青藏高原特有植物塊莖堇菜異型性種子萌發特性及其生態適應
2016-10-14 17:29:44侯云云吳瓊高婷
安徽農業科學 2016年9期
侯云云 吳瓊 高婷


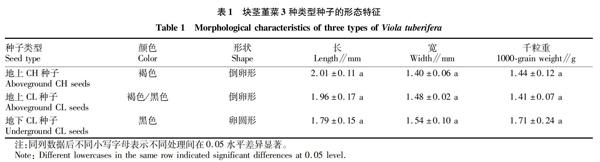
摘要[目的]明確青藏高原特有植物塊莖堇菜異型性種子的萌發特性,為進一步揭示其生理生態適應機制提供理論依據。[方法]研究塊莖堇菜地上開放花(CH)種子、地上閉鎖花(CL)種子和地下CL種子3種異型體種子的形態特征及萌發特性。[結果]3種類型的種子顏色和形狀存在差異,種子大小和質量無顯著差異;CH種子和地上、地下CL種子萌發率均較低,最高分別為35.56%、33.33%和3.33%;延長儲藏時間均能顯著促進地上CH和CL種子的萌發,也能部分提高地下CL種子的萌發率。[結論]塊莖堇菜異型性種子不同的萌發特性提高了該物種在高度異質性環境中的適合度,對種群能成功地適應高寒環境具有重要意義。
關鍵詞 塊莖堇菜;種子異型性;生態適應
中圖分類號 S311 文獻標識碼 A 文章編號 0517-6611(2016)09-010-03
Abstract[Objective]To find the germination characteristics of heteromorphic seeds of Viola tuberifera in Qinghai-Tibet Plateau, and to further reveal their physiological traits.[Method]Morphological characteristics and germination characteristics of three heteromorphic seeds of V.tuberifera were researched, including the aboveground CH (chasmogamous flowers) seeds, aboveground CL (cleistogamous flowers) seeds and underground CL (cleistogamous flowers) seeds.[Result]The three types of seeds were different in color and shape, but they did not have significant differences in size sand mass.The germination rates of aboveground CH seeds and two types of CL seeds were relatively low, which were 3556%, 33.33% and 3.33%, respectively.Prolonging storage period could significantly increase the germination of aboveground CH and CL seeds, and partly enhanced the germination rate of underground CL seeds, suggesting that aboveground CH and CL seeds were nondeep physiological dormancy; while underground CL seeds belonged to deep physiological dormancy.[Conclusion]Germination characteristics of heteromorphic seeds of V.tuberifera increase the species fitness to heterogeneous habitats and form an ecological adaptive strategy that may allow V.tuberifera to successfully adapt to the alpine environment.
Key words Viola tuberifera; Seed heteromorphism; Ecological adaptation
種子萌發階段是植物種群更新和物種得以繁衍的關鍵性環節,其萌發特性由種子自身及外界環境因子共同決定[1-2]。植物在種子萌發階段對環境的適應能力是植物對環境適應性的最重要體現[3]。與無性繁殖相比,種子繁殖在維持種群的遺傳多樣性、提高種群的潛在適合度進而影響種群未來的命運中具有更重要而積極的意義,但種子繁殖所產生的幼苗更易受不利環境的影響,因而具有更高的死亡風險[4]。因此,種子萌發階段是植物生活周期中最重要和最脆弱的階段[5-6]。
種子異型性是植物在種子階段來適應環境所采取的一種特殊策略,指同一植株產生多種在形態結構及萌發特性等方面存在顯著差異的種子[7-8]。異型性種子往往存在不同的形態學特征、傳播特點、萌發和休眠特性[9-13],它們的萌發在時間上交替出現,逃避了不利的生境,降低了萌發風險,有利于種群建成[13]。具有種子異型性的植物主要分布于沙漠、干旱半干旱、鹽漬土地區及擾動強烈地區[14],這是植物在嚴酷多變的生境條件下長期進化的結果[13]。種子異型性被認為在避免密集負效應、采取“兩頭下注”策略以及適應時空異質化環境等方面具有重要的進化生態意義[4,14],對了解植物的生態適應機制有重要價值[15]。近年來,種子異型性研究已成為國際上適應對策研究中的熱點問題和植物進化生態學的重要研究方面[16]。在被子植物中,種子異型性現象已在26科129屬292種上進行了報道[16],國內僅報道了19種[15-16],主要集中在藜科、菊科等5個科,關于堇菜科種子異型性現象尚未見報道。
塊莖堇菜(Viola tuberifera)屬堇菜科堇菜屬,為青藏高原特有的早春開花細弱草本植物,產于海拔2 500 m以上的高寒草甸[17],可通過完全開放花(CH)異交繁殖和地上閉鎖花(CL)嚴格的自交繁殖外,還可通過地下匍匐莖節產生大量的CL花進行自交繁殖[18],分別產生CH種子和地上、地下CL種子。塊莖堇菜是堇菜屬植物中繁殖方式最特殊的種類,國內外尚未見其相關報道。鑒于此,筆者研究了塊莖堇菜3種類型種子的形態特征及萌發特性,以期為揭示其生殖生態適應機理提供理論依據。
參考文獻
[1]何芳蘭,趙明,王繼和,等.幾種荒漠植物種子萌發對干旱脅迫的響應及其抗旱性評價研究[J].干旱區地理,2011,34(1):100-105.
[2]張景寬,張文輝,張潔明,等.種子萌發期4種植物對干旱脅迫的響應及其抗旱性評價研究[J].西北植物學報,2006,26(9):1811-1818.
[3]孫坤,唐潔娟,蘇雪,等.青藏高原特有植物肋果沙棘種子萌發對不同溫度的響應[J].西北師范大學學報,2009,45(3):83-86.
[4]馬文寶,譚敦炎,薛建輝,等.果實(種子)多態性及生態學意義[J].四川林業科技,2011,32(6):34-43.
[5]徐恒恒,黎妮,劉樹君,等.種子萌發及其調控的研究進展[J].作物學報,2014,40(7):1141-1156.
[6]RAJJOU L,DUVAL M,GALLARDO K,et al.Seed germination and vigor[J].Annu Rev Plant Biol, 2012,63:507-533.
[7]IMERT E.Ecological consequences and ontogeny of seed herteromorphism[J].Perspectives in plant ecology,evolution and systematic, 2002,5(1):13-36.
[8]VENABLE D L.The evolutionary ecology of seed heteromophism[J].American naturalist, 1985,126(5):577-595.
[9]MAUN M A,PAYNE A M.Fruit and seed polymorphism and its relation to seedling growth in the genus Cakile[J].Canadian journal of botany, 1989,67:2743-2750.
[10]魏巖,劉鵬偉,安沙舟.野榆錢菠菜的果實多型性及其萌發對策[J].干旱區研究,2007,24(6):835-839.
[11]王宏飛,魏巖,黃振英.散枝豬毛菜的種子多型性及其萌發行為[J].植物生態學報,2007,31(6):1046-1053.
[12]高蕊,魏巖,嚴成.角果藜的地上地下結果性與種子萌發行為[J].生態學雜志,2008,27(1):23-27.
[13]楊帆,曹德昌,楊學軍,等.鹽生植物角果堿蓬種子二型性對環境的適應策略[J].植物生態學報,2012,36(8):781-790.
[14]LEVYADUN S.Why are underground flowering and fruiting more common in Israel than anywhere else in the world[J].Current science, 2000,79:289.
[15]吳冬,黃姝博,李宏慶.意大利蒼耳二型性種子萌發、植株生長差異及生態適應性[J].生態學報,2009,29(10):5258-5263.
[16]王雷,董鳴,黃振英.種子異型性及其生態適應意義的研究進展[J].植物生態學報,2010,34(5):578-590.
[17]孫坤,孫文斌,蘇雪,等.青藏高原東緣塊莖堇菜克隆多樣性及其與生境特征的相關性分析[J].西北師范大學學報,2010,46(4):74-77.
[18]鞏明明,蘇雪,孫坤.青藏高原特有植物塊莖堇菜地上地下結果性及其散布機制[J].草業科學,2009,26(4):16-19.
[19]VENABLE D L.The evolutionary ecology of seed heteromophism[J].American naturalist,1985,126(5):577-595.
[20]吳澤昂,李予霞,莊麗,等.異型狹果鶴虱的二型性種子萌發特性及其生態適應[J].新疆農業科學,2013,50(20):307-312.
[21]BASKIN C C,BASKIN J M.Seeds ecology, biogeography, and evolution of dormancy and germination[M].San Diego: Academic Pres, 1998.
[22]劉鵬偉, 魏巖.準噶爾荒漠異苞濱藜(Atriplex mcrantha)的種子二型性及其萌發行為[J].生態學報, 2007,27(10):4233-4239.
[23]劉艷芳, 魏巖, 嚴成.異子蓬二型種子的萌發與休眠特性及其生態適應[J].生態學報, 2009,29(12):6609-6614.
