荒漠區嚙齒動物優勢種與植物因子的關系
2016-10-14 20:14:09劉永召龍華趙繼東楊路余俊國
安徽農業科學 2016年9期
劉永召 龍華 趙繼東 楊路 余俊國
摘要[目的]明確荒漠區在不同干擾下植物因子與嚙齒動物優勢物種之間的相互影響。[方法]研究西北某干旱地區輪牧與過牧干擾下嚙齒動物優勢種種群的生物量和植物因子之間的關系,并進行灰色關聯度及回歸分析。[結果]優勢物種三趾跳鼠、子午沙鼠和小毛足鼠均與灌木植物有較高的關聯度,植物因子對該地區優勢鼠種生物量的作用有明顯差異,從而形成相互適應、長期共存的格局。[結論]揭示了植物生物量與嚙齒動物生物量的變化關系,為研究該荒漠區嚙齒動物優勢物種群落分布規律提供了參考。
關鍵詞 嚙齒動物;優勢種;植物因子;輪牧;過牧
中圖分類號 X171.1 文獻標識碼 A 文章編號 0517-6611(2016)09-022-03
Abstract[Objective]The aim was to study the interaction effect between plant factors and dominant species of rodents in desert region under different disturbance.[Method]The relationship between biomass of rodent dominant species and plant factors under rotational grazing and overgrazing disturbance in desearch region in northwest region was studied, the grey correlation degree and regression analysis was conducted.[Result]The dominant species like threetoed Jerboa, Meriones meridianus and Xiaomao foot mouse had higher correlation with shrub plants. Plant factors had obvious differences on biomass of dominant species, thus forming the pattern of mutual adaptation, longterm coexistence.[Conclusion]The relationship between plant biomass and the biomass of rodents is revealed, which provides reference for study on distribution law of dominant species of rodent in desert region.
Key words Rodent; Dominant species; Plant factor; Rotational grazing; Overgrazing
由于近年來人為干擾不斷加重, 使得溫帶草地的荒漠化日益嚴重。依賴于草地植物生存的草食動物種群和群落格局隨之受到明顯影響[1]。嚙齒動物群落是荒漠生態系統食物鏈上必不可少的消費者[2],探究合理的嚙齒動物群落格局與植物群落的生長狀態關系對于荒漠區的環境與發展至關重要[3-6]。目前,有關荒漠區嚙齒動物與植物群落關系的研究逐漸得到重視,很多學者對不同干擾下荒漠區嚙齒動物群落構成、作用以及與生境中植物因子的影響關系進行了探討[7-10]。筆者針對西北某荒漠地區,在人為的輪牧與過牧干擾條件下研究了嚙齒動物優勢種與其棲息地植物因子之間的相互關系,旨在為研究該荒漠區嚙齒動物優勢物種群落分布規律提供理論依據。
1 數據來源與方法
1.1 數據來源 研究區地處西北某干旱地區,調查樣地包括過牧區和輪牧區。研究對象為各區域的草本植物、灌木植物和嚙齒動物優勢種。由于該荒漠區植被類型較少,選取主要植物類型為研究對象。將植物因子依據生長類型劃分為草本植物因子和灌木植物因子,其中植物因子的統計數據為植物的高度、蓋度、密度和生物量(干重)。調查季節為夏季(7月)和秋季(10月)。動物因子選取該地區嚙齒動物優勢物種三趾跳鼠、子午沙鼠和小毛足鼠[7-9],以相應物種的動物生物量為統計數據。
1.2 研究方法 于2005~2007年的7、10月上旬在過牧區和輪牧區采用百夾法捕獲鼠類,每次隨機抽取22個樣方,每個樣方規格為10 m×10 m,放置100個捕鼠夾,以鼠的捕獲率作為相對密度指標[10]。對樣方內相應調查灌木類植物的高度、蓋度、密度和生物量(干重)進行測定,再對隨機樣方內1 m×1 m小樣方草本植物的高度、蓋度、密度和生物量(干重)進行測定。
1.3 數據處理 試驗數據采用matlab進行灰色關聯度計算,用Excel和SPSS軟件進行正態檢驗和回歸分析。
2 結果與分析
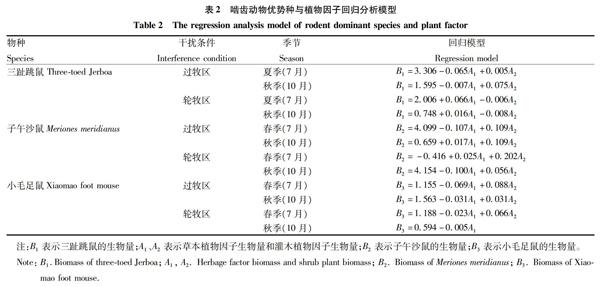
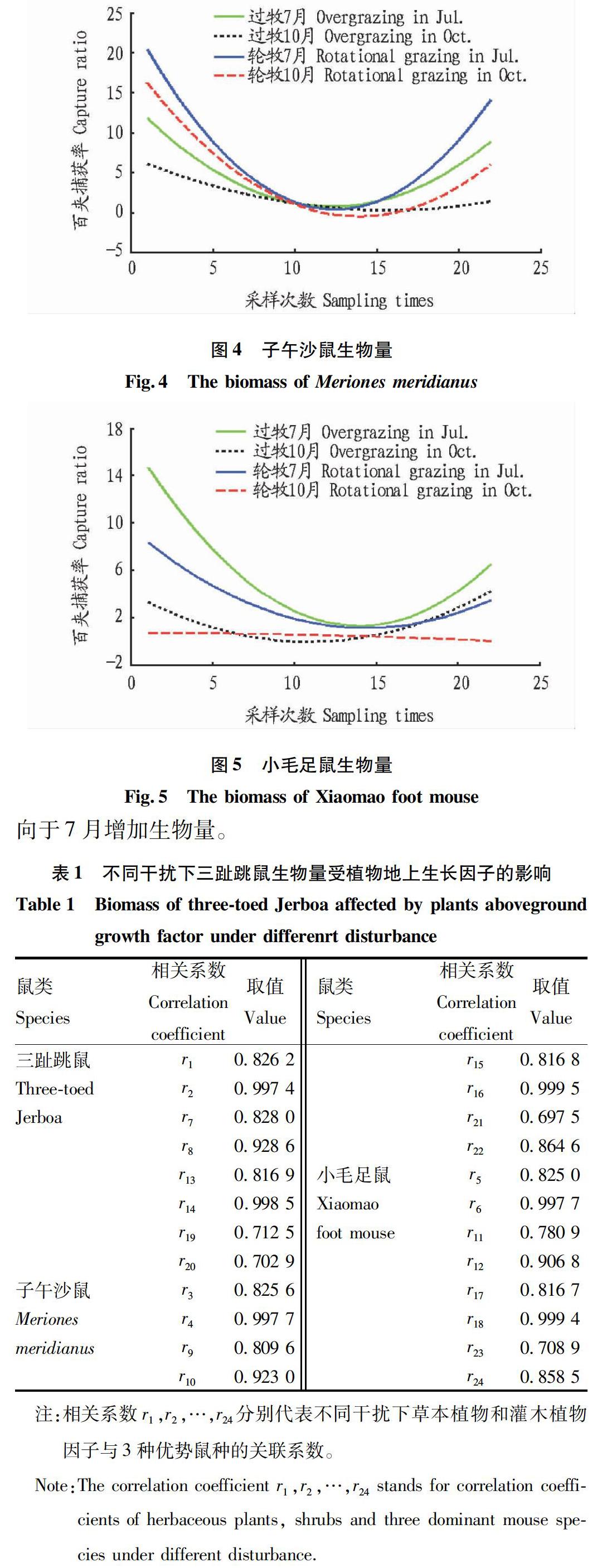
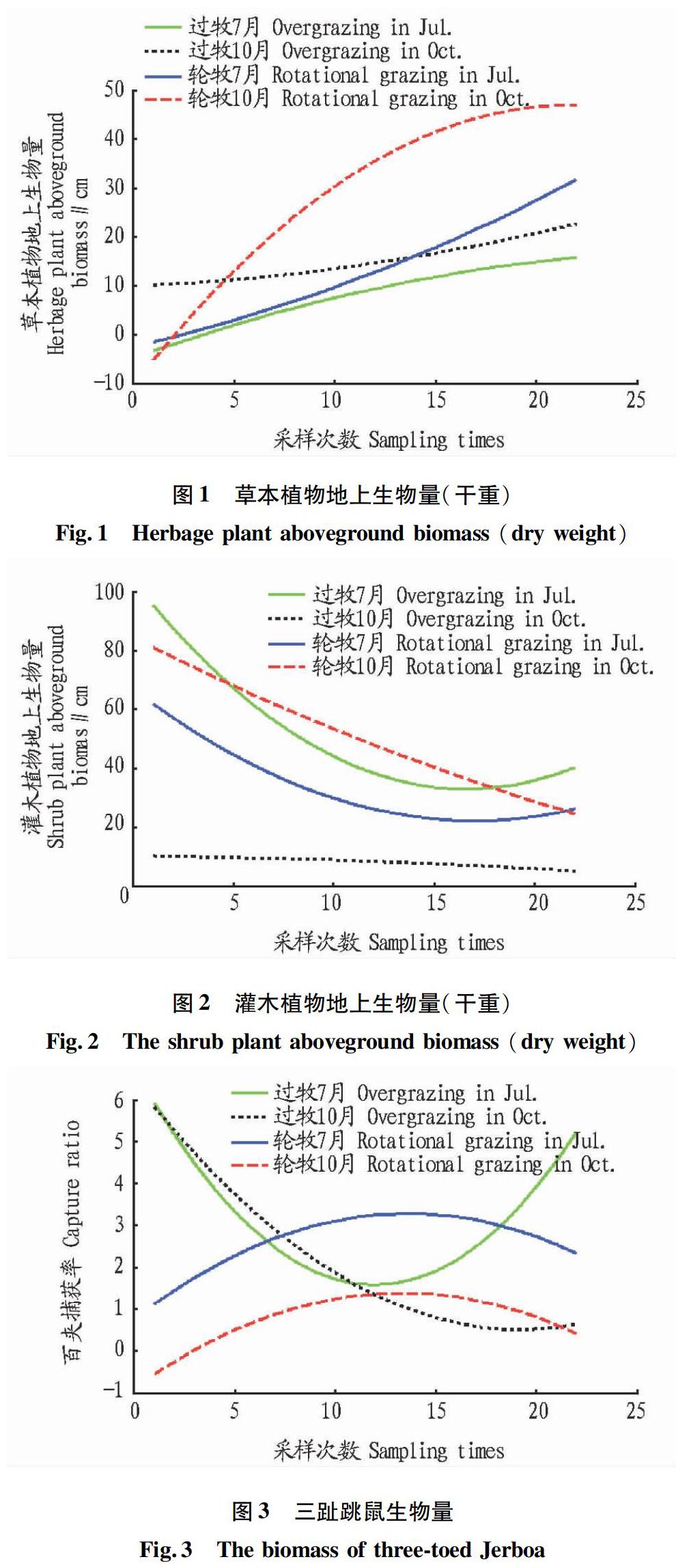
2.1 灰度關聯度分析不同干擾下各優勢鼠種受植物地上生長因子的影響 將2005~2007年測得的22組不同干擾條件下植物地上生物量離散值繪制成曲線(圖1、2)。三趾跳鼠、子午沙鼠、小毛足鼠的百夾捕獲量見圖3~5。以過牧區夏季(7月)的植物地上生物因子為灰色關聯度分析的研究對象,將其分為草本植物因子地上生物量和灌木植物因子地上生物量,以二者作為比較序列;同時,將待研究的動物因子生物量分別作為參考序列,進行灰色關聯度分析。
2.1.1 灰色關聯性分析優勢鼠種與植物因子相關系數。以過牧區7月三趾跳鼠與草本植物地上生物量和灌木植物地上生物量的灰度關聯為例進行分析。草本植物生物量、灌木植物生物量構成比較數列,用相應的動物生物量數據作為參考數列,計算出不同干擾下三趾跳鼠、子午沙鼠、小毛足鼠受植物地上生長因子的影響,結果見表1。
2.1.2 結果分析。在不同干擾下與三趾跳鼠生物量有關的各因子中,輪牧7月、過牧7月和過牧10月的灌木植物地上生物量具有較主要的影響,表明三趾跳鼠較喜歡在灌木環境下生長,且其傾向于選擇7月增加生物量。同時,子午沙鼠和小毛足鼠的習性與三趾跳鼠相似,以灌木植物地上生物量為顯著影響,傾向于7月增加生物量。小毛足鼠一年可繁殖1~2次,每年4月開始繁殖,10月結束,因此,小毛足鼠更傾向于7月增加生物量。
2.2 KS檢驗 由于灰色關聯分析只顯示相關大小,不能顯示他們之間的關聯是正相關還是負相關,故采用KS檢驗,提高結果的有效性。
2.2.1 對過牧區和輪牧區夏、秋季植物因子采樣數據正態分布KS檢驗。對該荒漠地區過牧區夏季(7月)草本植物因子(包括高、蓋、密、生物量)的樣本數據進行單樣本KS檢驗。結果表明,該組樣本數據均滿足正態分布。同樣檢驗過牧區夏季(7月)的灌木植物因子、過牧區秋季(10月)的植物因子以及輪牧區夏、秋季的植物因子,結果表明樣本數據均符合正態分布。
對荒漠地區7月的過牧區嚙齒動物生物量、10月的過牧區嚙齒動物生物量、7月的輪牧區嚙齒動物生物量、10月的輪牧區嚙齒動物生物量的樣本數據進行單樣本KS檢驗。結果表明,樣本數據均滿足正態分布。
2.2.2 對過牧區和輪牧區夏、秋季動物因子優勢種采樣數據正態分布KS檢驗。用SPSS統計學軟件對植物生物量與嚙齒動物生物量之間的關系進行回歸分析,結果見表2。
2.2.3 結果分析。分析線性回歸模型可知,7月過牧區草本植物生長最好時,放牧造成草本被吃掉,所以對三趾跳鼠產生負影響,灌木更多地為三趾跳鼠的生長和繁殖提供所需條件。子午沙鼠在過牧區的生物量比輪牧區多,7月過牧區灌木也比輪牧區多,說明子午沙鼠更多地依賴灌木生存,而放牧使得草本對子午沙鼠產生負影響。7月過牧區小毛足鼠比輪牧區多,灌木比輪牧區也要多,所以灌木主要影響小毛足鼠,由于放牧,草本負向影響小毛足鼠的生物量;10月,小毛足鼠生物量減少,灌木有所減少,其他影響與7月類似;此時草本和灌木都較穩定,主要因為小毛足鼠依賴于灌木生存,所以灌木正相關,無放牧影響,草本負相關;此時草本增長太快,小毛足鼠生物量減少,所以草本生物量和小毛足鼠生物量負相關。
3 結論
通過對草本生物量、灌木生物量、嚙齒動物生物量進行正態分布檢驗,對植物生物量與嚙齒動物生物量回歸分析,得到回歸數學模型,揭示了植物生物量與嚙齒動物生物量的變化關系。結果表明,三趾跳鼠、子午沙鼠、小毛足鼠生物量作為因變量,草本植物因子生物量和灌木植物因子生物量作為自變量,得出過牧主要對草本產生較大影響,對灌木影響不大,輪牧條件下草本和灌木生物量都是自然變化;子午沙鼠在過牧區較多是由于其對灌木的過度依賴,而三趾跳鼠和小毛足鼠在過牧區生物量都比輪牧區少,說明過度放牧導致三趾跳鼠和小毛足鼠生物量的降低。
參考文獻
[1]王德利,王嶺.草食動物與草地植物多樣性的互作關系研究進展[J].草地學報,2011,19(4):699-704.
[2]楊維康,蔣慧萍,王雪芹,等.古爾班通古特沙漠區大沙鼠對荒漠植物群落的擾動效應[J].生態學雜志,2009,28(10):2020-2025.
[3]楊維康,喬建芳,蔣慧萍,等.大沙鼠掘洞對準噶爾荒漠植物群落的小尺度影響[J].干旱區地理,2006,29(2):219-224.
[4]付和平,武曉東,楊澤龍.不同干擾條件下荒漠嚙齒動物生態位特征[J].生態學報,2005,25(10): 2637-2643.
[5]陳善科,保平,張學英.阿拉善荒漠草地生態危機及其治理對策[J].草原與草坪,2000(2):9-11.
[6]王利清,楊玉平,張福順,等.呼和浩特地區嚙齒動物群落年度動態研究[J].中國草地學報,2013,35(5):118-123.
[7]MELO A S,RANGEL T F L V B,DINIZFILHO J A F.Environmental drivers of betadiversity patterns in New—World birds and mammals[J].Eeography,2009,32(2):226-236.
[8]王利民,周延林,鮑偉東.鄂爾多斯高原沙地嚙齒動物群落資源利用研究Ⅰ:空間資源維利用研究[J].內蒙古大學學報(自然科學版),2001,32(5):567-574.
[9]袁帥,付和平,武曉東,等.荒漠嚙齒動物群落對開墾干擾的響應及其種群生態對策[J].生態學報,2013,33(20):6444-6454.
[10]QIAN H,BADGLEY C,FOX D L.The latitudinal gradient of beta diversity in relation to climate and topography for mammals in North America[J].Global ecologyand biogeography,2009,18(1):111-122.
