海拔對全緣葉綠絨蒿植株性狀和花特征的表型選擇分析
2016-09-02 02:41:37張霓雯劉光立
西北植物學(xué)報(bào) 2016年7期
吳 云, 張霓雯, 彭 瀚, 劉光立*
?
海拔對全緣葉綠絨蒿植株性狀和花特征的表型選擇分析
吳云1,2,3, 張霓雯4, 彭瀚1, 劉光立1*
(1 四川農(nóng)業(yè)大學(xué) 風(fēng)景園林學(xué)院,成都 611130;2 中國科學(xué)院西雙版納熱帶植物園 熱帶森林生態(tài)學(xué)重點(diǎn)實(shí)驗(yàn)室,云南勐臘 666303;3 中國科學(xué)院大學(xué),北京 100039; 4 四川航天職業(yè)技術(shù)學(xué)院 四川廣漢 618300)
為了研究海拔差異對植株性狀、花特征表型選擇的影響,以青藏高原高寒草甸的全緣葉綠絨蒿(Meconopsisintegrifolia)為研究材料,于盛花期內(nèi),測定不同海拔(4 452、4 081和3 681 m)種群中個(gè)體植株性狀、花特征、單果結(jié)實(shí)數(shù)并進(jìn)行統(tǒng)計(jì)分析,采用線性回歸模型估計(jì)不同海拔種群間植株性狀、花特征所受的表型選擇(選擇差與選擇梯度)。結(jié)果表明:(1)隨著海拔升高,全緣葉綠絨蒿植株性狀、花特征及單果結(jié)實(shí)數(shù)顯著降低,海拔越高的種群中株高越矮、葉面積越小、花數(shù)越少、花越小、單果結(jié)實(shí)數(shù)越低。(2)不同海拔種群中各性狀的表型選擇存在差異,較低海拔(3 681 m)種群中花數(shù)、花大小具有顯著的選擇差和選擇梯度,表現(xiàn)為花越多、花越大的個(gè)體雌性適合度越高;海拔較高(4 081 m)的種群中株高、葉面積及花數(shù)更容易受到選擇,表現(xiàn)為植株越高、葉面積越大、花越多的個(gè)體雌性適合度越高;海拔最高(4 452 m)的種群中葉面積與花數(shù)的選擇梯度接近顯著。(3)植物性狀分化伴隨著海拔的變化而呈現(xiàn)出差異,較低海拔種群中花特征容易受到選擇,而較高海拔種群中可能由于傳粉者稀少、資源限制等因素使得株高、葉面積更容易受到選擇。
株高;葉面積;花特征;雌性適合度;表型選擇;海拔
自然界中,植物的固著生長模式?jīng)Q定了其必須依賴一定的傳粉媒介(動(dòng)物、水、風(fēng)等)才能實(shí)現(xiàn)有性生殖,而這其中傳粉昆蟲作為最主要、最多樣的傳粉媒介,往往對植物的花部性狀及進(jìn)化具有重要的作用,并最終影響植物的雌雄適合度[1]。種群內(nèi)或種群間,不同傳粉者的訪花偏好往往造成花部性狀的適應(yīng)性分化,并最終導(dǎo)致個(gè)體適合度的改變[2-3]。如對毛茛狀金蓮花(Trolliusranunculoides)的研究表明,蜜蜂和蠅類偏好較大的花,在蠅類為主導(dǎo)傳粉者的種群中,個(gè)體具有更多的單花花瓣但更小的花,繼而造成毛茛狀金蓮花種群間結(jié)實(shí)率存在差異[4]。在一些高度特化的傳粉系統(tǒng)中,傳粉者與植物往往是一對一的關(guān)系。如,在南美厄瓜多爾安第斯山脈,作為加長鐘形花(Centropogonnigricans)的唯一的傳粉者,長舌蝙蝠(Anourafistulata)的舌頭長度幾乎與加長鐘形花花冠的長度一致(8~9 cm),造成加長鐘形花具有更長的雌性與雄性持續(xù)時(shí)間[5]。對分布于尼泊爾的象牙參(Roscoeapurpurea)的研究發(fā)現(xiàn),一種虻(Philolichelongirostris)的喙長與此種植物的花冠管長相吻合,作為此種象牙參最有效的傳粉者對植物有性繁殖的成功具有重要的作用[6]。
具有專化傳粉系統(tǒng)的傳粉者-宿主植物間的關(guān)系相對穩(wěn)定,但自然界中更多的是泛化傳粉系統(tǒng),且這種由泛化傳粉者介導(dǎo)對植株性狀、花特征的選擇往往存在時(shí)間和空間上的變異。如對多年生草本植物春苦豆(Lathyrusvernus)[7]和毛茛狀金蓮花(T.ranunculoides)[8]傳粉者介導(dǎo)的表型選擇研究表明,不同花期,訪花昆蟲的變化可能對植物花特征(花數(shù)量、花瓣及花萼大小)的分化起到了至關(guān)重要的作用,繼而影響了不同花期植株的適合度。對牛皮杜鵑(Rhododendronaureum)的深入研究表明,早花期與末花期傳粉者介導(dǎo)表型選擇的差異不但造成植株適合度的差異,而且對種子遺傳多樣性亦產(chǎn)生影響[9]。
值得注意的是,傳粉者介導(dǎo)的選擇作用更多表現(xiàn)為空間上的差異。如,對藍(lán)花耬斗菜(Aquilegiacoerulea)[10]、蘭科植物Gymnadeniaodoratissima[11]以及挪威Dovrefjell 國家公園中的植物[12]研究均表明,不同海拔間傳粉者種類的差異導(dǎo)致植物個(gè)體的花色存在差異。除對個(gè)體花色具有選擇作用外,傳粉者的差異亦對個(gè)體的花大小、雌雄蕊位置關(guān)系等具有各異的選擇作用。如,對毛茛(Ranunculusacris)的研究表明,低海拔種群中傳粉者偏好更大的花,高海拔種群中傳粉者無此偏好[13]。對蘭科植物Gymnadeniaodoratissima的研究亦有相似的結(jié)論[11]。有趣的是[14-15],由于高海拔種群中傳粉者的缺乏,植物雌雄蕊位置更為接近,有自交的傾向。隨著海拔的變化,傳粉者的種類、數(shù)量、組成、活動(dòng)能力、訪花頻率均會出現(xiàn)變化,這種變化所帶來的表型選擇作用于不同海拔種群可能不同,而目前大多數(shù)的研究均是集中于不同海拔種群間個(gè)體的形態(tài)指標(biāo),對于傳粉者差異所帶來的對特定性狀的選擇差、選擇梯度,繼而影響個(gè)體適合度的認(rèn)識較為缺乏。
本研究以傳粉昆蟲泛化的全緣葉綠絨蒿(Meconopsisintegrifolia)為研究對象,通過對不同海拔種群個(gè)體的植株性狀、花特征、雌性適合度及相關(guān)的表型選擇進(jìn)行分析,主要探討以下問題:(1)在不同海拔種群中,全緣葉綠絨蒿的植株性狀、花特征及雌性適合度是否存在差異?(2)通過相對雌性適合度在不同海拔種群估計(jì)的表型選擇是否有差異?
1 材料和方法
1.1研究材料及研究地點(diǎn)
全緣葉綠絨蒿為罌粟科綠絨蒿屬一年生或多年生草本,主要分布于海拔2 500 m的林緣到海拔5 000 m的雪線區(qū)間,生境多為高山草甸。植株高15~150 cm,花黃色,通常3~5朵生于莖上部葉腋內(nèi);雄蕊多數(shù);心皮3~6,子房卵形;蒴果。花期5~8月,果期9~10月。
研究于2014年6月中旬(盛花期)在四川省阿壩藏族羌族自治州汶川縣臥龍自然保護(hù)區(qū)巴郎山(102.90°~102.95°E, 30.88°~30.91°N,海拔4 484 m)進(jìn)行。巴郎山年均溫8.6 ℃(1月均溫-1.7 ℃,7月均溫17.0 ℃),年降水量700~900 mm,雨季5~9月。在充分踏查的基礎(chǔ)上,共選擇了3個(gè)全緣葉綠絨蒿研究種群[種群1(4 452 m)、種群2(4 081 m)、種群3(3 681 m)]。
1.2研究方法
1.2.1性狀指標(biāo)2014年6月中旬(盛花期)于各種群隨機(jī)標(biāo)記30株全緣葉綠絨蒿成熟植株,測量并統(tǒng)計(jì)株高、葉片數(shù)、葉片長寬(每株隨機(jī)測量3個(gè)葉片)、花數(shù)、花瓣數(shù)(每株隨機(jī)標(biāo)記3朵花)、花瓣長寬(隨機(jī)測量標(biāo)記的3朵花的花瓣,每朵花測量3個(gè)花瓣)。其中,株高的測量標(biāo)準(zhǔn)為貼近地面部分到最高點(diǎn)的高度,葉片長寬與花瓣長寬的測量方法見圖1。待果實(shí)成熟尚未開裂前,每株隨機(jī)收集3個(gè)成熟果實(shí),統(tǒng)計(jì)種子數(shù)。
1.2.2替代指標(biāo)及替代方法為減小相似指標(biāo)對分析結(jié)果的干擾,分別以葉面積和花大小作為替代指標(biāo)加以分析。具體替代如下:1)葉面積= 葉片數(shù)×葉片長×葉片寬;2)花大小= 花瓣數(shù)×花瓣長×花瓣寬。分別計(jì)算每株葉面積的平均值、花大小的平均值、單果結(jié)實(shí)數(shù)的平均值,并以平均值作為該株相應(yīng)指標(biāo)的指標(biāo)值。
1.2.3回歸模型表型選擇分析中,參照Lande等[16]的模型進(jìn)行分析。
選擇差:選擇差表示每個(gè)性狀所受選擇壓力的
總和,包括直接選擇和間接選擇。以相對雌性適合度(個(gè)體種子數(shù)/種群平均種子數(shù))作為因變量,標(biāo)準(zhǔn)化的性狀指標(biāo)(平均值=0,方差=1)為自變量。通過相對雌性適合度與每個(gè)標(biāo)準(zhǔn)化的性狀指標(biāo)作一元線性回歸,所得的回歸系數(shù)“β”為選擇差。
選擇梯度:選擇梯度表示每個(gè)性狀所受的直接選擇壓力。以相對雌性適合度(個(gè)體種子數(shù)/種群平均種子數(shù))為因變量,標(biāo)準(zhǔn)化的性狀指標(biāo)(平均值=0,方差=1)為自變量,采用多元線性回歸,所得到的每個(gè)標(biāo)準(zhǔn)化性狀的偏回歸系數(shù)“βi”,即為線性選擇梯度[17-18]。
1.3數(shù)據(jù)分析
采用單因素方差分析(one-way ANOVA)檢驗(yàn)不同海拔種群間全緣葉綠絨蒿株高、葉面積、花數(shù)、花大小、單果結(jié)實(shí)數(shù)的差異并計(jì)算變異系數(shù)(CV)。采用Spearman相關(guān)性分析檢驗(yàn)株高、葉面積、花數(shù)、花大小間的相關(guān)性,相關(guān)分析數(shù)據(jù)采用3個(gè)種群的數(shù)據(jù)。
以相對雌性適合度為因變量,標(biāo)準(zhǔn)化的性狀為自變量分別進(jìn)行一元與多元線性回歸,分別求出選擇差與選擇梯度。以顯著的選擇差對應(yīng)的標(biāo)準(zhǔn)化性狀與相對雌性適合度為變量作圖。以上分析均在SPSS 19.0中進(jìn)行,圖在Excel 2007中完成。
2 結(jié)果與分析
2.1不同海拔種群全緣葉綠絨蒿植株性狀、花特征及其相關(guān)性
表1顯示,全緣葉綠絨蒿株高、葉面積、花數(shù)、花大小及種子產(chǎn)量在不同海拔種群間均存在顯著差異(P<0.001)。海拔越高的種群,個(gè)體的株高越低,葉面積越小,花數(shù)越少,花越小,種子產(chǎn)量越低。
各海拔種群中個(gè)體株高、葉面積、花數(shù)、花大小及種子產(chǎn)量均存在不同程度的差異。其中,種群中個(gè)體的葉面積、花數(shù)、花大小的變異較大,變異系數(shù)較高(表2)。隨著海拔升高,性狀的差異逐漸增大,海拔最高的種群1中個(gè)體的株高、葉面積、花大小和種子產(chǎn)量變異最大。
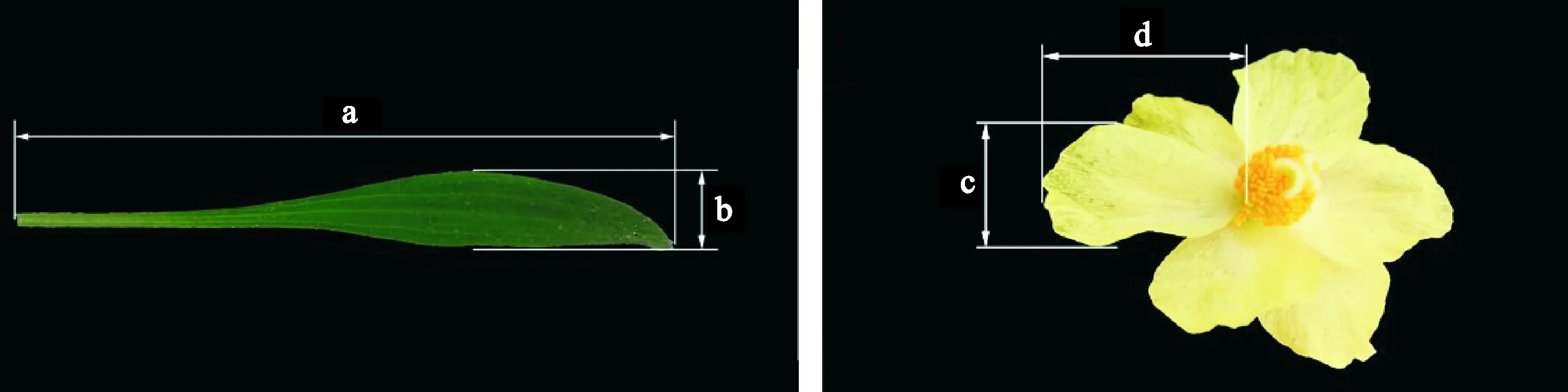
a.葉片長;b.葉片寬;c.花瓣寬;d.花瓣長圖1 全緣葉綠絨蒿葉片長寬和花瓣長寬測量示意圖a. Leaf length; b. Leaf width; c. Petal width; d. Petal lengthFig. 1 Measurements on leaf length, leaf width, petal length and petal width of Meconopsis integrifolia
Spearman相關(guān)性分析表明(表3),株高、葉面積、花數(shù)及花大小間均存在極顯著正相關(guān)性(P<0.01)。植株越高,葉面積越大,花越多,花越大。
2.2不同海拔全緣葉綠絨蒿的表型選擇分析
通過相對雌性適合度與各標(biāo)準(zhǔn)化的性狀指標(biāo)間線性回歸分析結(jié)果發(fā)現(xiàn),種群1中個(gè)體的各性狀沒有顯著的選擇差和選擇梯度。由表4表5可以看出,種群2中個(gè)體的株高(β=0.135,P<0.001)、葉面積(β=0.108,P<0.01)和花數(shù)(β=0.079,P=0.014)存在顯著的選擇差,且種群2中個(gè)體的株高(βi=0.115,P<0.001)與花數(shù)(βi=0.056,P=0.013)亦存在顯著的選擇梯度。種群3中個(gè)體的花數(shù)與花大小存在顯著的選擇差(β=0.066,P=0.023;β=0.086,P=0.011)和選擇梯度(βi=0.103,P=0.02;βi=0.09,P=0.015)。圖2表明,種群2中株高、葉面積、花數(shù)與適合度間存在顯著的正定向選擇,表現(xiàn)為植株越高、葉面積越大、花越多,個(gè)體的適合度越大。圖3表明,種群3中花數(shù)、花大小與適合度間存在顯著的正定向選擇,表現(xiàn)為花越多、花越大,個(gè)體適合度越大。

表3 全緣葉綠絨蒿各性狀間的相關(guān)性
注:**表示顯著水平P<0.01
Notes: ** indicated significant levelP<0.01
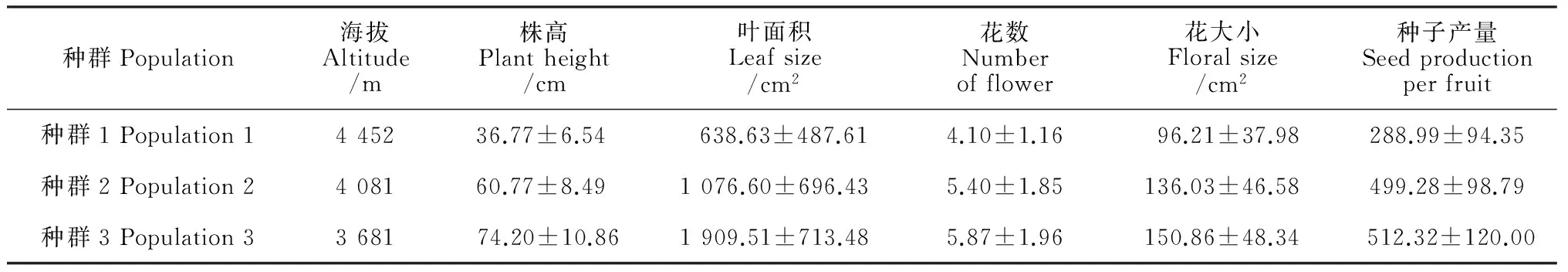
表1 不同海拔種群全緣葉綠絨蒿植株性狀、花特征及種子產(chǎn)量
注:表中數(shù)據(jù)為平均值±標(biāo)準(zhǔn)差。
Notes: data= mean±S.D.
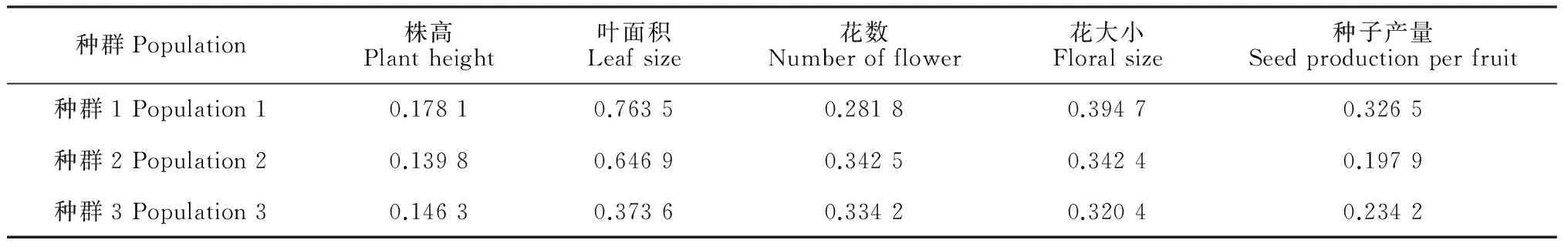
表2 不同海拔種群全緣葉綠絨蒿性狀變異系數(shù)
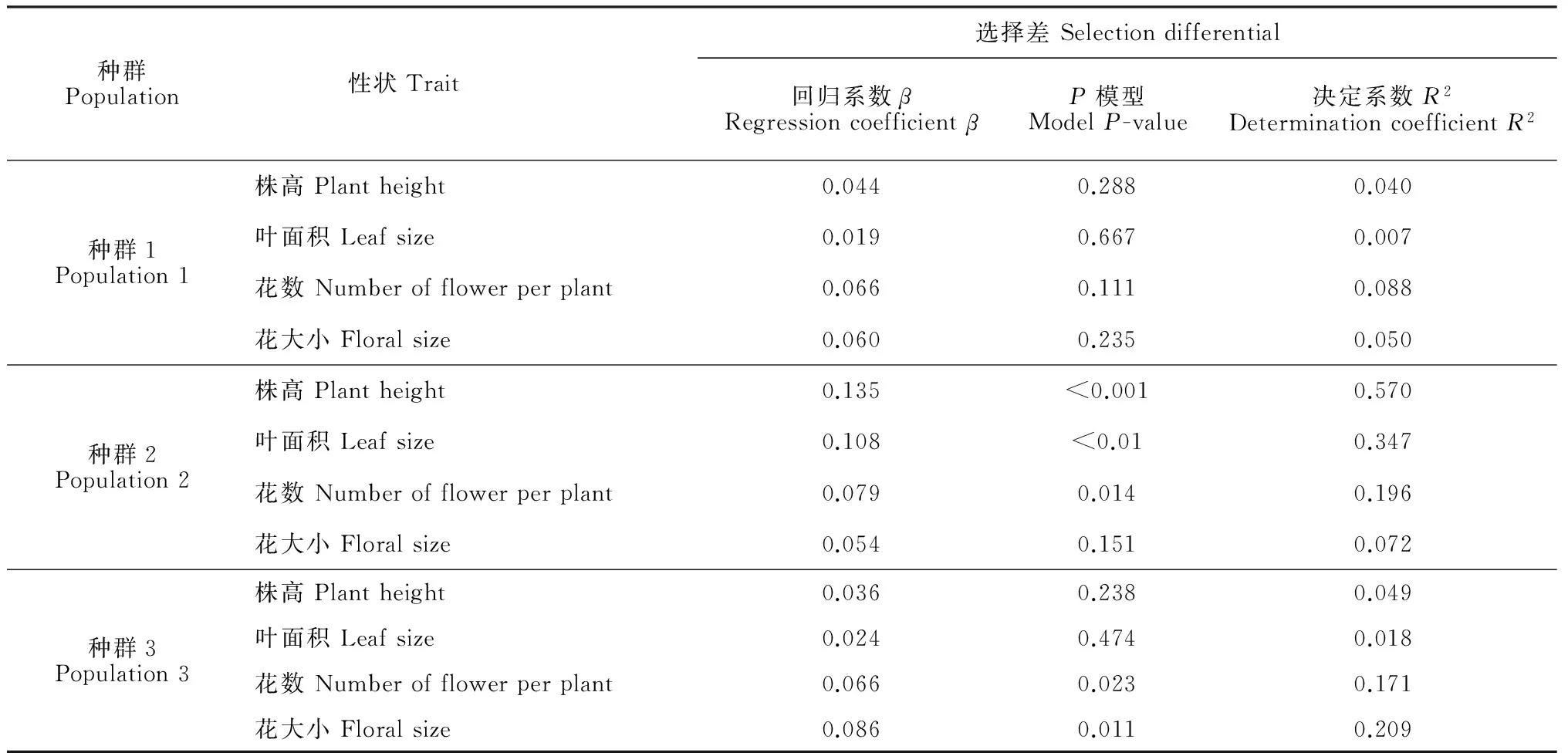
表4 全緣葉綠絨蒿各性狀在不同海拔種群間受到的選擇差
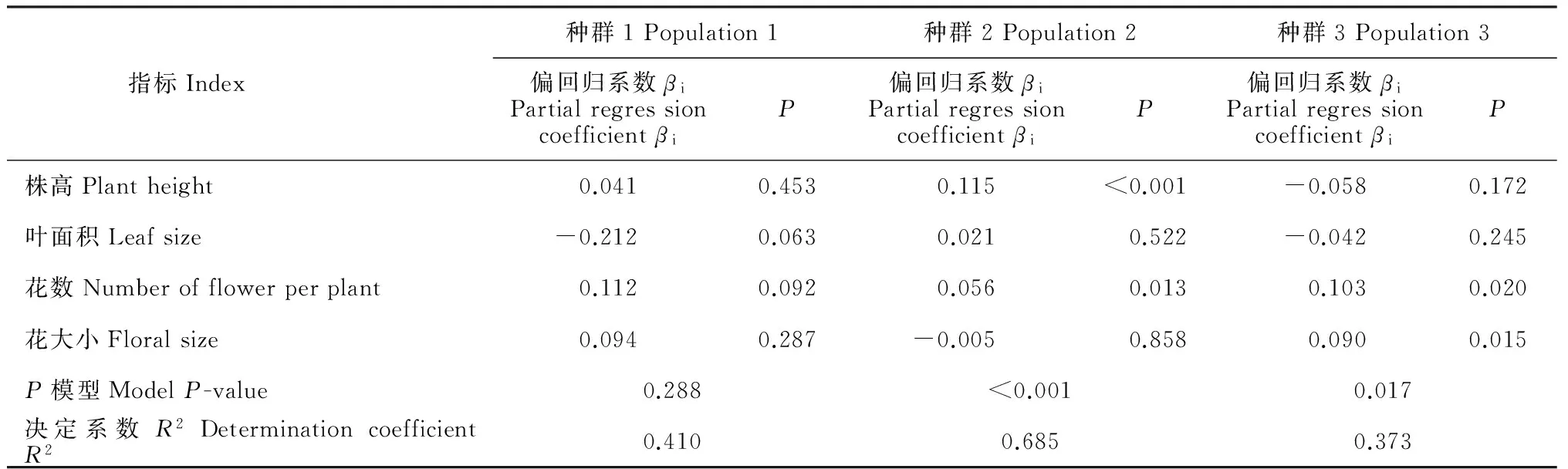
表5 全緣葉綠絨蒿各性狀在不同海拔種群間的選擇梯度
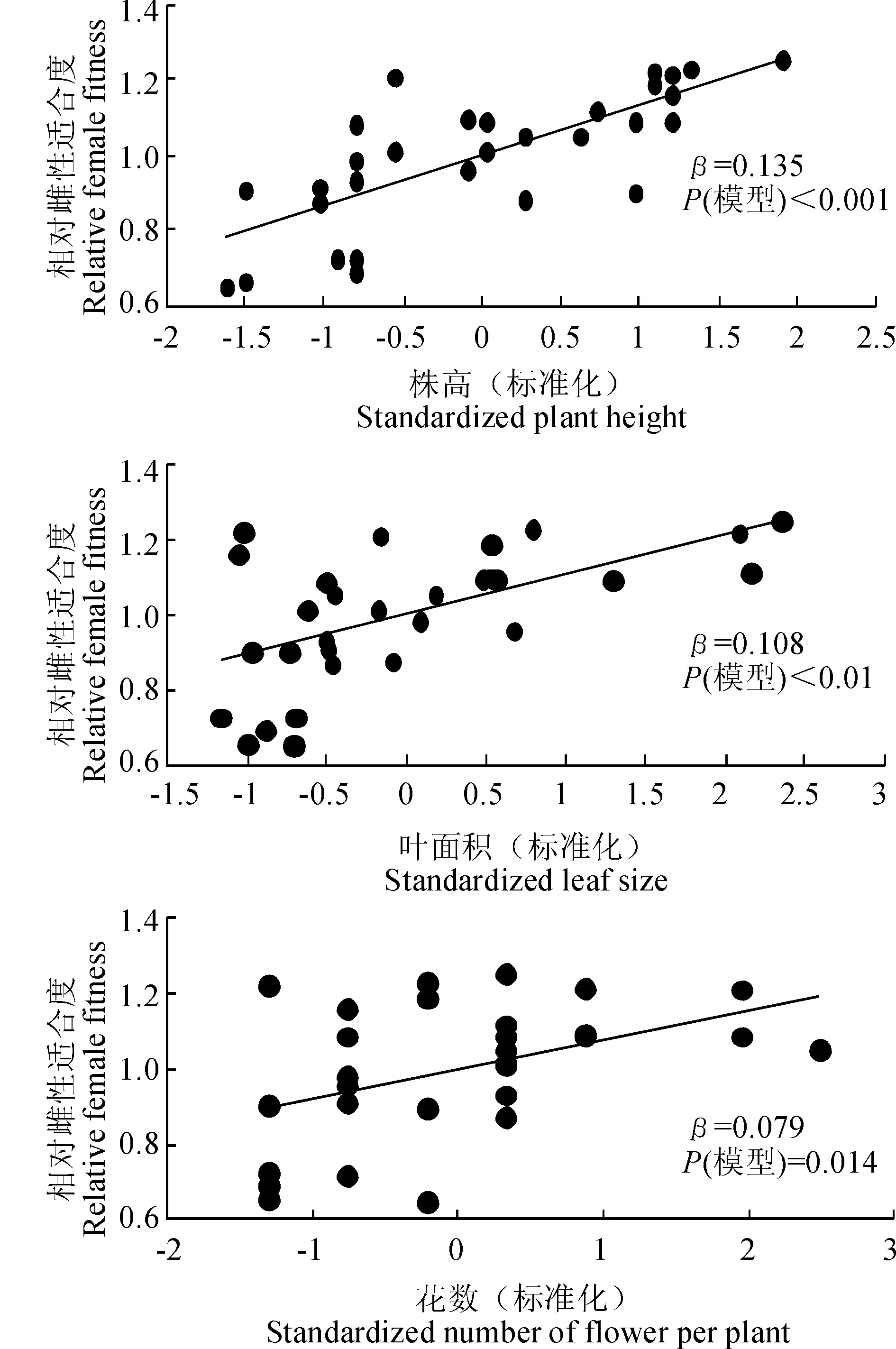
圖2 種群2全緣葉綠絨蒿相對雌性適合度與具有顯著選擇差的性狀的關(guān)系Fig. 2 Relationships between relative female fitness and traits with significant selection differential in population 2 of M. integrifolia
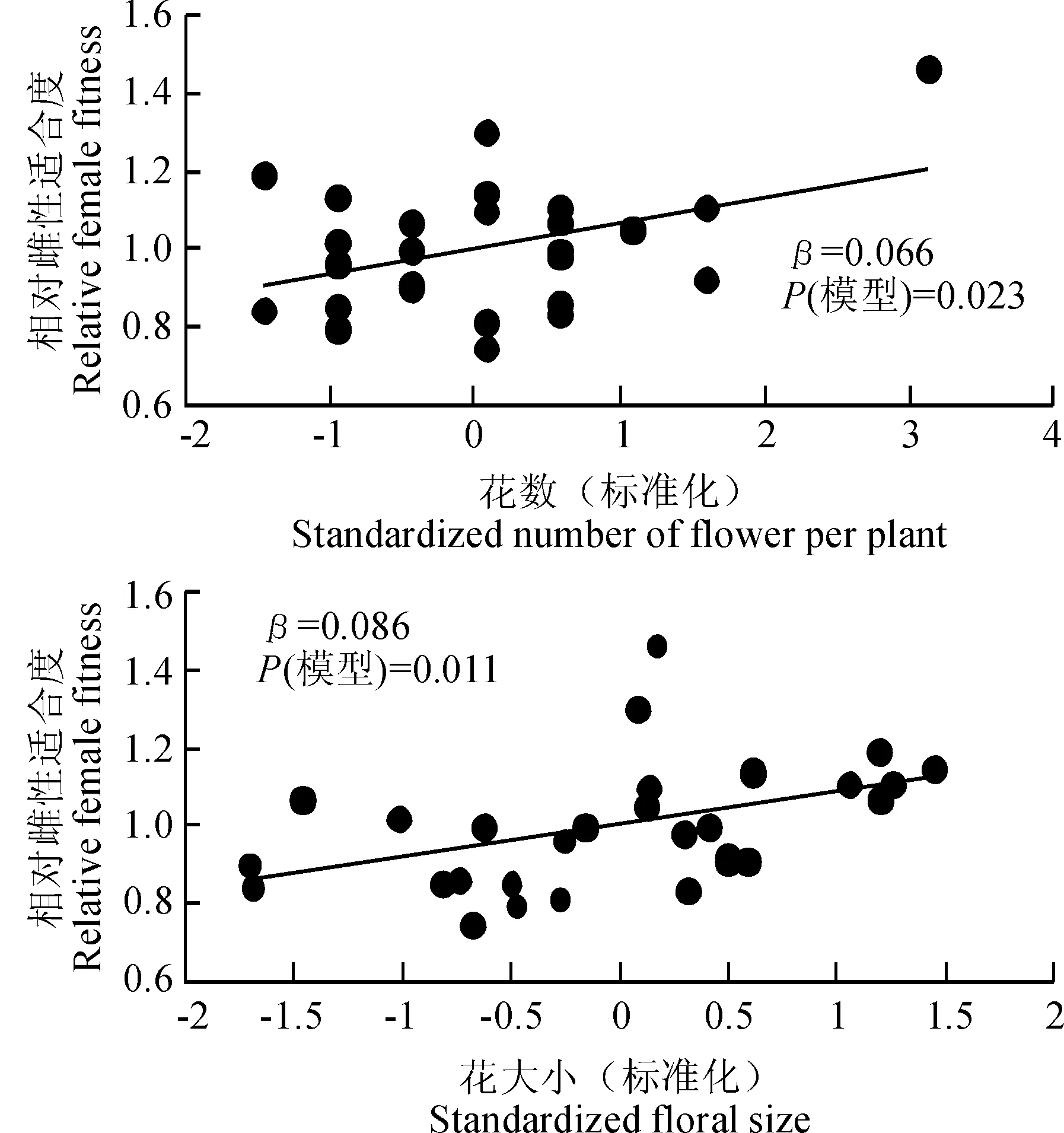
圖3 種群3全緣葉綠絨蒿相對雌性適合度與具有顯著選擇差的性狀的關(guān)系Fig. 3 Relationships between relative female fitness and traits with significant selection differential in population 3 of M. integrifolia.
3 討 論
3.1不同海拔植株性狀、花特征的變化
全緣葉綠絨蒿的株高、葉面積、花數(shù)、花大小及單果結(jié)實(shí)數(shù)在不同海拔種群間均存在顯著差異。海拔越高的種群中株高越矮、葉面積越小、花數(shù)越少、花越小、單果結(jié)實(shí)數(shù)越低。高山植物中,營養(yǎng)器官的生物量和資源投入會隨著海拔的升高而呈現(xiàn)出逐漸降低的趨勢,使得株高、葉面積等性狀呈現(xiàn)下降[19-21]。由于資源限制的存在,隨著海拔的升高,大多數(shù)高山植物會選擇減少對營養(yǎng)器官、繁殖器官絕對資源量的投入,但會增加對繁殖器官相對資源量的投入,即隨著海拔升高,繁殖器官生物量的比重會增加。在很多植物中,花特征如花數(shù)、花色、花大小、氣味、花蜜、雌雄蕊位置關(guān)系、開花物候等受到傳粉者的選擇作用[11,18,22-23]。如,對花荵科植物膠粘花荵(Polemoniumviscosum)的研究表明,該植物花序高度、花大小隨著海拔的變化可能是由傳粉者介導(dǎo)的選擇作用造成的[24]。對挪威Dovrefjell國家公園的高山植物的研究發(fā)現(xiàn),隨著海拔升高,傳粉者由以蜂類為主轉(zhuǎn)變?yōu)橐韵夘悶橹鳎参锏幕ㄉ灿傻秃0蔚腷ee-blue和UV-blue轉(zhuǎn)變?yōu)楦吆0蔚腷ee-blue-green和green[12]。而對耬斗菜屬植物Aquilegiacoerulea連續(xù)4年的研究發(fā)現(xiàn),海拔越高,植物花色越藍(lán);相對于僅有大戟天蛾(Hyleslineata)傳粉的種群,在另一種天蛾傳粉者Sphinxvashti為主導(dǎo)的種群中個(gè)體具有更長的距[10]。不同海拔種群中全緣葉綠絨蒿傳粉者的數(shù)量、傳粉效率存在差異,隨著海拔升高,傳粉者數(shù)量、傳粉效率大致呈現(xiàn)降低的趨勢[25],加之可能存在的資源限制,造成不同海拔種群間株高、葉面積及花特征的分化。作為一種“溫室植物”,全緣葉綠絨蒿更多的采用花內(nèi)溫度的升高以吸引傳粉者,作為泛化傳粉系統(tǒng)的物種,高海拔種群中的低溫可能使得此物種增加對升溫機(jī)制的選擇。因此,對于全緣葉綠絨蒿而言,不同海拔種群間的溫度、傳粉者的數(shù)量、傳粉效率以及資源限制可能共同造成其植株性狀、花特征在空間上的分化。
3.2不同海拔表型選擇的差異
通過相對雌性適合度估計(jì)發(fā)現(xiàn),不同海拔種群中全緣葉綠絨蒿植株性狀及花特征受到的表型選擇存在差異。海拔較低的種群3中,花數(shù)和花大小受到顯著的正定向選擇差和選擇梯度;海拔更高的種群2中,株高和花數(shù)受到顯著的正定向選擇差和選擇梯度,且葉面積亦受到顯著的正定向選擇差;海拔最高的種群1中,各性狀未受到顯著的正定向選擇差,但葉面積和花數(shù)受到的正定向選擇梯度接近顯著。傳粉者選擇具有更多的、更大的花的個(gè)體,可能是因?yàn)楦嗟幕ā⒏蟮幕ㄒ馕吨鼜?qiáng)的吸引力和更多的訪花報(bào)酬[26-27],較低海拔種群中,全緣葉綠絨蒿不是優(yōu)勢種且往往與其他草本植物同時(shí)開放,在傳粉者介導(dǎo)的選擇作用下,此類植物采取提高花展示的方式以吸引傳粉者,繼而維持一定的適合度利益。隨著海拔的升高,全緣葉綠絨蒿逐漸成為群落中的優(yōu)勢種,花展示較為突出,此時(shí)吸引傳粉者成為次要目標(biāo),由于資源限制和各器官間資源分配權(quán)衡效應(yīng)的存在[20-21],保障種子的發(fā)育成為主要目標(biāo),因此傳粉者介導(dǎo)的表型選擇更多的對株高、葉面積這類獲取資源的性狀具有選擇作用。
植物的株高、葉面積及花特征不僅在空間上存在分化,亦在時(shí)間上存在分化。同時(shí),導(dǎo)致植物花特征的因素眾多,既包括非生物因素中的溫度、光照、水分、資源有限性及分配模式等,又包括生物因素中的傳粉者、盜蜜者、植食者、傳粉限制等。在以后的研究中,應(yīng)該弄清傳粉者介導(dǎo)花特征的選擇是否受到花粉限制的影響,不同傳粉者對某一特定花特征的選擇強(qiáng)度如何,以及不同年際間、同一生長季內(nèi)不同花期間傳粉者介導(dǎo)花特征的選擇作用如何,并更加全面地考慮各花特征及引起花特征分化的各類因素。
[1]STEBBINS G L. Adaptive radiation of reproductive characteristics in angiosperms. I. Pollination mechanism [J].AnnualReviewofEcologyandSystematics, 1970, 1: 307-326.
[2]SCHIESTL F P, PEAKALL R, MANT J G,etal. The chemistry of sexual deception in an orchid-wasp pollination system[J].Science, 2003, 302: 437-438.
[3]PARRA-TABLA V, VARGAS C F. Flowering synchrony and floral display size affect pollination success in a deceit-pollinated tropical orchid[J].ActaOecologica, 2007, 32: 26-35.
[4]ZHAO Z G, HUANG S Q. Differentiation of floral traits associated with pollinator preference in a generalist-pollinated herb,Trolliusranunculoides(Ranunculaceae)[J].InternationalJournalofPlantSciences, 2013, 174(4): 637-646.
[5]MUCHHALA N, THOMSON J D. Going to great length: selection for long corolla tubes in an extremely specialized bat-flower mutualism[J].ProceedingoftheRoyalSocietyB:BiologicalSciences, 2009, 276: 2 147-2 152.
[6]BABU R P, SHRESTHA M, DYER A G,etal. Out of Africa: evidence of the obligate mutualism between long corolla tubed plant and long-tongued fly in the Himalayas[J].EcologyandEvolution, 2015, 22(5): 5 240-5 251.
[7]EHRLéN J, MüNZBERGOVZ. Timing of flowering: opposed selection on different fitness components and trait covariation[J].TheAmericanNaturalist, 2009, 173(6): 819-830.
[8]劉樂樂, 劉左軍, 杜國禎, 等. 毛茛狀金蓮花不同花期的花特征和訪花昆蟲的變化及表型選擇[J]. 生物多樣性, 2012, 20(3): 317-323.
LIU L L, LIU Z J, DU G Z,etal. Floral traits, pollinator assemblages, and phenotypic selection at different flowering time forTrolliusranunculoides[J].BiodiversityScience, 2012, 20(3): 317-323.
[9]HIRAO A S, KAMEYAMA Y, OHARA M,etal. Seasonal changes in pollinator activity influence pollen dispersal and seed production of the alpine shrubRhododendronaureum(Ericaceae)[J].MolecularEcology, 2006, 15: 1 165-1 173.
[10]BRUNET J. Pollinators of the Rocky Mountain columbine: temporal variation, functional groups and associations with floral trait [J].AnnalsofBotany, 2009, 103: 1 567-1 578.
[11]SUN M, GROSS K, SCHIESTL F P. Floral adaptation to local pollinator guilds in a terrestrial orchid[J].AnnalsofBotany, 2014, 113: 289-300.
[12]ARNOLD S E J, SAVOLAINEN V, CHITTKA L. Flower colours along an alpine gradients, seen through the eyes of fly and bee pollinators[J].Arthropod-PlantInteraction, 2009, 3: 27-43.
[13]TOTLAND?. Environment-dependent pollen limitation and selection on floral traits in an alpine species[J].Ecology, 2001, 82(8): 2 233-2 244.
[14]TOTLAND?. Breeding system, insect flower visitation and floral traits of two alpine Cerastium species in Norway[J].Arctic,Antarctic,andAlpineResearch, 2003, 35(2): 242-247.
[15]JACQUEMYN H, MICHENEAU C, ROBERTS D L,etal. Elevational gradients of species diversity, breeding system and floral traits of orchid on Réunion island[J].JournalofBiogeography, 2005, 32: 1 751-1 761.
[16]LANDE R, ARNOLD S J. The measurement of selection on correlated characters[J].Evolution, 1983, 54: 112-123.
[17]STINCHCOMBE J R, AGRAWAL A F, HOHENLOHE P A,etal. Estimating nonlinear selection gradients using quadratic regression coefficients: double or nothing[J].Evolution, 2008, 62(9): 2 435-2 440.
[18]CHAPURLAT E, ?GREN J, SLETVOLD N. Spatial variation in pollinator-mediated selection on phenology, floral display and spur length in the orchidGymnadeniaconopsea[J].NewPhytologist, 2015, 208: 1 264-1 275.
[19]FABBRO T, K?RNER C. Altitudinal differences in flower traits and reproductive allocation[J].Flora, 2004, 199: 70-81.
[20]ZHAO Z G, DU G Z, ZHOU X H,etal. Variations with altitude in reproductive traits and resource allocation of three Tibetan species of Ranunculaceae[J].AustralianJournalofBotany, 2006, 54: 691-700.
[21]MILLA R, REICH P B. Multi-trait interactions, not phylogeny, fine-tune leaf size reduction with increasing altitude[J].AnnalsofBotany, 2011, 107: 455-465.
[23]HOPKINS R, RAUSHER M D. Pollinator-mediated selection on flower color allele drives reinforcement[J].Science, 2012, 335: 1 090-1 092.
[24]GALEN C. Measuring pollinator-mediated selection on morphometric floral traits: bumblebees and the alpine sky pilot,Polemoniumviscosum[J].Evolution, 1989, 43: 882-890.
[25]吳 云, 劉玉蓉, 彭 瀚, 等. 高山植物全緣葉綠絨蒿在不同海拔地區(qū)的傳粉生態(tài)學(xué)研究[J]. 植物生態(tài)學(xué)報(bào), 2015, 39(1): 1-13.
WU Y, LIU Y R, PENG H,etal. Pollination ecology of alpine herbMeconopsisintegrifoliaat different altitudes[J].ChineseJournalofPlantEcology, 2015, 39(1): 1-13.
[26]BLARER A, KEASAR T, SHMIDA A. Possible mechanisms for the formation of flowering size preferences by foraging bumblebees[J].Ethology, 2002, 108: 341-351.
[27]HAEDER L D, JOHNSON S D. Darwin’s beautiful contrivances: evolutionary and functional evidence for floral adaptation[J].NewPhytologist, 2009, 183: 530-545.
(編輯:潘新社)
Phenotypic Selection on Plant Traits and Floral Traits at Different Altitudes forMeconopsisintegrifolia
WU Yun1,2,3, ZHANG Niwen4, PENG Han1, LIU Guangli1*
(1 College of Landscape Architecture, Sichuan Agriculture University, Chengdu 611130, China; 2 Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan 666303, China; 3 University of Chinese Academy of Sciences, Beijing 100039, China; 4 Sichuan Aerospace Vocational College, Guanghan Sichuan 618300, China)
We measured plant traits and floral traits ofMeconopsisintegrifoliaat different altitude populations (4 452 m, 4 081 m, 3 681 m), collected ripe fruits in the end of flowering and counted seed production per fruit to estimate the overall phenotypic selection differential and selection gradient. We used one-way ANOVA analysis to estimate variations of plant traits, floral traits and seed production per fruit at different altitude populations. We evaluated phenotypic selection (selection differential and selection gradient) on plant traits (plant height and leaf size), floral traits at different altitudes populations using multi linear regression model. The results showed that: (1) there were significant reducing of plant traits, floral traits and seed production with the increasing altitude. The higher altitude, the lower plant height, the smaller leaf size, the lower number of flower per plant, the smaller floral size and the lower seed production per fruit were in population. (2) We found significant differences on phenotypic selection of plant traits and floral traits at different altitude populations. In particularly, there were significant selection differential and selection gradient on number of flower per plant and floral size in population 3 (3 681 m); however, plant height, leaf size and number of flower per plant were more likely to be chosen in population 2 (4 081 m); interestingly, selection differential and selection gradient on leaf size and number of flower per plant were marginal significant in population 1 (4 452 m). (3) Our results indicated that divergent altitudes might lead to different phenotypes, floral traits were more likely to be chosen at lower altitude. By contrast, plant height and leaf size were more likely to be chosen because of rare pollinators and resources limitation at higher altitude.
plant height; leaf size; floral traits; female fitness; phenotypic selection; altitude
1000-4025(2016)07-1443-07
10.7606/j.issn.1000-4025.2016.07.1443
2016-03-16;修改稿收到日期:2016-05-21
國家自然科學(xué)基金(31370436)
吳 云(1989-),男,在讀博士研究生,主要從事高山植物繁殖生態(tài)、進(jìn)化生態(tài)學(xué)研究。E-mail: wuyun15902858767@163.com
劉光立,副教授,碩士生導(dǎo)師,主要從事園林植物栽培與應(yīng)用研究。E-mail: liugl_1@163.com
Q948.114
A
