冰溫貯藏中南美白對蝦特定腐敗菌的分離鑒定及其腐敗能力分析
2016-09-10 06:04:40謝麗丹李蕾蕾王素英董世瑞
食品工業科技 2016年5期
謝麗丹,李蕾蕾,王素英,董世瑞
(天津市食品生物技術重點實驗室 天津商業大學生物技術與食品科學學院,天津 300400)
?
冰溫貯藏中南美白對蝦特定腐敗菌的分離鑒定及其腐敗能力分析
謝麗丹,李蕾蕾,王素英*,董世瑞
(天津市食品生物技術重點實驗室 天津商業大學生物技術與食品科學學院,天津 300400)
為了分離鑒定南美白對蝦在冰溫貯藏條件下的腐敗菌,并分析其腐敗能力的大小。通過凍結實驗,確定南美白對蝦的冰點,在0 ℃與冰點之間貯藏南美白對蝦,從中分離腐敗菌,進行形態學觀察和分子生物學鑒定,將分離得到的腐敗菌接種至滅菌蝦汁中,通過產硫化氫,降解蛋白質,產揮發性鹽基氮(TVB-N)的測定衡量腐敗能力的大小。結果表明:從南美白對蝦中分離得到12株腐敗菌,分別是Shewanellahafniensis,Acinetobacterjohnsonii,Planoccoccuscitreus,Bacilluscereus,Acinetobacterbeijerinckii,Enterobacterhormaechei,Arthrobacterbergeri,Bacilluslicheniformis,未鑒定出的菌株X1和X10。在冰溫貯藏條件下,導致南美白對蝦腐敗的優勢腐敗菌依次是Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii。
南美白對蝦,冰溫貯藏,腐敗菌,分離鑒定,腐敗能力
鮮蝦由于營養豐富、含水量高,極易被微生物污染而導致腐敗變質。相關研究表明,在不同條件下引起新鮮水產品腐敗變質的優勢菌不同,且優勢菌占菌落總數的比例會隨著貯藏時間的延長而不斷增加[1]。
冰溫貯藏指在冰點溫度范圍內貯藏食品[2]。與凍藏相比,冰溫貯藏南美白對蝦,不僅可以有效降低冷藏設備的能耗,還可以克服冰結晶帶來的蛋白質變性、組織結構損傷等現象,與冷藏相比其貯藏期得到顯著延長[3]。就品質而言,冷藏的南美白對蝦,腐敗微生物迅速增加,美味因冷藏時間增加而降低;冷凍貯藏則因生物細胞破壞,解凍營養流失,風味降低;而冰溫貯藏使南美白對蝦營養風味物質增加,且腐敗微生物增長速度相對較慢,較好地保持了鮮蝦特有的美味[4]。
許多報道認為溫度是影響南美白對蝦貯藏時間的關鍵因素,趙海鵬等[5]研究了冷藏條件下南美白對蝦腐敗菌的菌相分析,蘇永玲等[6]報道了南美白對蝦冷凍保鮮研究進展,但對南美白對蝦冰溫貯藏尤其有關優勢腐敗菌的研究鮮有報道。本文擬對冰溫貯藏南美白對蝦的主要腐敗菌進行分離純化,采用形態學特征、生理生化實驗以及16S rRNA序列相結合的方法,對優勢腐敗菌進行鑒定,在此基礎上測定其腐敗能力,為尋求既能保證鮮蝦品質,又能相對延長貨架期的保鮮方案奠定基礎。
1 材料與方法
1.1材料與儀器
鮮活南美白對蝦天津市南開區王頂堤水產批發市場;PCR擴增試劑及引物生工生物工程(上海)股份有限公司購買與合成;其它化學試劑北京奧博星生物技術有限公司。
UDK-159凱氏定氮儀意大利VELP公司;MLS-3780高壓滅菌鍋日本SANYO電器集團;SPX-150B-D型振蕩培養箱上海博迅實業有限公司醫療設備廠;PCR擴增儀美國ABI公司;JY04S-3C凝膠成像系統北京君意東方電泳設備有限公司。
1.2實驗方法
1.2.1實驗材料預處理購買時即在鮮活南美白對蝦中放入少許冰塊,充氧氣包裝后保活運輸至實驗室,立刻用碎冰使鮮活南美白對蝦休克,用清水洗凈瀝干后,按5只/袋標準裝入保鮮袋置于冰溫貯藏。
1.2.2南美白對蝦冰點的測定將溫度測量電極探針儀器插入南美白對蝦中心肌肉,放在-30 ℃,每2 s采集,繪制時間-溫度曲線,根據凍結曲線的拐點判定南美白對蝦的冰點,取多點測定的平均值作為冰溫貯藏溫度。
1.2.3腐敗菌的分離純化及計數將冰溫貯藏中的南美白對蝦在無菌條件下定期取樣制成勻漿,在貨架期內(8 d)每隔2 d采用梯度稀釋、有氧平板菌落計數法[7]進行計數,每個稀釋梯度計數3個平板,取平均值。采用平板劃線分離法反復純化直至獲得菌落純菌株。
1.2.4腐敗菌的鑒定觀察菌落形態,菌體的大小、形態、排列方式和芽孢的有無。在此基礎上采用傳統的酚/氯仿抽提法提取腐敗菌基因組DNA[8-10],并采用通用引物27F(AGAGTTTGATCCTGGCTCAG)和1492R(CTACGGCTACCTTGTTACGA)進行16S rRNA基因的PCR擴增。PCR反應體系總體積50 μL,包括模板DNA2.0 μL,正反向引物 各2.0 μL,2×Mix 25 μL,ddH2O 19 μL。PCR擴增參數設置為:94 ℃預變性4 min后,按照94 ℃ 變性1 min;,55 ℃ 退火1 min;,72 ℃延伸1 min進行28個循環,最后于72 ℃延伸10 min,反應結束后PCR產物經過1%瓊脂糖凝膠電泳檢測,符合測序要求的擴增產物由生工生物工程(上海)股份有限公司測序,將測序所得的16S rRNA基因序列信息在韓國標準菌株EzTaxon-e數據庫[11]中利用Identify程序進行同源性比對分析,挑選出與其同源性最高的標準菌株序列,采用軟件MEGA 6.0程序,構建系統發育樹。結合形態特點和系統發育位置,給出腐敗菌株鑒定結論。
1.2.5腐敗菌致腐能力的測定參照徐振偉[14-15]的方法制備無菌蝦汁,將5 μL濃度為106CFU/mL的菌懸液加入到10 mL滅菌蝦汁中進行冰溫貯藏,以未接種的蝦汁為對照,定時取樣采用凱氏定氮法測定揮發性鹽基氮(TVB-N)[16-17],考馬斯亮藍法測定蛋白質含量[12]。同時將腐敗菌接種到三糖鐵培養基中觀察其產硫化氫的情況[12]。
1.3數據處理
文中圖表采用Excel軟件處理,利用SPSS17.0軟件進行數據的差異顯著性分析,Origin Pro V8.0軟件繪制曲線。
2 結果與分析
2.1南美白對蝦冰點測量
利用多點溫度測定儀,測定南美白對蝦凍結曲線,結果見圖1。可知在-2.2 ℃時出現了過冷點,后上升至-1.4 ℃,維持短暫時間后迅速下降同環境溫度一致,根據這種溫度變化現象,可以判斷溫度反彈上升到最高時的溫度即為南美白對蝦的冰點,即為-1.4 ℃,可以確定南美白對蝦的冰溫帶為-1.4~0 ℃[18]。由于冰溫庫的溫度波動在±0.5 ℃,由此將冰溫庫的溫度設置為-0.8 ℃。

圖1 南美白對蝦凍結曲線Fig.1 The freezing temperature curve of Penaeus vannamei
2.2腐敗菌的分離鑒定
根據菌落顏色、隆起程度、表面含水狀態、邊緣是否整齊和透明程度,將分離自冰溫貯藏南美白對蝦的腐敗細菌歸為12種,觀察它們的個體形態及相關特征結果見表1。
對形態存在明顯差異的12株細菌進行16S rRNA基因PCR擴增,擴增產物在1%瓊脂糖凝膠電泳中均顯示約1.5 kb的明亮條帶,與預期片段大小相符(見圖2)。將測序所得16S rRNA基因序列在韓國標準菌株數據庫中利用Identify程序進行同源性比對分析,采用軟件Mega 6.0程序,構建系統發育樹,結果見圖3。
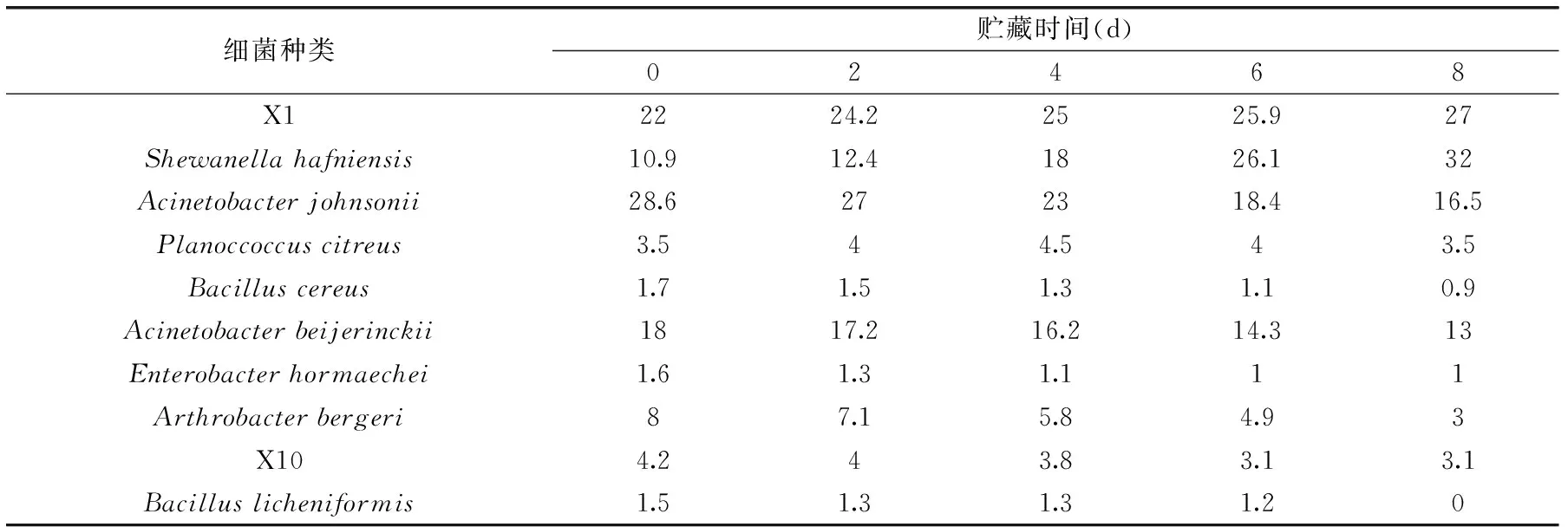
表2 南美白對蝦冰溫貯藏過程中腐敗菌的菌相組成(%)Table 2 Composition of spoilage bacteria of penaeus vannamei during ice-temperature storage(%)

表1 冰溫貯藏南美白對蝦腐敗菌的菌體形態及相關指標Table 1 Bacterial morphology and related indexes of Penaeus vannamei spoilage bacteria during ice temperature storage

圖2 16S rRNA基因PCR擴增產物的電泳圖譜Fig.2 Electrophorogram of PCR amplification products of 16S rRNA genes
根據同源性≥98%為同一細菌中的共識[19],結合形態特征指標,由圖3可知,X2為哈夫尼希瓦氏菌(Shewanellahafniensis),X3與X4為約式不動桿菌(Acinetobacterjohnsonii),X5為檸檬色動性球菌(Planoccoccuscitreus),X6為蠟樣芽孢桿菌(Bacilluscereus),X7與X12為Acinetobacterbeijerinckii,X8為霍氏腸桿菌(Enterobacterhormaechei),X9為Arthrobacterbergeri,X11為地衣芽孢桿菌(Bacilluslicheniformis)。X1和X10與不動桿菌屬的菌種親緣關系最近,但與任何一種的16S rRNA基因同源性均遠遠低于98%的水平,因此初步判斷它們可能為不動桿菌Acinetobacter的潛在新種或新屬,且根據革蘭氏染色結果和芽孢有無,可以確定X1與X10為不同種或不同屬細菌。
2.3腐敗菌在貯藏過程中的變化規律
在南美白對蝦冰溫貯藏貨架期內,定時對各種腐敗菌進行計數,并計算占總菌數的比例,探討腐敗菌的菌相組成變化規律,結果見表2和圖4。

圖3 基于16S rRNA基因序列的系統發育樹Fig.3 Phylogenetic tree based on sequence of 16S rRNA gene
由表2可以看到,新鮮南美白對蝦的初始菌相中,Acinetobacterjohnsonii和X1菌株的數量最大,分別占28.6%和22%,其次為Acinetobacterbeijerinckii和Shewanellahafniensis,分別占18%和10.9%,再次為Arthrobacterbergeri和Planoccoccuscitreus,分別占8%和3.5%。在冰溫貯藏過程中,Shewanellahafniensis和X1菌株的數量逐漸增加,到貨架期終點時兩者所占比例增至32%和27%,成為主要優勢菌。Acinetobacterjohnsonii、Acinetobacterbeijerinckii的數量雖呈下降趨勢,但到貨架期終點時,兩者所占比例仍達到16.5%和13%,成為次優菌株,Arthrobacterbergeri的數量也呈現下降趨勢,貨架期終點時所占比例僅為3%。Planoccoccuscitreus在初始菌相中所占比例為3.5%,在整個貯藏過程中比例變化很小。其余菌株在初始菌相中所占比例僅為9%,在貯藏期間各菌株數量逐漸減少,第8 d時Bacilluslicheniformis消失,到貨架期終點時,總體所占比例下降至5%。在整個貨架期內,Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii分別所占比例均高于其他菌種,說明它們在冰溫下的代謝繁殖能力仍高于其他菌種,成為南美白對蝦腐敗的主要菌種。
由圖4可知,冰溫貯藏貨架期內,隨著時間的延長,菌落總數呈增加趨勢,說明接種的各腐敗菌,在低溫下仍能利用蝦汁中的營養物質生長繁殖,但第4 d后因低溫長時間的作用,營養物質的消耗和代謝產物的積累,腐敗微生物繁殖速度減緩,菌落總數的增加速度變小。
綜上分析,冰溫貯藏可較好抑制細菌的生長繁殖,但不同菌株對冰溫冷害的敏感性不同,因此在貨架期內,耐低溫性較強的菌株數量所占比例逐漸增大,到貨架期終點,Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii成為數量優勢菌株。
2.4腐敗菌致腐能力測定
將接種各種腐敗細菌的無菌蝦汁置于冰溫條件下保藏,無菌蝦汁未接菌前測得的揮發性鹽基氮(TVB-N)為8.72 mg/100 g,利用考馬斯亮藍法,繪制出蛋白質標準曲線,求得蛋白質與吸光度的線性關系方程式y=14.758x+0.1805,相關系數R2=0.9972,未接菌前測得的蛋白質濃度為0.1063 mg/mL,以此為對照,將優勢腐敗菌分別接種無菌蝦汁,在貨架期結束(即第8 d)時測定蝦汁揮發性鹽基氮和蛋白質濃度,因在貨架期內揮發性鹽基氮呈上升趨勢,蛋白質濃度呈下降趨勢,所以用極差代表貨架期內的發展趨勢,并采用SPSS16.0軟件對極差進行多重比較,顯著性水平為p<0.01,結果見表3。

表3 冰溫條件下蛋白質濃度和揮發性鹽基氮的多重比較Table 3 Multiple comparisons of protein concentration and volatile base nitrogen during ice-temperature storage

圖4 南美白對蝦冰溫貯藏過程中 腐敗菌細菌總數的變化規律Fig.4 The change of the total number of spoilage bacteria of penaeus vannamei during ice-temperature storage
注:A,B,C,D,E,F,G,H,I為多重比較結果,同列標有不同大寫字母者表示組間差異極顯著(p<0.01),標有相同大寫字母者表示組間差異極不顯著(p>0.01)。
從表3可以看出,在冰溫條件下,數量占優勢的四種腐敗菌分解蝦汁產揮發性鹽基氮和降解蛋白質的能力較強,不僅與非優勢菌株相比均存在顯著差異,而且優勢菌株彼此之間同樣存在顯著差異,由此可以認為數量優勢菌株即為腐敗優勢菌株,各菌株腐敗能力的強弱依次為Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii。
為了進一步探討冰溫條件下,優勢腐敗菌引起鮮蝦腐敗的時間進程,在不同時間點取樣測定揮發性鹽基氮和蛋白質濃度,結果見圖5。

圖5 冰溫條件下揮發性鹽基氮含量和蛋白質濃度的變化Fig.5 The change of volatile basic nitrogen content and protein concentration during ice temperature storage
從圖5可以看出,各腐敗菌株降解蛋白質產揮發性氨基氮的時間進程基本一致,通過SPSS分析各菌株在同一時間內揮發性鹽基氮和蛋白質濃度的變化,發現冰溫貯藏第2 d接種各種優腐敗菌的蝦汁,其揮發性鹽基氮和蛋白質濃度彼此之間無差異,意味著致腐能力幾乎無差別。除此之外,各時間點的測定值均存在顯著差異(p<0.01),優勢腐敗菌產揮發性鹽基氮從高到低依次是:Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii,蛋白質濃度從低到高依次是Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii,進一步證明各菌株腐敗能力的強弱依次為Shewanellahafniensis、X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii。
另外通過三糖鐵培養基產硫化氫實驗,發現南美白對蝦冰溫貯藏過程中的12株腐敗細菌,僅有優勢腐敗菌Shewanellahafniensis在接種2 d后產生硫化氫,可見在冰溫貯藏南美白對蝦時,腐敗菌Shewanellahafniensis不僅數量占優勢,而且是形成硫化氫等不良腐敗氣味的關鍵菌株。
3 結論
許多研究表明細菌是引起對蝦腐敗變質的主要原因,本實驗以冰溫貯藏南美白對蝦為研究對象,在腐敗菌分離純化的基礎上,通過形態、生理生化特征測定和16S rRNA序列分析等多種方法,發現分離自冰溫貯藏南美白對蝦的12株腐敗細菌分別為Shewanellahafniensis、Acinetobacterjohnsonii、Planoccoccuscitreus、Bacilluscereus、Acinetobacterbeijerinckii、Enterobacterhormaechei、Arthrobacterbergeri、Bacilluslicheniformis,另外X1和X10菌株可能為潛在的新屬種。對在菌群中占有較高比例且致腐能力較強的優勢腐敗菌進行腐敗能力分析,發現冰溫條件下致腐能力最強的菌株是Shewanellahafniensis,其次依次是X1、Acinetobacterjohnsonii和Acinetobacterbeijerinckii。
本文通過對冰溫條件下南美白對蝦中優勢腐敗菌及其腐敗能力的分析,可以在實際貯藏過程中有針對性的抑制優勢腐敗菌的代謝活動,有效的降低其腐敗能力,從而延長南美白對蝦的貨架期。當然,傳統的微生物分離鑒定方法很難真實反映腐敗菌污染南美白對蝦的多樣性,不同特征的南美白對蝦對不同的腐敗菌相都會產生較大的動態響應,為了延長南美白對蝦的貨架期還需要進一步探索南美白對蝦內不同菌群之間的相互作用以及南美白對蝦內源酶等相互之間的影響。
[1]Nirmaln P,Beniakul S. Effect of ferulic acid on inhibition of polyphenoloxidase and quality changes of Pacific white shrimp(Litopenaeus vannamei)during iced storage[J]. Food Chemistry,2009,116(1):323-331.
[2]黎冬明,葉云花,劉成梅,等.冰溫技術在食品工業中的應用[J].食品科技,2006(1):32-34.
[3]凌萍華,謝晶,趙海鵬,等.冰溫貯藏對南美白對蝦保鮮效果的影響[J].江蘇農業學報,2010,26(4):828-832
[4]張娟,婁永江.冰溫技術及其在食品保鮮中的應用[J].食品研究與開發,2006,27(8):150-152.
[5]趙海鵬,謝 晶,嚴文蓉.南美白對蝦冷藏過程中的細菌分離初步鑒定及菌相分析[J].江蘇農業學報,2011,27(1):164-168.
[6]蘇永玲,謝晶. 南美白對蝦冷凍保鮮研究進展[J].安徽農業科學,2011,39(4):2286-2288.
[7]GB/T 4789.2-2008.食品微生物學檢驗—菌落總數測定[S].北京:中國標準出版社,2008.
[8]夏涵,府偉靈,陳鳴,等.快速提取細菌DNA方法的研究[J].現代預防醫學,2005,32(5):571-573.
[9]朱飛舟,陳利玉,陳漢春,等.16S rRNA基因序列分析法鑒定病原細菌[J].中南大學學報:醫學版,2013,38(10):1036.
[10]錢雪琴,張軍,沈芳,等.Chelex-100法和堿性裂解法提取細菌DNA的比較[J].中國衛生檢驗雜志,2008,18(8):1565.
[11]Chun J,Lee J H,Jung Y,et al. EzTaxon:a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences[J].International Journal of Systematic and Evolutionary Microbiology,2007,57(10):2259-2261.
[12]葉日英,孫力軍,王雅玲,等.凡納濱對蝦內源優勢腐敗菌的分離與鑒定[J]. 水產學報,2013,37(9):1426.
[13]李娟,張耀庭,曾偉,等.應用考馬斯亮藍法測定總蛋白含量[J]. 中國生物制品學雜志,2000,13(2):118-120.
[14]李學英,許鐘,楊憲時,等. 大黃魚腐敗菌腐敗能力分析與特定腐敗菌鑒別[J].上海海洋大學學報,2010,19(4):547-552.
[15]許振偉.冷藏魚類腐敗菌腐敗能力分析[D].上海:上海海洋大學,2011.
[16]陳培基,李劉冬,黃珂.自動定氮儀測定水產品中的揮發性鹽基氮[J].南方水產,2005,1(6):50-54.
[17]Santlagoa P,Aubour G,Carmen G,et al. Quality assessment of sardines during storage by measurement of fluorescent compounds[J].Journal of Food Science,1997,62(2):295-299.
[18]張娟,婁永江.冰溫技術及其在食品保鮮中的應用[J].食品研究與開發,2006,27(8):150-152.
[19]楊霞,陳陸,王川慶,等. 16S rRNA基因序列分析技術在細菌分類中應用的研究進展[J].西北農林科技大學學報,2008,36(2):56-58.
Isolation,identification and spoilage ability analysis on the particular spoilage bacteria of penaeus vannamei in the ice temperature storage
XIE Li-dan,LI Lei-lei,WANG Su-ying*,DONG Shi-rui
(Tianjin Key Laboratory of Food Biotechnology,College of Biotechnology and Food Science,Tianjin University of Commerce,Tianjin 300400,China)
In order to isolate and identify penaeus vannamei,s spoilage bacteria and measure their spoilage ability in ice temperature storage conditions,the freezing point of penaeus vannamei was determined based on freezing experiment and the spoilage bacteria were separated from penaeus vannamei which was stored under the temperature between 0 ℃and the freezing point,the isolated microbes were identified by morphological observation and molecular biology methods. The separated spoilage bacteria were inoculated into sterilized shrimp juice,hydrogen sulfide,degradated proteins and volatile base nitrogen(TVB-N)were selected to evaluate the spoilage ability. The results showed that 12 strains of spoilage bacteria were isolated from penaeus vannamei,they wereShewanellahafniensis,Acinetobacterjohnsonii,Planoccoccuscitreus,Bacilluscereus,Acinetobacterbeijerinckii,Enterobacterhormaechei,Arthrobacterbergeri,Bacilluslicheniformis,of which X1 and X10 may be a potential new species or genus. In ice temperature storage conditions,dominant spoilage bacteria leading to the corruption of penaeus vannamei wereShewanellahafniensis,X1,AcinetobacterjohnsoniiandAcinetobacterbeijerinckii.
penaeus vannamei;ice temperature storage;dominant spoilage bacteria;isolation and identification;spoilage ability
2015-07-09
謝麗丹(1992-),女,碩士研究生,研究方向:微生物資源的前期開發,E-mail:13820159533@163.com。
王素英(1964-),女,博士,教授,研究方向:微生物資源的前期開發和利用,E-mail:wsying@tjcu.edu.cn。
國家自然科學基金項目(31270050);天津市高等學校創新團隊(TD12-5049)。
TS254.4
A
1002-0306(2016)05-0171-06
10.13386/j.issn1002-0306.2016.05.025
