田菁莖瘤固氮根瘤菌對(duì)小麥葉組織的促生作用研究
2016-09-02 06:26:11陳永超齊懷廷王小晶劉華偉
西北植物學(xué)報(bào) 2016年7期
陳永超,齊懷廷,王小晶,李 強(qiáng),劉華偉
(西北農(nóng)林科技大學(xué) 生命科學(xué)學(xué)院,陜西 楊陵 712100)
?
田菁莖瘤固氮根瘤菌對(duì)小麥葉組織的促生作用研究
陳永超,齊懷廷,王小晶,李強(qiáng),劉華偉*
(西北農(nóng)林科技大學(xué) 生命科學(xué)學(xué)院,陜西 楊陵 712100)
利用GFP標(biāo)記的田菁莖瘤固氮根瘤菌(AzorhizobiumcaulinodansORS 571)侵染露白24 h的小麥種子,分別在侵染后0、6、12、24、48、72和96 h采樣,利用實(shí)時(shí)熒光定量PCR方法檢測(cè)小麥體內(nèi)6條與促生作用相關(guān)miRNAs(miR156、miR159、miR160、miR167、miR168和miR403)的表達(dá)模式,檢測(cè)其中3條miRNAs(miR159、miR167和miR168)的靶基因表達(dá)模式;以接菌8 d 的小麥樣品做切片,利用激光共聚焦顯微鏡檢測(cè)小麥葉部田菁莖瘤固氮根瘤菌的分布,并測(cè)定小麥的生理指標(biāo)。結(jié)果顯示:(1)田菁莖瘤固氮根瘤菌侵染小麥后能夠在葉片邊緣部位定殖。(2)小麥葉片中與促生作用相關(guān)的6條miRNAs出現(xiàn)了不同程度變化,在12~24 h到達(dá)其峰值,隨后逐漸下降,其中miR159在峰值時(shí)的表達(dá)量為初始表達(dá)量的2.88倍。(3)3條miRNAs的靶基因表達(dá)模式與相應(yīng)miRNA表達(dá)模式相對(duì)應(yīng),但并不嚴(yán)格。(4)生理指標(biāo)測(cè)定結(jié)果顯示,接種田菁莖瘤固氮根瘤菌對(duì)小麥葉片產(chǎn)生明顯的促生作用,其中葉鮮重在96 h的變化與對(duì)照差異極顯著。研究表明,接種的田菁莖瘤固氮根瘤菌能夠到達(dá)小麥葉組織,對(duì)小麥葉片的生長(zhǎng)產(chǎn)生明顯的促生作用,其中miRNAs在促生過(guò)程中發(fā)揮重要作用。
小麥;田菁莖瘤固氮根瘤菌;促生作用;miRNA
根瘤菌是能與豆科寄主共生,形成根瘤并固定空氣中的氮?dú)夤┲参餇I(yíng)養(yǎng)的一類(lèi)桿狀細(xì)菌。田菁莖瘤固氮根瘤菌(AzorhizobiumcaulinodansORS 571)屬于固氮根瘤菌屬,在其天然寄主豆科植物田菁(Sesbaniarostrata)的根部和莖部結(jié)瘤固氮,共生固氮效率高,且對(duì)氧的要求低于其他根瘤菌,是研究根瘤菌與非豆科植物相互作用的理想材料[1-2]。當(dāng)其作為一種植物根際促生菌時(shí),能夠?qū)⒎肿討B(tài)氮轉(zhuǎn)換為植物可利用的氨態(tài)氮形式,促進(jìn)互作植物的生長(zhǎng),增強(qiáng)互作植物抗逆境脅迫能力,提高互作植物對(duì)病原菌的免疫力[3]。田菁莖瘤固氮根瘤菌侵染小麥后能夠在其體內(nèi)定殖,也能夠在2,4-D、類(lèi)黃酮等誘導(dǎo)產(chǎn)生的類(lèi)根瘤中定殖[1,3]。但田菁莖瘤固氮根瘤菌侵染小麥后如何發(fā)揮其促生作用卻一直沒(méi)有得到合理的解釋。
miRNA屬于小RNA(small RNA)的一種,長(zhǎng)度約為22 nt,由RNA聚合酶Ⅱ轉(zhuǎn)錄,再由RNase Ⅲ的核酸內(nèi)切酶Dicer加工產(chǎn)生。miRNA能夠與RNA誘導(dǎo)的沉默復(fù)合體(RNA-induced silencing complex,RISC)結(jié)合,識(shí)別其靶mRNA,并通過(guò)兩種方式調(diào)節(jié)靶基因的表達(dá):如果miRNA與mRNA完全互補(bǔ),miRNA即引導(dǎo)mRNA特異性切割;如果兩者沒(méi)有足夠的互補(bǔ)性,則引起翻譯抑制[4]。miRNA作為主要調(diào)控因子參與植物整個(gè)生長(zhǎng)發(fā)育進(jìn)程的調(diào)控[5],處于基因表達(dá)調(diào)控的中心位置,可在不同層次上對(duì)靶基因進(jìn)行調(diào)控[6]。
李強(qiáng)[3]研究了接種田菁莖瘤固氮根瘤菌小麥根部相關(guān) miRNAs的表達(dá)變化,但接菌小麥葉組織相關(guān)miRNAs的表達(dá)變化尚未報(bào)導(dǎo)。本研究在室內(nèi)限菌條件下利用GFP標(biāo)記田菁莖瘤固氮根瘤菌侵染小麥種子,利用實(shí)時(shí)熒光定量PCR研究田菁莖瘤固氮根瘤菌侵染小麥幼苗葉中與促生作用相關(guān)miRNAs及其靶基因的表達(dá)變化,用激光共聚焦顯微鏡檢測(cè)了接菌8 d小麥葉片中田菁莖瘤固氮根瘤菌的分布情況,并測(cè)定了小麥葉片的生理指標(biāo),為深入研究田菁莖瘤固氮根瘤菌對(duì)小麥的促生機(jī)制,更好地將其應(yīng)用于農(nóng)業(yè)生產(chǎn)提供依據(jù)。
1 材料和方法
1.1材料
1.1.1小麥品種和接種菌株小麥品種‘小偃22’由西北農(nóng)林科技大學(xué)農(nóng)學(xué)院趙繼新博士提供。GFP標(biāo)記的田菁莖瘤固氮根瘤菌由本實(shí)驗(yàn)室保存。
1.1.2培養(yǎng)基與條件TY液體培養(yǎng)基:胰蛋白胨(Tryptone) 5 g/L、酵母粉(Yeast Powder)3 g/L、二水合氯化鈣(CaCl2·2H2O)0.88 g/L,pH 7.4,121 ℃滅菌20 min,用于田菁莖瘤固氮根瘤菌的活化與擴(kuò)大培養(yǎng)。PBS緩沖液:KH2PO40.24 g/L,KCl 0.2 g/L,NaCl 8 g/L,Na2HPO41.44 g/L,pH 7.4,121 ℃滅菌20 min,用于田菁莖瘤固氮根瘤菌菌液的重懸浮與稀釋。瓊脂粉-水固體培養(yǎng)基:1 L水加瓊脂粉5 g,121 ℃滅菌20 min,用于小麥種子預(yù)培養(yǎng)。1/2Hogland培養(yǎng)基配方參照遲峰[8]的方法。培養(yǎng)基質(zhì):清洗石英砂,180 ℃烘烤4 h,待降到室溫后分裝至1 L的三角瓶。每瓶加350 mL石英砂和50 mL 1/2Hogland營(yíng)養(yǎng)液,121 ℃滅菌20 min,用于小麥種子的接菌培養(yǎng)。
1.2方法
1.2.1小麥種子消毒、萌發(fā)及接菌處理選取籽粒飽滿(mǎn)、均勻一致的小麥種子,用1%次氯酸鈉浸泡10 min,用無(wú)菌水沖洗3遍;再用70%酒精浸泡30 s,無(wú)菌水沖洗3遍,完成表面消毒。參照Senthilkumar方法將表面消毒的小麥種子均勻置于大三角瓶中事先滅菌的瓊脂粉-水固體培養(yǎng)基上,腹溝朝下,胚朝向同一個(gè)方向,28 ℃萌發(fā)1 d,檢查表面消毒是否徹底[9]。消毒后的種子整齊擺放在培養(yǎng)皿內(nèi)滅菌的濾紙上,腹溝朝下,加入適量的無(wú)菌蒸餾水,置于黑暗條件下待其萌發(fā)露白。
田菁莖瘤固氮根瘤菌的復(fù)蘇與擴(kuò)大培養(yǎng):在小試管中加5 mL TY液體培養(yǎng)基,再加5 μL氨芐青霉素,挑入單菌田菁莖瘤固氮根瘤菌落,28 ℃ 200 r/min恒溫培養(yǎng)48 h使其活化。將活化的菌液按2%比例加到大三角瓶TY液體培養(yǎng)基中,擴(kuò)大培養(yǎng)至對(duì)數(shù)生長(zhǎng)期(OD600=0.6-0.8)。
參照李強(qiáng)[3]的方法,在無(wú)菌條件下將露白24 h的小麥種子移載到大三角瓶事先滅好菌的石英砂中,將20 mL田菁莖瘤固氮根瘤菌PBS稀釋液均勻加到‘小偃22’種子周?chē)灰缘润w積無(wú)菌PBS緩沖液處理同批平行露白小麥種子作為對(duì)照,用無(wú)菌透氣封口膜封住瓶口,放入光照培養(yǎng)箱培養(yǎng)并開(kāi)始計(jì)時(shí)。培養(yǎng)條件: 光照16 h,光強(qiáng)4級(jí),溫度25 ℃,相對(duì)濕度46%;黑暗8 h,光強(qiáng)0級(jí),溫度18 ℃,相對(duì)濕度37% 。
1.2.2樣本采集分別在0、6、12、24、48、72和96 h等不同時(shí)間點(diǎn)取樣,對(duì)照組與處理組同步進(jìn)行。先采集0 h的小麥樣品,在接菌處理后6、12、24、48、72和96 h等不同時(shí)間點(diǎn)分別取樣,每個(gè)樣品做3個(gè)重復(fù),平行對(duì)照組同步進(jìn)行。樣品于液氮中迅速冷凍,-80 ℃冷凍保存。接菌處理8 d后將全部樣品取出,除去黏連的石英砂,用自來(lái)水清洗表面3遍,自然風(fēng)干后挑選有代表性的樣品進(jìn)行激光共聚焦檢測(cè),其余樣品用直尺測(cè)量其葉長(zhǎng),用電子天平測(cè)量其地上部分鮮重。測(cè)量結(jié)果用Excel 2013繪制相應(yīng)曲線(xiàn),用IBM SPSS Statistics 22進(jìn)行統(tǒng)計(jì)學(xué)分析[10]。
1.2.3小麥葉組織根瘤菌的激光共聚焦檢測(cè)將選取的用以檢測(cè)田菁莖瘤固氮根瘤菌分布的小麥葉組織樣品進(jìn)行徒手切片,滴加50%甘油蓋上蓋玻片后用凡士林封片。首先用10×物鏡觀察樣品,找到需要檢測(cè)的區(qū)域,然后對(duì)檢測(cè)區(qū)域照射488和561 nm激光。GFP熒光信號(hào)由488 nm激光激發(fā)產(chǎn)生,小麥組織的自發(fā)熒光信號(hào)由561 nm 激光激發(fā)產(chǎn)生。在操作面板上以恰好能看到葉片組織為原則確定其z軸的上下界,由系統(tǒng)自動(dòng)生成最佳的層距間隔和層數(shù),從樣品表面由上到下逐層進(jìn)行掃描,采集圖像信號(hào)。在掃描得到的不同層照片間選擇最具代表性的圖像,使用軟件 FV10-ASW 3.1調(diào)整相應(yīng)的參數(shù),對(duì)樣品圖像進(jìn)行保存。
1.2.4小麥葉片總RNA的提取及miRNA的反轉(zhuǎn)錄小麥葉片總RNA采用Trizol(Invitrogen)法提取。提取總RNA使用One Step PrimeScript?miRNA cDNA Synthesis Kit(寶生物工程有限公司,大連)試劑盒進(jìn)行miRNA3’加poly A尾巴和反轉(zhuǎn)錄兩個(gè)過(guò)程。反應(yīng)體系為20 μL,包括3 μL總RNA,10 μL 2×miRNA Reaction Buffer Mix,2 μL 0.1% BSA和2 μL miRNA PrimeScript?RT Enzyme Mix。反應(yīng)條件為37 ℃ 1 h,然后85 ℃ 5 s。反轉(zhuǎn)錄產(chǎn)物保存于-20 ℃。
1.2.5miRNAs表達(dá)模式使用SYBR?Premix ExTaqTMⅡ(寶生物工程有限公司,大連)試劑盒在CFX96 Real-Time System(Bio-Rad,USA)儀器上進(jìn)行實(shí)時(shí)熒光定量PCR反應(yīng),使用β-tublin作為內(nèi)參基因,miRNA的正向引物和通用反向引物由上海生工公司合成(表1)。反應(yīng)體系為25 μL,包括1 μL cDNA、1 μL(10 μmol/L)miRNA上游引物、1 μL(10 μmol/L)下游引物、12.5 μL SYBR Premix Ex Taq和9.5 μL ddH2O。反應(yīng)條件為:95 ℃ 30 s,95 ℃ 5s,60 ℃ 30 s,40個(gè)循環(huán)。將RT-PCR產(chǎn)物的溫度從45 ℃上升到95 ℃,每升高0.5 ℃測(cè)定1 次熒光值,最后由儀器生成溶解曲線(xiàn),根據(jù)溶解曲線(xiàn)驗(yàn)證反應(yīng)產(chǎn)物的特異性。每個(gè)樣品設(shè)置3個(gè)重復(fù),每次反應(yīng)均設(shè)置1個(gè)無(wú)模板對(duì)照。實(shí)時(shí)熒光定量的miRNA表達(dá)量采用相對(duì)定量法,即2-ΔΔCt法進(jìn)行處理[11]。

表1 實(shí)時(shí)熒光定量PCR引物序列
1.2.6miRNAs的靶基因表達(dá)模式使用psRNATarget在線(xiàn)軟件預(yù)測(cè)其中3條miRNA的靶基因,在CNKI中查找對(duì)應(yīng)的基因序列并使用primer BLAST設(shè)計(jì)引物。以GAPDH作為內(nèi)參基因,所有基因的正向引物和反向引物由上海生工公司合成(表1)。反轉(zhuǎn)錄使用GoScriptTMReverse Transcriptase(Promega,美國(guó))及配套的試劑,以oligo(dT)15為引物;實(shí)時(shí)熒光定量PCR使用GoTaq?qPCR Master Mix(Promega,美國(guó))試劑盒在CFX96 Real-Time System(Bio-Rad,USA)儀器上進(jìn)行。反應(yīng)體系為20 μL,包括4 μ LcDNA,0.8 μL上下游引物,10μL GoTaq?qPCR Master Mix和5.2 μL Nuclease-Free Water。反應(yīng)條件和計(jì)算方法與1.2.5描述一致。
2 結(jié)果與分析
2.1小麥葉內(nèi)田菁莖瘤固氮根瘤菌的激光共聚焦檢測(cè)
GFP熒光蛋白在488 nm的激發(fā)光照射下能發(fā)出綠色的熒光。用激光共聚焦顯微鏡觀察處理后的小麥葉片組織,發(fā)現(xiàn)GFP標(biāo)記的田菁莖瘤固氮根瘤菌發(fā)出的綠色熒光集中在葉片邊緣部位,其他部位為紅色(圖1),說(shuō)明田菁莖瘤固氮根瘤菌能夠到達(dá)小麥葉片,并在葉片邊緣部位定殖。
2.2接菌小麥葉組織與促生作用相關(guān)miRNAs的表達(dá)模式
實(shí)時(shí)熒光定量PCR檢測(cè)結(jié)果表明,接種田菁莖瘤固氮根瘤菌小麥葉片與促生作用相關(guān)的miRNAs表現(xiàn)出了不同的表達(dá)模式(圖2)。miR156在接菌后表達(dá)量隨之波動(dòng),首先在6 h下調(diào)到初始表達(dá)量的0.36倍,隨后又在24 h達(dá)到其峰值(1.78倍),最后逐漸恢復(fù)到初始表達(dá)量的0.42倍。miR159在接菌12 h內(nèi)相對(duì)穩(wěn)定,但在24 h升到初始表達(dá)量的2.88倍,之后迅速回落,并在96 h降低到初始表達(dá)量的0.40倍。miR160在接菌6 h內(nèi)相對(duì)穩(wěn)定,在12h達(dá)到其峰值(1.54倍),之后逐漸下調(diào)到初始表達(dá)量的0.60倍。miR167在接菌6 h內(nèi)表達(dá)量下調(diào),但在12 h逐漸上升,并在24 h恢復(fù)到初始水平,隨后又下調(diào),在96 h為初始表達(dá)量的0.20倍。miR168的表達(dá)變化較為劇烈,其表達(dá)量在接菌后略微下調(diào),在12 h升到最大值(1.98倍)后逐漸降落,在96 h為初始表達(dá)量的0.16倍。miR403的表達(dá)量則在接菌后穩(wěn)步上升,在24 h達(dá)到初始表達(dá)量的1.77倍,隨后急劇下調(diào),在96 h為初始表達(dá)量的0.31倍。
綜上所述,接種田菁莖瘤固氮根瘤菌小麥葉片中與促生作用相關(guān)的6條miRNAs在接菌12 h或48 h達(dá)到峰值,隨后逐步跌落并在96 h達(dá)到最低水平。其不同之處在于各miRNA表達(dá)量到達(dá)峰值后的持續(xù)時(shí)間和峰值的相對(duì)表達(dá)量。

圖1 GFP標(biāo)記的田菁莖瘤固氮根瘤菌定殖于小麥的葉片邊緣部位Fig. 1 GFP-labeled A. caulinodans ORS 571 colonized in margin area of wheat leaves
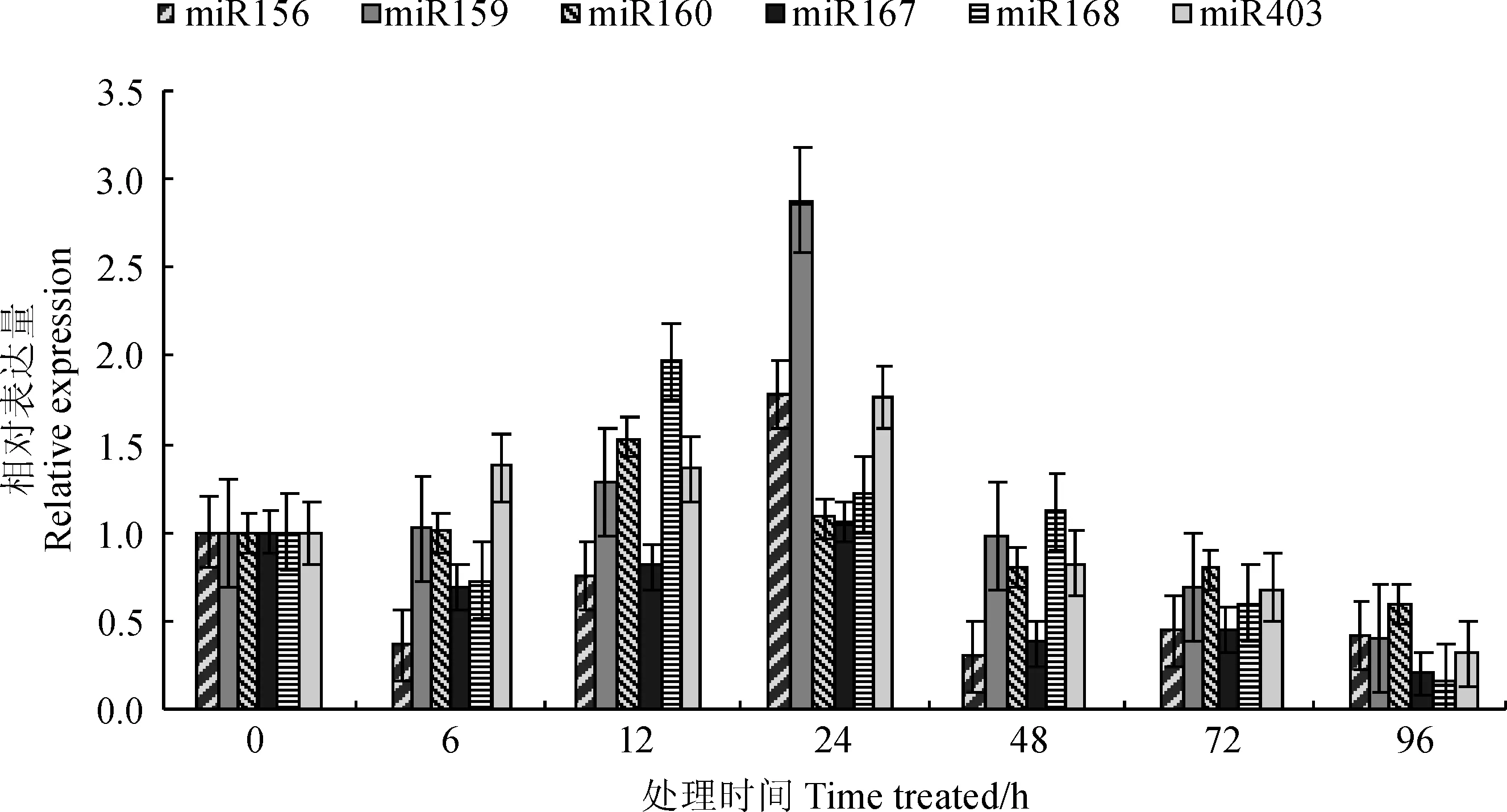
miRNAs在各時(shí)間點(diǎn)的表達(dá)量均相對(duì)于其0 h的初始表達(dá)量(設(shè)定為1)進(jìn)行計(jì)算圖2 接種田菁莖瘤固氮根瘤菌的小麥葉組織與促生作用相關(guān)miRNAs在不同時(shí)間點(diǎn)的表達(dá)模式All expression data were calculated based on the score of 0-pionts which were set as 1Fig. 2 The expression pattern of miRNAs in different time points relating to the growth-promoting effect of wheat leaves inoculated by A. caulinodans ORS 571
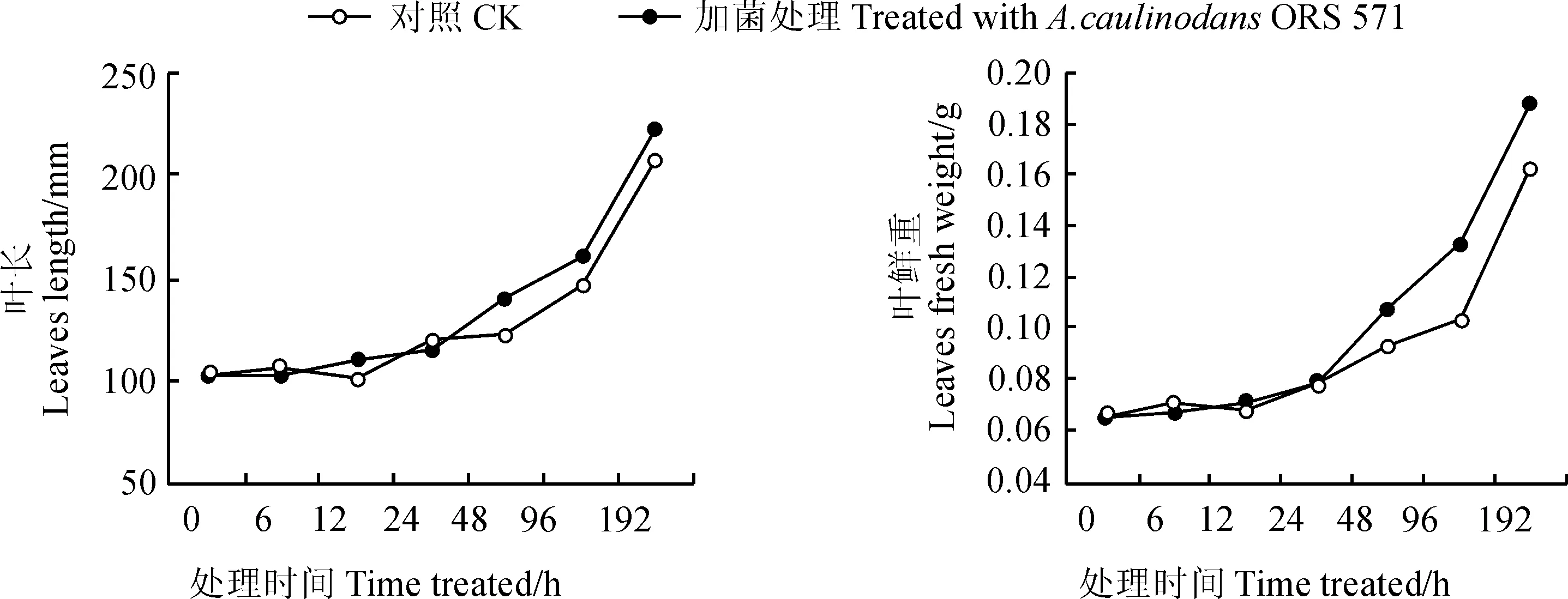
圖3 接種田菁莖瘤固氮根瘤菌后葉長(zhǎng)和葉鮮重的變化Fig. 3 Change of leaf length and leaf fresh weight of leaves after adding GFP-labeled A. caulinodans ORS 571
2.3接菌小麥葉部生理指標(biāo)分析
在用田菁莖瘤固氮根瘤菌接種小麥后,分別在取樣的各個(gè)時(shí)間段測(cè)量小麥葉長(zhǎng)度、葉鮮重等生理指標(biāo)(圖3),測(cè)定結(jié)果表明,接菌小麥的葉長(zhǎng)和葉鮮重均發(fā)生明顯變化,其中葉鮮重在96 h的變化差異極顯著(P<0.01)。
2.4小麥葉組織促生相關(guān)miRNAs的靶基因表達(dá)分析
實(shí)時(shí)熒光定量PCR檢測(cè)結(jié)果表明,促生作用相關(guān)的miRNAs靶基因表達(dá)在處理后發(fā)生了較大的變化(圖4)。其中miR159的靶基因TaMyb3在6和12 h的表達(dá)都受到了抑制,在48 h后達(dá)到表達(dá)峰值,為初始表達(dá)量的3.10倍,而在96 h則逐漸恢復(fù)到相對(duì)較低的表達(dá)水平(2.19倍);miR167的靶基因OsARF8的表達(dá)量則在12 h內(nèi)相對(duì)平穩(wěn),在24 h達(dá)到峰值(6.78倍),隨后一直保持較高的表達(dá)水平。miR168的靶基因OsAGO1在6與12 h的表達(dá)量都稍高于初始表達(dá)量,而OsAGO1在24 h達(dá)到表達(dá)峰值(9.50倍)后逐步下降到初始表達(dá)量的2.04倍。
3 討 論
3.1根瘤菌在小麥葉片部位的共聚焦檢測(cè)
李強(qiáng)[3]研究表明,田菁莖瘤固氮根瘤菌能夠定殖于小麥的根表皮和細(xì)胞間,同時(shí)也能向上遷移至小麥葉片部位。本研究使用激光共聚焦顯微鏡檢測(cè)小麥葉片GFP標(biāo)記田菁莖瘤固氮根瘤菌的熒光,在葉片邊緣處觀測(cè)到大量GFP標(biāo)記田菁莖瘤固氮根瘤菌,表明田菁莖瘤固氮根瘤菌能夠定殖于小麥的葉片組織。
3.2接菌小麥葉部促生作用相關(guān) miRNAs 表達(dá)分析
miR156負(fù)調(diào)控其靶基因SBP-box(Squamosa promoter binding protein),或稱(chēng)SPL(SQUAMOSA Promoter-binding protein-like)。該基因是植物特有的一類(lèi)轉(zhuǎn)錄因子,廣泛參與植物生長(zhǎng)發(fā)育[12], 如葉片發(fā)育[13]、生長(zhǎng)時(shí)期轉(zhuǎn)換[14]、抗逆[15]和植物株型[16]等。在玉米中miR156調(diào)控幼年期到成年期的轉(zhuǎn)變,并調(diào)控成年期表皮細(xì)胞的形成[17];在水稻中miR156調(diào)控幼年期到成年期的轉(zhuǎn)變[18]。本研究中,miR156在接菌處理后下調(diào),隨后又逐漸上升,最后又下調(diào),表明接種田菁莖瘤固氮根瘤菌引起了miR156對(duì)小麥SPLs等靶基因家族的調(diào)控,加速了小麥幼年期到成年期的轉(zhuǎn)變進(jìn)程,從而發(fā)揮了促生的作用,從生理指標(biāo)來(lái)看,在接菌24 h后小麥葉片的鮮重和長(zhǎng)度變化都有了較為明顯的增加。
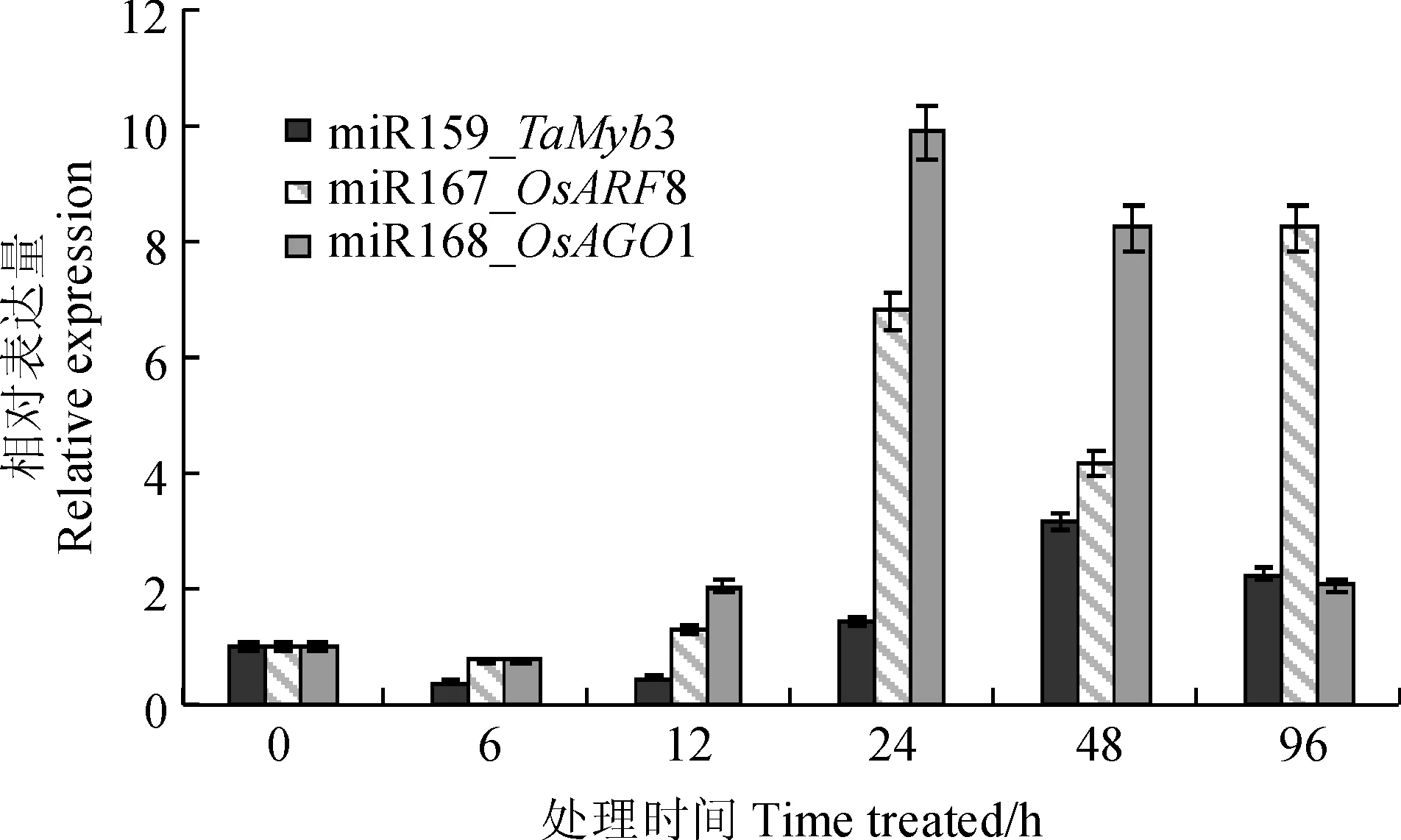
miRNAs靶基因在各時(shí)間點(diǎn)的表達(dá)量均相對(duì)其0 h的初始表達(dá)量(設(shè)定為1)進(jìn)行計(jì)算圖4 接種田菁莖瘤固氮根瘤菌小麥葉部與促生作用相關(guān)miRNAs靶基因在不同時(shí)間點(diǎn)的相對(duì)表達(dá)量All expression data were calculated based on the score of 0-pionts which was set as 1Fig.4 The relative expression of miRNAs’ target genes in different time points relating to the growth-promoting effect of wheat leaves inoculated by A. caulinodans ORS 571
miR159編碼Myb3轉(zhuǎn)錄因子,靶基因?yàn)镚AMYB基因家族(編碼 GAMYB 轉(zhuǎn)錄因子,參與赤霉素信號(hào)調(diào)節(jié)[19])。miR159不僅參與植物營(yíng)養(yǎng)生長(zhǎng)也參與生殖生長(zhǎng)[20]。Pluskota 等[21]研究發(fā)現(xiàn) miR156、miR159、miR160 參與種子的萌發(fā)過(guò)程。miR156通過(guò)調(diào)控激素在胚根萌發(fā)之后發(fā)揮作用;而miR159則在胚根萌發(fā)之前發(fā)揮作用。在小麥中miR159在根瘤菌侵染后首先維持其正常水平,在24 h達(dá)到其峰值,最后維持在較低的表達(dá)水平,表明miR159可能在小麥與田菁莖瘤固氮根瘤菌相互作用過(guò)程中參與調(diào)控,促進(jìn)了小麥的營(yíng)養(yǎng)生長(zhǎng)進(jìn)程。
miR160的靶基因?yàn)锳RF10、ARF16、ARF17(auxin response factor),參與生長(zhǎng)素信號(hào)調(diào)節(jié)。過(guò)表達(dá)突變的ARF17(5mARF17,對(duì)miR160具有抗性)導(dǎo)致許多生長(zhǎng)激素應(yīng)答基因表達(dá)改變,擬南芥呈現(xiàn)畸形發(fā)育,如葉片鋸齒狀突起、卷曲、育性降低等[22-23]。在擬南芥、水稻、玉米等植物中,miR160 通過(guò)負(fù)調(diào)控ARF而有效地影響種子萌發(fā)、苗期發(fā)育及根冠形成等生理發(fā)育過(guò)程[24-25]。在小麥接菌處理后,miR160逐漸加強(qiáng)了對(duì)ARF基因家族的負(fù)調(diào)控,24 h后表達(dá)又逐漸降低,從而影響小麥苗期發(fā)育和根冠形成。但miR160的表達(dá)強(qiáng)度低于miR156和miR159,其表達(dá)峰值僅為初始表達(dá)量的1.54倍。
miR167靶向ARF6、ARF8[26],在玉米中,miR167受干旱脅迫表達(dá)下調(diào),從而導(dǎo)致其各自的靶基因磷脂酶D表達(dá)上調(diào),進(jìn)而導(dǎo)致植物葉片氣孔關(guān)閉以及多種抗氧化系統(tǒng)活性的增加,最終提高了對(duì)干旱的抵抗能力[27]。此外,在擬南芥中,ARF8調(diào)控著生長(zhǎng)素介導(dǎo)的側(cè)根結(jié)構(gòu)的性狀,從而影響擬南芥的生長(zhǎng)發(fā)育[28]。推測(cè)接種田菁莖瘤固氮根瘤菌后,miR167表達(dá)逐漸降低,其靶基因ARF家族表達(dá)上調(diào),通過(guò)調(diào)控小麥側(cè)根發(fā)育發(fā)揮其促生作用。
miR168 靶基因編碼蛋白AGO1是RISC(RNA誘導(dǎo)沉默復(fù)合體)的重要組成部分[29]。miR168靶向AGO1,兩者之間存在負(fù)反饋調(diào)節(jié),AGO1蛋白參與miRNA自身積累調(diào)控,它與miR168的平衡可以保證其它miRNAs處于正常狀態(tài)[30]。此外,miR168也與植物的抗逆、抗旱等有關(guān)[31]。田菁莖瘤固氮根瘤菌侵染小麥12 h后miR168表達(dá)量上升,由于miR168與靶基因AGO1之間存在負(fù)反饋調(diào)節(jié),因此miR168在表達(dá)量上升后又逐漸回落,在96 h降到初始表達(dá)量的0.16倍,發(fā)揮其促生抗逆等作用。
miR403靶向AGO2,并通過(guò)負(fù)反饋?zhàn)饔脜⑴c自身代謝途徑的調(diào)控[6]。在擬南芥中,miR403可以通過(guò)miR403-AGO2-miR168-AGO1循環(huán)途徑參與其生長(zhǎng)發(fā)育的調(diào)控過(guò)程[32]。本研究中miR403的相對(duì)表達(dá)量首先上調(diào)然后下調(diào),這一變化表明接種田菁莖瘤固氮根瘤菌可能通過(guò)調(diào)節(jié)AGO家族蛋白的表達(dá)來(lái)控制相關(guān)miRNAs的合成,加快miR403的積累,調(diào)節(jié)小麥幼苗的自身代謝,適應(yīng)接種田菁莖瘤固氮根瘤菌所產(chǎn)生的變化。
3.3接種根瘤菌對(duì)小麥葉片生理指標(biāo)的影響
在接種田菁莖瘤固氮根瘤菌侵染后,小麥的葉長(zhǎng)度及地上部分鮮重均發(fā)生明顯的變化。在96 h接菌處理組和對(duì)照組的葉鮮重分別為0.133和0.104,在統(tǒng)計(jì)學(xué)分析中差異極顯著,葉長(zhǎng)平均值分別為151.90和146.20。表明接種田菁莖瘤固氮根瘤菌對(duì)小麥葉組織具有促生作用,使葉片生物量累積并加速伸長(zhǎng),強(qiáng)化了對(duì)營(yíng)養(yǎng)元素的吸收和利用,這與本研究前期室內(nèi)和田間試驗(yàn)結(jié)果一致[33]。這些結(jié)果也與3.2中所討論的miRNAs變化相輔相成,即由內(nèi)源的miRNAs變化調(diào)節(jié)了小麥的外部生長(zhǎng)狀態(tài),而接菌小麥所表現(xiàn)出的促生作用也進(jìn)一步證實(shí)了小麥體內(nèi)由于miRNAs的變化而發(fā)生的一系列基因表達(dá)變化。
3.4小麥葉組織促生相關(guān)miRNAs的靶基因表達(dá)分析
本研究以小麥的TaMyb3作為miR159的靶基因進(jìn)行驗(yàn)證。在接菌處理后miR159的表達(dá)量不斷上升,在24 h達(dá)到峰值后逐漸下降,而TaMyb3的表達(dá)量則在接菌處理后隨即下調(diào),從24 h開(kāi)始逐漸上升,并在48 h達(dá)到峰值,96 h維持在2.19倍。表明miR159調(diào)控TaMyb3,并通過(guò)TaMyb3控制其下游赤霉素信號(hào)相關(guān)基因的表達(dá),促進(jìn)小麥生長(zhǎng)。但TaMyb3的表達(dá)可能還受到其他因素的影響,因此在6和24 h時(shí)間點(diǎn)的表達(dá)量并沒(méi)有與miR159的表達(dá)高度一致。
本研究以水稻的OsARF8作為miR167的靶基因進(jìn)行驗(yàn)證。與miR167的表達(dá)相對(duì)應(yīng),OsARF8在接菌處理后即開(kāi)始變化,但直到12 h僅有對(duì)照的1.26倍,在24 h大量表達(dá)并達(dá)到其峰值,之后一直維持較高的表達(dá)水平。表明miR167負(fù)調(diào)控ARF8,從而通過(guò)影響側(cè)根發(fā)育發(fā)揮其促生作用。但ARF8在各個(gè)時(shí)間點(diǎn)的表達(dá)量不僅受到miR167的調(diào)控,可能也受到外界環(huán)境和自身生長(zhǎng)狀態(tài)的影響。
本研究以水稻OsAGO1作為miR168的靶基因進(jìn)行驗(yàn)證。miR168的表達(dá)量在接菌處理12 h升高,24 h則下降并一直維持在較低的水平,與之對(duì)應(yīng)的OsAGO1基因則在24 h達(dá)到峰值并在后期維持較高的表達(dá)水平。表明AGO1 受miR168的調(diào)控而繼續(xù)參與其他miRNA的積累,并改變其下游基因的表達(dá),發(fā)揮促生作用。
綜上所述,小麥葉片中與促生作用相關(guān)的miRNAs在接菌處理后能夠調(diào)節(jié)其各自的靶基因,從而發(fā)揮其各自的功能。但值得注意的是本研究中所選取的靶基因并非只受到上游miRNAs的調(diào)節(jié),其他的內(nèi)外因素亦對(duì)靶基因產(chǎn)生影響,從而使其表達(dá)并不能與miRNAs的表達(dá)嚴(yán)格匹配。
[1]陳廷偉, SCHERER S, B?GER P. 固氮根瘤菌(Azorhizoboium)在人工誘發(fā)小麥類(lèi)根瘤種的固氮作用[J]. 中國(guó)科學(xué)B輯, 1992,22(7): 712-716.
CHEN T W, SCHERER S, B?GER P. The azotification effect ofAzorhizobiumin artificially induced para-nodules in wheat[J].ScienceinChinaSeriesB:Chemistry, 1992,22(7):712-716.
[2]LEE K B, BACKER P D, AONO T,etal. The genome of the versatile nitrogen fixerAzorhizobiumcaulinodansORS571[J].BMCGenomics, 2008, 9(25):271.
[3]李強(qiáng).田菁莖瘤固氮根瘤菌與小麥互作中相關(guān)microRNA鑒定及其功能分析[D]. 楊陵: 西北農(nóng)林科技大學(xué), 2014.
[4]JACKSON R J, STANDART N. How do microRNAs regulate gene expression[J].ScienceSTKE, 2007,367:rel.
[5]CHEN X. MicroRNA biogenesis and function in plants[J].FEBSLetters, 2005, 579(26): 5 923-5 931.
[6]VOINNET O. Origin, biogenesis, and activity of plant microRNAs[J].Cell, 2009,136(4): 669-687.
[7]陳新宏, 李璋, 趙繼新,等. 國(guó)審小麥新品種小偃22的研究與利用[J]. 農(nóng)藝科學(xué), 2007,23(9): 218-220.
CHEN X H, LI Z, ZHAO J X,etal. Study and utilize on a new wheat variety of nation approve Xiaoyan22[J].ChineseAgriculturalScienceBulletin, 2007,23(9): 218-220.
[8]遲峰. 根瘤菌在植物內(nèi)的遷移運(yùn)動(dòng)及其與植物相互作用的蛋白質(zhì)組學(xué)研究[D]. 北京: 中國(guó)科學(xué)院研究生院, 2006.
[9]SENTHIKUMAR M, MADHAIYAN M, SUNDARAM S P,etal. Induction of endophytic colonization in rice (OryzasativaL.) tissue culture plants byAzorhizobiumcaulinodans[J].BiotechnologyLetters, 2008,30(8): 1 477-1 487.
[10]馬蕊, 張愛(ài)霞, 生慶海. SPSS軟件在食品感官評(píng)分結(jié)果分析中的應(yīng)用[J]. 乳業(yè)科學(xué)與技術(shù), 2007,30(1): 12-14.
MA R, ZHANG A X,SHENG Q H. The application of SPSS in analyzing the results of food sensory scoring[J].DairyIndustryScienceandTechnology, 2007,30(1): 12-14.
[11]LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod[J].Methods, 2001,25(4): 402-408.
[12]CHEN X B, ZHANG Z L, LIU D M,etal.SQUAMOSApromoter-binding protein -like transcription factors: star players for plant growth and development[J].JournalofIntegrativePlantBiology, 2010,52(11): 946-951.
[13]WU G, POETHIG R S. Temporal regulation of shoot development inArabidopsisthalianaby miR156 and its targetSPL3[J].Development, 2006, 133(18): 3 539-3 547.
[14]USAMI T, HORIGUCHI G, YANO S,etal. The more and smaller cells mutants ofArabidopsisthalianaidentify novel roles forSQUAMOSAPROMOTERBINDINGPROTEIN-LIKEgenes in the control of heteroblasty[J].Development, 2009, 136(6): 955-964.
[15]王冰, 宋娜, 孫燕飛,等, MicroRNA156家族在小麥非生物脅迫中的表達(dá)分析[J]. 植物病理學(xué)報(bào), 2013,43(2): 201-204.
WANG B, SONG N, SUN Y F,etal. Expression analysis of wheat mircoRNA156 family in response to abiotic stress[J].ActaPhytopathologicaSinica, 2013,43(2): 201-204.
[16]JIAO Y, WANG Y, XUE D,etal. Regulation ofOsSPL14 by OsmiR156 defines ideal plant architecture in rice[J].NatureGenetics, 2010,42(6): 541-544.
[17]CHUCK G, CIGAN A M, SAETEURN K,etal. The heterochronic maize mutantCorngrass1 results from overexpression of a tandem microRNA[J].NatureGenetics, 2007,39(4): 544-549.
[18]WANG S, ZHU Q H, GUO X,etal. Molecular evolution and selection of a gene encoding two tandem microRNAs in rice[J].FEBSLetters, 2007,581(24): 4 789-4 793.
[19]REYES J L, CHUA N H. ABA induction of miR159 controls transcript levels of two MYB factors duringArabidopsisseed germination[J].ThePlantJ., 2007,49(4): 592-606.
[20]ALONSO-PERAL M M, LI J, LI Y,etal. The microRNA159-regulatedGAMYB-likegenes inhibit growth and promote programmed cell death in Arabidopsis[J].PlantPhysiology, 2010,154(2): 757-771.
[21]PLUSKOTA W E, MARTINEZ-ANDUJAR C, MARTIN R C,etal. MicroRNA function in seed biology[M].ERDMANN V A, BARCISZEWSKI J. Non coding RNAs in plants. New York: Springer-Verlag Berlin Heidelberg, 2011: 339-357.
[22]TURNER M, NIZAMPATNAM N R, BARON M,etal. Ectopic expression of miR160 results in auxin hypersensitivity, cytokinin hyposensitivity, and inhibition of symbiotic nodule development in soybean[J].PlantPhysiology, 2013,162(4): 2 042-2 055.
[23]MALLORY A C, BARTEL D P, BARTEL B. MicroRNA-directed regulation of Arabidopsisauxinresponsefactor17 is essential for proper development and modulates expression of early auxin response gene[J].ThePlantCell, 2005,17(5): 1 360-1 375.
[24]WANG J W, WANG L J, MAO Y B,etal. Control of root cap formation by microRNA-targeted auxin response factors in arabidopsis[J].ThePlantCell, 2005,17(8): 2 204-2 216.
[25]LIU P P, MONTGOMERY T A, FAHLGREN N,etal. Repression ofAUXINRESPONSEFACTOR10 by microRNA160 is critical for seed germination and post-germination stages[J].ThePlantJournal, 2007,52(1): 133-146.
[26]王磊, 范云六. 植物微小RNA(microRNA)研究進(jìn)展[J]. 中國(guó)農(nóng)業(yè)科技導(dǎo)報(bào), 2007,9(3): 18-23.
WANG L, FAN Y L. Progress of microRNA in plants[J].JournalofAgriculturalScienceandTechnology, 2007,9(3): 18-23.
[27]WEI L Y, ZHANG D F, XIANG F,etal. Differentially expressed miRNAs potentially involved in the regulation of defense mechanism to drought stress in maize seedlings[J].InternationalJournalofPlantSciences, 2009,170(8): 979-989.
[28]GIFFORD M L, DEAN A, GUTIERREZ R A, Gutierrez,etal. Cell-specific nitrogen response mediate developmental plasticity[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2008,105(2):803-808.
[29]BAUMBERGER N, BAULCOMBE D C. Arabidopsis ARGO-NAUTEI is an RNA slicer that selectively recruits microRNAs and short interfering RNAs[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2005,102(33): 11 928-11 933.
[30]VAUCHERET H, MALLORY A C, BARTEL D P. AGO 1 home-stasis entails co-expression ofmiR168 andAGO1 and preferential stabilization of miR168 by AGO1[J].MolecularCell, 2006,22(1): 129-136.
[31]韓璐, 欒雨時(shí). 植物miR168的研究進(jìn)展[J]. 基因組學(xué)與應(yīng)用生物學(xué), 2014,33(2): 438-444.
HAN L, LUAN Y S. Progress on the research of plant miR168[J].GenomicsandAppliedBiology, 2014,33(2): 438-444.
[32]ZHANG C, XIAN Z Q, HUANG W,etal, Evidence for the biological function of miR403 in tomato development[J].ScientiaHorticulturae, 2015,197: 619-626.
[33]劉華偉, 孫超, 楊呼,等. 田菁莖瘤固氮根瘤菌對(duì)小麥種子侵染的促生作用及其在根系內(nèi)的定殖[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào), 2012,18(1): 210-217.
LIU H W, SUN C, YANG H,etal. Promotion for wheat growth and root colonization after infecting wheat seeds withAzorhizobiumcaulinodans[J].PlantNutritionandFertilizerScience. 2012,18(1): 210-217.
(編輯:潘新社)
Growth-Promoting Effect ofAzorhizobiumcaulinodansto Wheat Leaves
CHEN Yongchao, QI Huaiting, WANG Xiaojing, LI Qiang, LIU Huawei*
(College of Life Science, Northwest A&F University, Yangling, Shaanxi 712100,China)
In this study, germinated 24h wheat seeds were inoculated withAzorhizobiumcaulinodans, samples were collected at 0, 6, 12, 24, 48, 72 and 96 h, respectively and 6 miRNAs (miR156, miR159, miR160, miR167, miR168 and miR403)in wheat leaves, 3 of these miRNAs’ target genes were detected using Real-time quantifying PCR. The distribution of GFP-labeledA.caulinodansin 8 d wheat leaves were detectedviaconfocal microscopy, and relevant physiological indices were measured and compared. The results showed that: (1) GFP-labeledA.caulinodanscould colonize in margin area of wheat leaves. (2) All 6 miRNAs relating to growth-promoting varied in different degrees after treatment withA.caulinodans, and soared to their peaks at 12-24 h. The peak of miR159 was 2.88 times as strong as regular expression. (3) The expression of 3 miRNAs’ target genes were corespondent to their upstream miRNA, but not restrict. (4) Physiological indices of length and fresh weight of leaves showed sensible growth promoting, the weight of leaves in 96 h showed significant difference statistically (P<0.01). The study suggested that:A.caulinodansinoculated in wheat could colonize leaves, exerting evident growth-promoting effect to wheat leaves, and miRNAs play an intrinsic role in the growthpromotion.
wheat;AzorhizobiumcaulinodansORS 571; growth-promoting; miRNA
1000-4025(2016)07-1383-08
10.7606/j.issn.1000-4025.2016.07.1383
2016-04-12;修改稿收到日期:2016-06-14
國(guó)家自然科學(xué)基金(31071870);陜西省農(nóng)業(yè)科技創(chuàng)新與攻關(guān)項(xiàng)目(2015NY006)
陳永超(1990-),男,碩士,主要從事非豆科作物固氮研究。E-mail: break0910@126.com
劉華偉,博士,副教授,主要從事非豆科作物內(nèi)生固氮的分子機(jī)理研究。E-mail:bioche@nwsuaf.edu.cn
Q939.11+4;Q789
A
