,等.塔里木河上游胡楊種群結構與動態[J].生態學報,2014,34(16):4640-4651.
[26]張文輝,郭連金,劉國彬.黃土丘陵區不同生境沙棘種群數量動態分析[J].西北植物學報,2005,25(4):641-647.
Research on the Population Structure and Dynamics of the NaturalTaxuschinensisin Funiushan Mountain Area of He′nan Province
ZHANG Zhilu1, LIU Zhonghua1, CHEN Minghui1, ZHANG Qiaolian2, YANG Fengling1
(1.PingdingshanUniversity,Pingdingshan,He′nan467000,China;2.XuchangVocationalTechnicalCollege,Xuchang,He′nan461000,China)
Based on the aggregation indices and time-specific life table, the structure and dynamics of naturalTaxuschinensispopulation were studied. The results showed that: (1) the naturalTaxuschinensisresources distributed sporadically, but they relatively concentrated in the distribution area, and the aggregation class decreased first and then seemed to follow the U-shaped curve in the process of seedling to old trees; (2) the differences of survival amount were significant among the age classes, the survival curve followed the Deevey-Ⅲ type;(3) the mortality was higher at the stage of seedling and sapling, but it declined significantly after Ⅲ level (3 m). These results indicated that the trends of naturalTaxuschinensispopulation were a recession in Funiushan Mountain, high mortality of seedlings caused by harsh environmental sieve might be the main reason for the decline of population, meanwhile lack of resources protection induced the illegal felling trees, habitat destruction and so on also could not be ignored. So it is urgent to take measures to protect and restore the precious resources and to take the strict closure policy and auxiliary measures of artificial breeding.
endangered plant; life table; population structure;Taxuschinensis; time series analysis
2015-11-16
2015-12-20
河南省科技攻關計劃項目“中國中部伏牛山特有紅豆杉生態適應性與繁育關鍵技術研究”(122102310652);河南省環境科學重點學科(Pxy-zdxk-2013003);平頂山學院高層次人才科研啟動經費(2012005)
張志錄(1967—),男,河南上蔡人,博士,副教授,主要從事種群生態學和園藝學研究。E-mail:276996899@qq.com
楊風嶺(1963—),男,河南許昌人,博士,教授,主要從事環境科學和植物化學研究。E-mail:13903900069@163.com
S718.5
A
1005-3409(2016)03-0262-07
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
