荒漠綠洲區不同種植年限人工梭梭林土壤化學計量特征
2016-08-09 01:16:22蘇永中劉婷娜
生態學報 2016年11期
張 珂,蘇永中,王 婷,劉婷娜
1 中國科學院寒區旱區環境與工程研究所臨澤內陸河流域研究站,蘭州 730000 2 中國科學院大學,北京 100049
曾清蘋1,何丙輝1,*,毛巧芝1,秦華軍2,李 源3,黃 祺4
1 西南大學資源環境學院,三峽庫區生態環境教育部重點實驗室,重慶 400715 2 貴州省生物研究所,貴陽 550009 3 重慶市涪陵區林業局,重慶 408000 4 中國電建集團貴陽勘測設計研究院有限公司,貴陽 550009
?
荒漠綠洲區不同種植年限人工梭梭林土壤化學計量特征
張珂1,2,蘇永中1,*,王婷1,2,劉婷娜1
1 中國科學院寒區旱區環境與工程研究所臨澤內陸河流域研究站,蘭州730000 2 中國科學院大學,北京100049
摘要:為闡明梭梭建立對林下土壤養分化學計量特征的影響,分析了2、5、9、13、16、31、39a荒漠綠洲區梭梭林灌叢下和流沙區(0a)土壤有機碳(SOC)、全氮(TN)、全磷(TP)、碳酸鈣(CaCO3)、速效磷(Available P)含量及其化學計量特征變化規律。結果顯示:1)SOC和TN在9a后出現顯著的成層化分布,而TP的這一特征相對滯后;不同土壤深度SOC、TN均隨林齡增加而顯著增加,而TP未表現出明顯變化。2)C∶P和N∶P在9a后表現出明顯的成層化分布且不同土層C∶P和N∶P隨林齡增加顯著增加,而C∶N保持相對穩定。3)較低含量的Available P在2a后即表現出0—5 cm含量大于5—20 cm且表層Available P和CaCO3隨林齡增加而顯著增加。而CaCO3∶Available P在不同林齡間并未發生顯著變化。 4)隨林齡增加,土壤表層風蝕可蝕性極顯著降低且與土壤表層養分含量呈顯著的負相關關系。以上結果表明,梭梭的建立提高了土壤SOC和TN含量且隨林齡增加變化更顯著,而變化較小的C∶N說明土壤氮的形成需要固定比例的碳。變化幅度較小的TP說明主要來源于巖石風化的磷素受時間作用的限制,而較低含量的Available P和變化幅度較小的CaCO3∶Available P則表明梭梭的生長受P的限制且有限的Available P易被CaCO3固定。另外,土壤養分含量與土壤風蝕可蝕性顯著的負相關關系,進一步說明梭梭的建立改善了土壤質地,增加了土壤養分含量,這對荒漠綠洲區土壤恢復和植被建設有極大的促進和指導作用。
關鍵詞:人工梭梭林;林齡;土壤化學計量特征;荒漠-綠洲區
生態化學計量學作為近年來對生態系統養分循環研究的一種重要工具,是分析多重化學元素(尤其是碳(C)、氮(N)、磷(P))平衡關系及其生態系統交互影響的理論依據[1- 3]。目前,生態化學計量學研究的重點主要集中在對植物葉片中C∶N∶P化學計量特征的研究,其中,C∶N和C∶P反映了植物在同化CO2同時對N、P的吸收利用情況,N∶P臨界值則是判斷土壤對植物養分供應狀況的指標[4- 5]。作為影響植物養分特征的重要因子,土壤是一個比較復雜的生態系統,由于其不可移動性,且受植被類型、蓋度、氣候、地形、土壤母質、生物多樣性及發育時間等因素的影響,使得土壤生態系統具有較高的空間異質性,且不同區域土壤養分循環特征也有顯著差異[6-8]。如,在較高緯度易受風蝕影響的荒漠區,植物體生命活動不僅受水分短缺的影響,還受土壤貧瘠養分以及脅迫環境的限制,從而影響著植物葉片中的養分元素含量[9- 11]。因此,對不同區域不同植被類型土壤化學計量特征的研究顯得尤為重要。
在干旱荒漠區建立人工固沙植被是生態恢復的重要措施之一[12]。研究表明,人工固沙林的建立能顯著提高了土壤表層有機質和N、P等養分含量,且隨著人工植被建立時間的延長土壤肥力顯著改變[13- 15]。另外,人工植被對流沙的固定不僅為物種繁衍和生物多樣性的恢復創造必要的環境條件,而且促進了土壤的形及空間異質性的改變,進一步影響著植被格局的變化[13]。自20世紀70年代以來,為保護綠洲環境及恢復由沙化導致的退化的植被-土壤系統,梭梭作為重要的人工固沙植被在河西走廊中段荒漠綠洲帶被廣泛的種植[16]。隨著不同時間梭梭人工防護林的建立,形成了不同林齡的防護林系統并起到了極大的防風固沙作用[16]。現有的對梭梭的研究顯示,當梭梭林齡大于10a時,由于很難利用到深達20 m的地下水及干旱區有限的降水,梭梭林出現自疏現象,這對梭梭在干旱區固沙可持續發展產生重要的影響[17-18]。然而,隨梭梭林齡的增加,土壤養分特征如何變化?土壤狀況是否發生顯著改變?土壤狀況的改變與土壤養分特征的關系如何?對上述問題的研究將豐富對梭梭的研究并對荒漠綠洲區人工梭梭林的建立和生態恢復提供一定的理論指導。
本研究以荒漠綠洲區不同種植年限梭梭林為研究對象,考慮到梭梭自身獨特的泌鹽特性及其他生物因子的共同作用導致土壤環境具有相對較高的鹽分含量,而鹽分離子中高含量的Ca2+又對土壤P的有效性有顯著的影響[19-21],因此,本研究將比較不同種植年限梭梭灌叢下土壤有機碳(SOC)、全氮(TN)、全磷(TP)、速效磷(Available P)、碳酸鈣(CaCO3)含量,分析不同林齡土壤化學計量特征及其影響因子,從而揭示荒漠-綠洲區土壤養分平衡和循環,為梭梭林在荒漠綠洲區的可持續發展、環境保護和生態恢復提供理論依據及參考資料。
1研究區概況
研究區位于黑河中游甘肅省臨澤縣中科院臨澤內陸河流域研究站區(39°21′N,100°07′E,海拔1350 m)東北部,為近幾十年來由綠洲向荒漠擴展的過渡區。綠洲外圍與巴丹吉林沙漠南緣和戈壁荒漠相接。該區域為典型的荒漠氣候,年均降雨量為117 mm且多集中在7—9月份,年蒸發量2390 mm,年均氣溫7.6℃,無霜期165 d。西北風向為主,年均風速為3.2 m/s,大風日數(> 17 m/s)為15 d,風沙活動主要集中在3—5月[16]。地下水深度為4—10 m。該區地帶性土壤為灰棕漠土,由于受風沙長期侵襲,在綠洲邊緣有大片的表層結構疏松且有機質含量很低的風沙土發育,極易發生風蝕。綠洲邊緣主要荒漠植物種有紅砂(Reaumuriasoongarica)、珍珠(Salsolapasserina)、泡泡刺(Nitrariatangutorum)、沙拐棗(Calligonummongolicum)等灌木半灌木和堿蓬(Suaedaprzewalskii)、沙米(Agriophyllumsquarrosum)、霧冰藜(Bassiadasyphylla)、白莖鹽生草(Halogetonarachnoideus)等1年生草本植物[16,22]。

圖1 不同林齡梭梭林采樣點分布圖Fig.1 Distribution of sampling sites of Haloxylon ammodendron in different plantation ages
2材料與方法
2.1植物-土壤樣品采集
于2014年8月初在研究區選擇通過植苗建立的2、 5、 9、 13、 16、 31、39年的梭梭林為研究對象(圖1)。在每個年代梭梭林中分別建立3個10 m × 10 m的研究樣地,每個樣地間隔約10 m。在每個樣地分別選擇5株大小長勢相似的梭梭 (表1),以每株梭梭根部為中心,在距每株梭梭根部5 cm處的4個方向(東、西、南、北)上采集0—5 cm和5—20 cm土壤樣品并混合。同時,在流動沙丘處采集土壤樣品作為參考(0a)。
2.2土壤養分測定
土壤樣品于室內自然風干,研磨過100目篩制成供試樣品。土壤有機碳(SOC)采用重鉻酸解外加熱法測定,土壤全碳(TC)和全氮(TN)用德國元素分析儀進行測定(a vario MACRO CUBE elementar,Germany),土壤全磷(P)和速效磷(Available P)分別采用H2SO4-H2O2消煮法和NaHCO3提取法。土壤CaCO3含量利用土壤無機碳含量轉化而來,公式為CaCO3(mg/g) = (TC-SOC) × 8.33[23]。
2.3數據處理
土壤風蝕可蝕性結合了土壤物理和化學性質,利用公式EF(Erodible Fraction) (%) = 0.29 + 0.31Sa + 0.17Si + 0.33Sc-2.59OM-0.95CaCO3(R2= 0.67)來評價土壤對于風力引起的剝離和搬運的敏感程度,式中,Sa、Si、Sc和OM分別代表土壤砂粒含量、土壤粉粒含量、土壤黏粒含量和土壤有機質含量,所有指標均用%表示[24-25]。
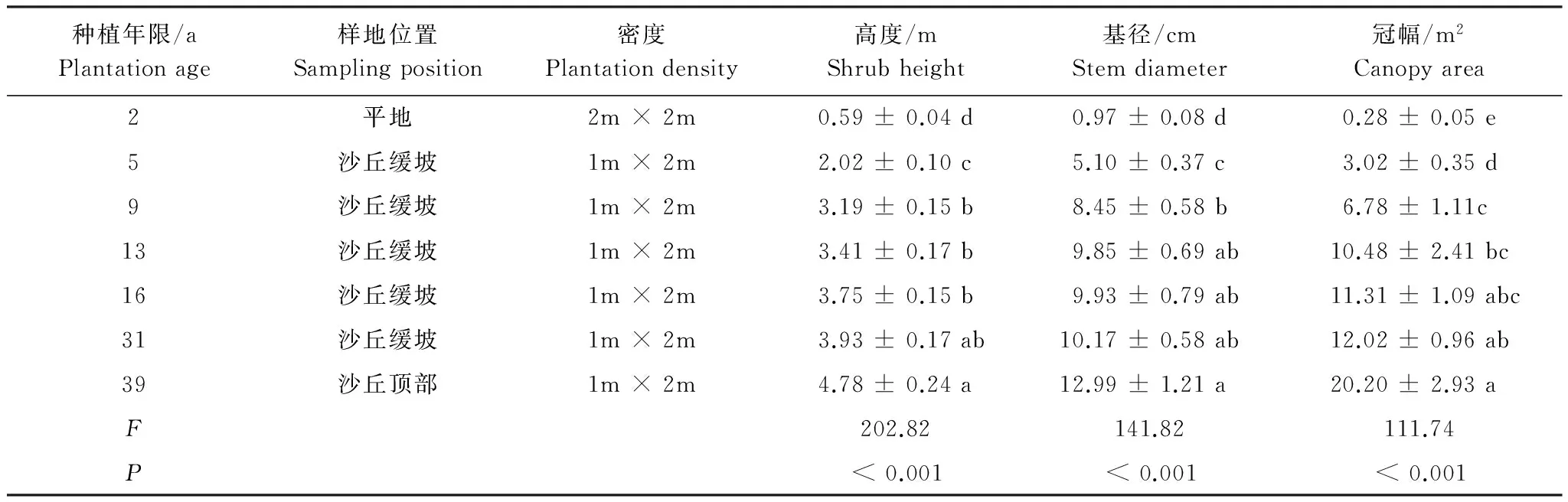
表1 不同林齡梭梭林形態特征
數據分析采用SPSS16.0軟件進行分析,用ORINGIN 9.2做圖。土壤SOC、TN、TP含量均采用質量含量,C∶N、C∶P、N∶P、CaCO3∶Available P采用質量比。用One-Sample T test 對0—5 cm和5—20 cm的土壤養分及化學計量比進行比較;利用One-Way ANOVA對不同林齡梭梭生長特征、土壤SOC、TN、TP、Available P、CaCO3、C∶N、C∶P、N∶P、CaCO3∶Available P和風蝕可蝕性的比較;SOC、TN和TP的相關性用Linear regression analysis進行分析; 利用Person product-moment correlation analysis對土壤變量和影響因子進行相關性分析。所有數據均經過K-S檢驗(One-Sample Kolmogorv-Smirnov Test)。
3結果與分析
3.1不同林齡土壤養分成層化分布特征
如表1所示,SOC和TN含量在5a之前隨土壤深度增加無顯著變化趨勢(P>0.05),在5a之后表層(0—5 cm)SOC和TN含量則顯著高于5—20 cm的含量(P<0.05)。TP含量在13a之后0—5cm含量顯著高于5—20 cm含量(P<0.05)。表層Available P含量在2a之后表現為顯著高于深層(P<0.05),而CaCO3含量并未隨林齡增加表現出明顯的成層化分布(P>0.05)。
土壤C∶N在2、16、31a均表現為0—5 cm顯著高于5—20 cm(P<0.05),其余林齡下無顯著差異(P>0.05)。垂直方向上C∶P和N∶P變化趨勢一致,即在5a之后表層顯著高于深層(P<0.05)。CaCO3∶Available P則在不同林齡處(除0a)均表現為深層比值顯著高于表層(P<0.05)(表2)。
3.2不同林齡土壤養分水平分布化學計量特征
隨梭梭種植年限的增加,水平方向上0—5 cm和5—20 cm的SOC和TN含量均表現為顯著的增加(P<0.05),而TP含量并未隨林齡的增加發生顯著變化(P>0.05)。另外,Available P和CaCO3含量僅在土壤表層隨林齡增加顯著增加(P<0.05)且Available P的最大值(0.016 mg/g)僅占TP(0.36 mg/g)的5%(表2)。
水平方向上,隨梭梭林齡的增加,土壤C∶N和CaCO3∶Available P在0—5 cm和5—20 cm均未有顯著變化(P>0.05);C∶P和N∶P在表層則隨林齡顯著增加(P<0.05),5—20 cm處的C∶P和N∶P在31a之前均未有顯著變化,而在39a處達到最大值(表2)。
3.3土壤養分相關性及其與影響因子的關系
分析不同林齡土壤SOC, TN及TP的相關關系發現,三者均呈顯著的正相關關系(P<0.001)(圖2)。Pearson相關分析表明,土壤pH與SOC,TN,C∶P和N∶P均呈極顯著的正相關關系(P<0.01);土壤電導率(Electrical conductivity, EC)與SOC,TN, C∶P和N∶P均呈極顯著的正相關關系(P<0.01),與TP和Available P呈顯著的相關關系(P<0.05);除與CaCO3呈顯著的負相關關系(P<0.05),與CaCO3∶Available P呈顯著的正相關關系(P<0.05)及與C∶N無顯著關系(P> 0.05)外,土壤砂粒與其他土壤變量均呈極顯著的負相關關系(P<0.01);土壤粉粒和粘粒均與SOC,TN,TP,C∶P,N∶P呈極顯著的正相關關系(P<0.01),與CaCO3呈顯著的正相關關系(P<0.05),與CaCO3∶Available P呈顯著的負相關關系(P<0.05)。另外,Available P 與SOC,TN和粉粒呈極顯著的正相關關系(P<0.01),與粘粒呈顯著正相關(P<0.05),且SOC與CaCO3,N∶P,和CaCO3∶Available P均有顯著的相關關系(P<0.05)。總體來看,土壤砂粒、粉粒和粘粒是影響土壤養分及其化學計量特征的主要因子(表3)。
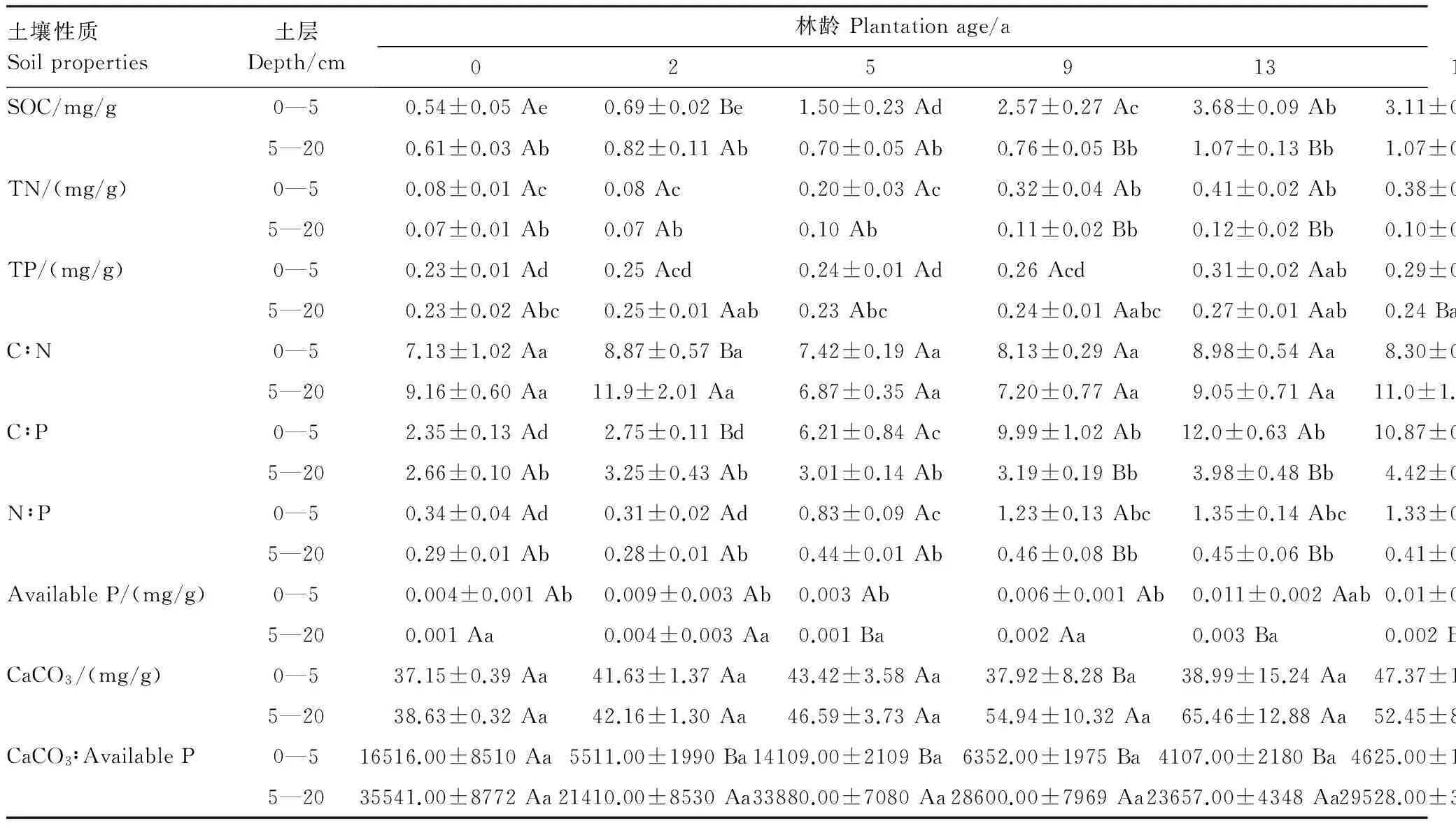
表2 不同林齡土壤養分成層化分布特征
同列同一元素不同大寫字母表示差異顯著(P<0.05);同行同一元素不同小寫字母表示差異顯著(P<0.05)
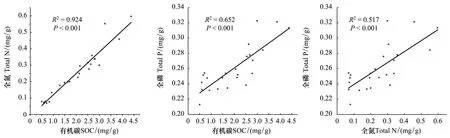
圖2 土壤有機碳、全氮和全磷的關系Fig.2 Relationship between SOC, TN, and TP
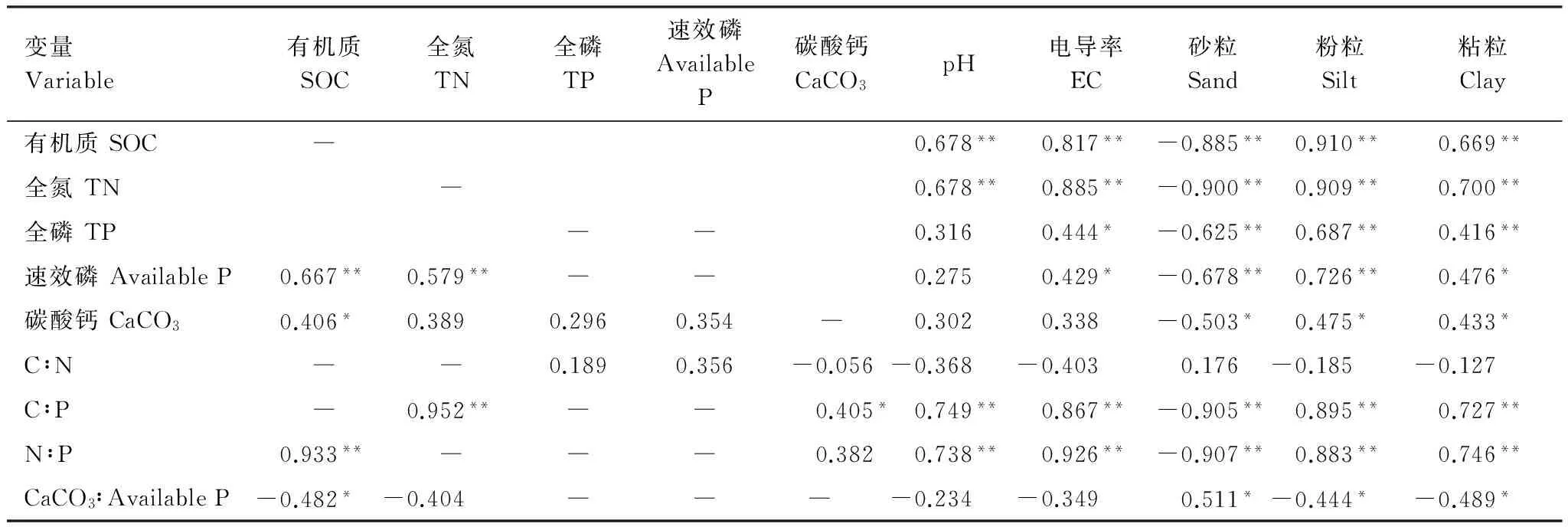
變量Variable有機質SOC全氮TN全磷TP速效磷AvailableP碳酸鈣CaCO3pH電導率EC砂粒Sand粉粒Silt粘粒Clay有機質SOC―0.678**0.817**-0.885**0.910**0.669**全氮TN―0.678**0.885**-0.900**0.909**0.700**全磷TP――0.3160.444*-0.625**0.687**0.416**速效磷AvailableP0.667**0.579**――0.2750.429*-0.678**0.726**0.476*碳酸鈣CaCO30.406*0.3890.2960.354―0.3020.338-0.503*0.475*0.433*C∶N――0.1890.356-0.056-0.368-0.4030.176-0.185-0.127C∶P―0.952**――0.405*0.749**0.867**-0.905**0.895**0.727**N∶P0.933**―――0.3820.738**0.926**-0.907**0.883**0.746**CaCO3∶AvailableP-0.482*-0.404――― -0.234-0.3490.511*-0.444*-0.489*
**表示極顯著相關(P<0.01); *表示顯著相關(P<0.05); ― 表示存在自相關,不宜進行相關性分析
3.4土壤風蝕可蝕性與土壤養分的關系
結果表明,隨林齡的增加,土壤表層(0—5 cm)的風蝕可蝕性極顯著的減弱,并且表層SOC、TN、TP、Available P和CaCO3含量均隨土壤風蝕可蝕性的減弱而顯著增加(P≤ 0.001)(表4,圖3)。而5—20 cm深度的土壤風蝕可蝕性呈顯著的減弱(P<0.05)(表4),且僅與土壤CaCO3含量有關(表5)(P= 0.001)。

表4 土壤風蝕可蝕性(EF)隨林齡變化特征
同列同一元素不同小寫字母表示差異顯著(P<0.05)
4討論
植被-土壤是一個相互作用的過程,土壤SOC、N、P元素作為植物正常生長發育所必需的養分,在植物生長過程中發揮著重要的作用,其含量的多少及成分組合狀況,均會受到土壤養分含量的影響,而植被的建立對土壤環境也有一定的反饋作用,如:植物凋落物的分解、根系分泌物及根際微生物的積累等,對土壤質地的改善和土壤養分的積累有極大的促進作用[26-29]。
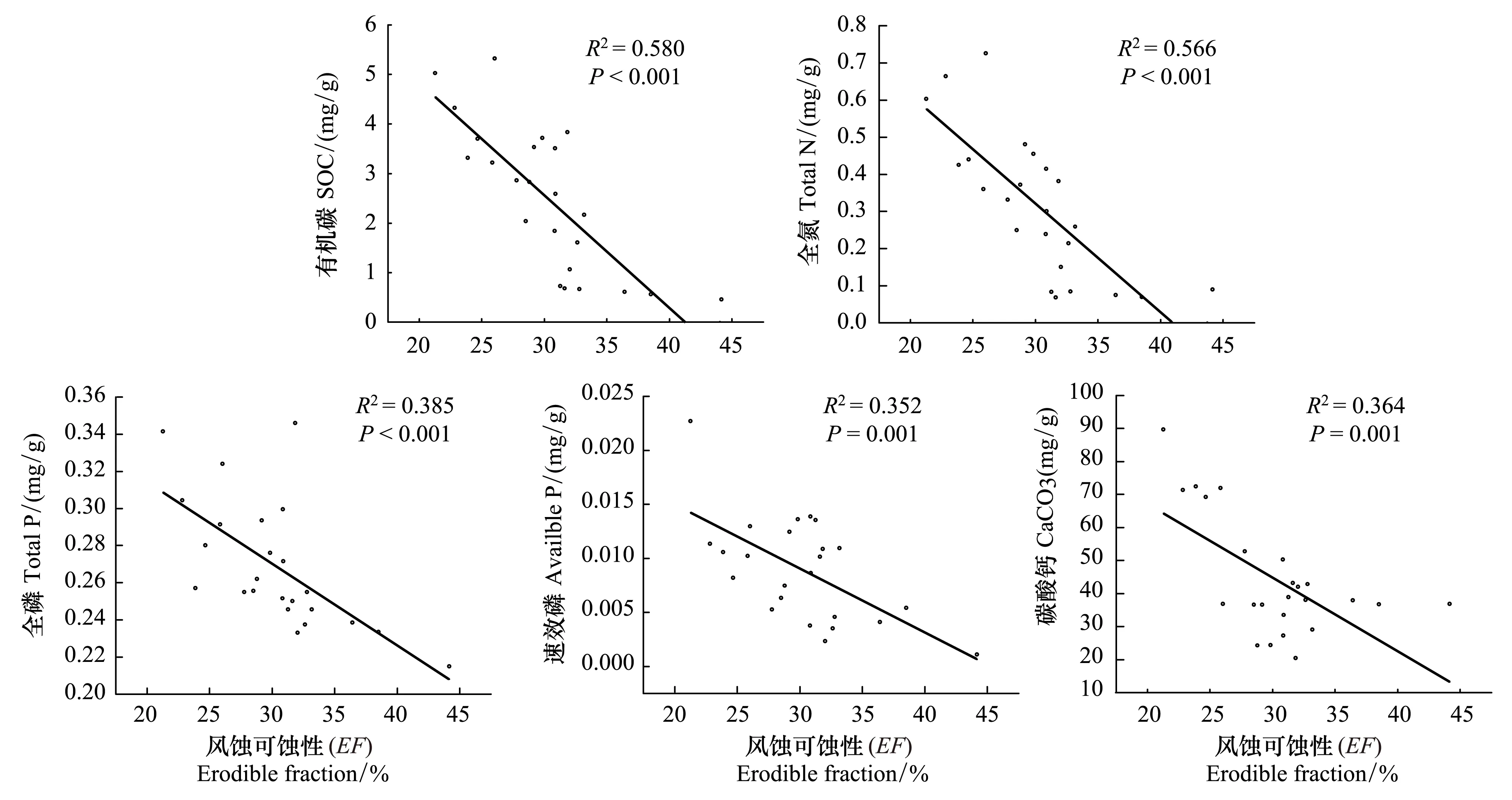
圖3 土壤風蝕可蝕性 (0—5 cm) 與土壤養分 (0—5 cm) 的關系Fig.3 Relationship between soil erodible fraction with soil nutrients in 0—5 cm
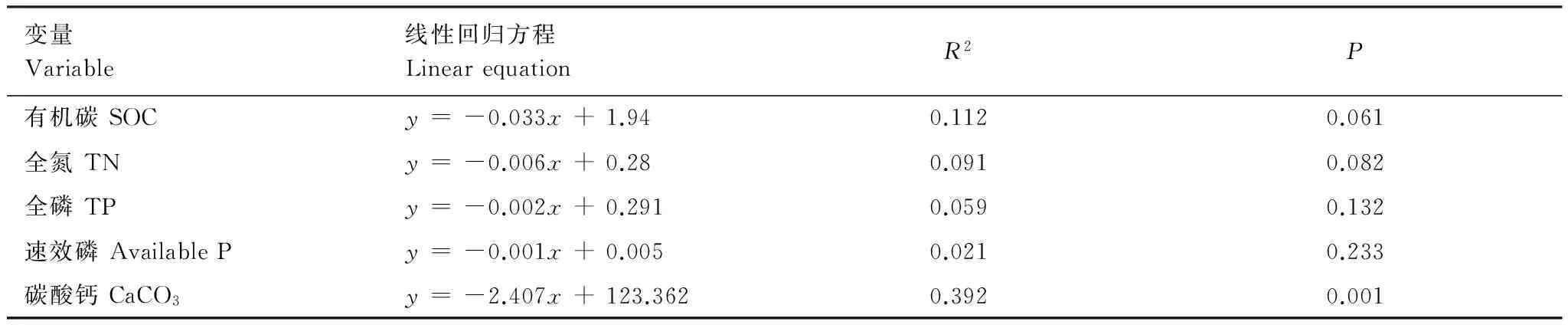
變量Variable線性回歸方程LinearequationR2P有機碳SOCy=-0.033x+1.940.1120.061全氮TNy=-0.006x+0.280.0910.082全磷TPy=-0.002x+0.2910.0590.132速效磷AvailablePy=-0.001x+0.0050.0210.233碳酸鈣CaCO3y=-2.407x+123.3620.3920.001
與流動沙丘相比,隨著梭梭的建立,SOC和TN均顯著增加并在梭梭建立5a后0—5 cm土壤SOC和N顯著高于5—20 cm土壤,一方面說明梭梭建立后能夠降低風速和減少地面的粗糙程度,不斷的改善土壤表面的物理性質,為土壤養分的積累提供必要條件,并且隨著梭梭林齡的增加,凋落物的不斷增加直接影響著SOC和TN的積累;另一方面也說明了在受水分限制的干旱區土壤養分的表層積聚作用[30-32]。相對于SOC和TN,TP含量隨梭梭林齡的增加沒有顯著變化且直到13a后0—5 cm TP含量才顯著高于5—20 cm,這是由于TP的增加主要是通過來源相對固定且時間漫長的巖石風化作用,且表層土壤更易受巖石風化的影響,這就導致土壤P成層化分布的形成相對滯后[11]。 隨林齡的增加,SOC和TN在0—5 cm和5—20 cm 均顯著增加,而TP并未發生顯著的變化,一方面說明梭梭的建立為SOC和TN的積累創造了有利的環境條件,而TP的變化則直接受土壤恢復時間的影響;另一方面表明梭梭的生長可能受P的限制。
C∶N∶P是衡量土壤有機質組成和營養平衡的一個重要指標,是確定土壤C、N、P平衡特征的重要參數[7,33]。與SOC和TN相比,土壤C∶N在不同深度和不同林齡間均維持相對穩定,這與Tian等[8]對中國土壤C∶N∶P的研究相一致[8],即SOC和TN含量具有較大的空間變異性,而受氣候的影響較小的C∶N相對穩定,同時驗證了不同深度土壤C∶N相對穩定的基本原則,即有機物質的形成需要一定數量的N與其相應的相對固定比率的C[2]。另外,相比于穩定的C∶N,土壤C∶P和N∶P隨林齡的增加而顯著增加,這與Tian等[8]、Cleveland等[7]分別從區域尺度和全球尺度上探討的天然生態系統土壤C∶N、C∶P和N∶P化學計量比得出的結論一致,即:由于C元素和P元素來源的差異性,土壤C∶P和N∶P隨氣候、土壤類型、土壤深度和風化階段而呈現顯著的差異性。同時表明天然生態系統土壤C∶N、C∶P和N∶P的變化特征也適用于研究區人工梭梭林土壤。
對C、N和P的相關性分析顯示三者間均有極顯著的相關性,且C和N、C和P、N和P的相關系數分別為0.92、0.65和0.52。這與Tian等[8]對中國土壤8000個土壤層C、N和P的相關分析結果(相關系數分別為0.93、0.62和0.51)相似,這說明相對于大尺度上有機質豐富的土壤,人工梭梭林建立后SOC含量顯著增加,且逐漸恢復的土壤中C、N和P也存在顯著的耦合關系,這與最近的研究相符[34-35],表明小尺度上研究C、N和P的耦合關系也具有重要的生態指示作用。另外,人工梭梭林建立后土壤養分及化學計量特征與土壤砂粒、粉粒和粘粒顯著的相關關系,進一步表明土壤質地的改善促進了土壤養分含量的提高[14]。
由于鈣離子在植物抗旱性中起重要作用,同時速效磷是植物直接可吸收利用的磷素直接影響著植物體蛋白質的合成[1-2,36-37],因此,土壤CaCO3和速效磷對植物生長有重要的影響。然而,研究表明[38],土壤中高含量的CaCO3會大大降低土壤磷的有效性,進而影響植物生長。并且衡量土壤碳酸鈣對速效磷固持指標的土壤碳酸鈣/速效磷(鈣磷比)化學計量特征對植物生物量、密度變化的影響顯著大于土壤碳酸鈣、速效磷含量的影響。本研究中,隨林齡增加,表層Available P顯著增加且在0—5 cm和5—20 cm間表現出明顯的成層化現象,而Available P的最大值(0.016 mg/g)僅占總磷 (0.36 mg/g)的5%,說明盡管梭梭的建立提高了土壤Available P的含量,而較低的Available P含量進一步說明梭梭的生長受P的限制。此外,CaCO3在表層隨林齡的增加而增加,而CaCO3∶Available P隨林齡增加并未發生顯著變化,說明土壤CaCO3含量與Available P增加的同步性,從而使得土壤中有限的速效磷被吸附固持。
土壤風蝕是導致干旱半干旱區土壤退化的一個普遍存在的現象[39]。土壤風蝕可蝕性臨界值顯示[24-25],EF<40%表明研究區較弱的風蝕可行性,40%
本研究在一定程度上揭示了梭梭建立對荒漠綠洲區土壤恢復的促進作用,為該區生態恢復提供了理論依據和參考。然而,植被-土壤是一個交互作用的復雜生態系統,因此,梭梭如何在有限的水分條件下實現可持續發展、梭梭群落特征及草本物種如何響應土壤環境變化將是今后的研究重點。
參考文獻(References):
[1]Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L J. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540- 550.
[2]Sterner R W, Elser J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton, New Jersey, USA∶Princeton University Press, 2002.
[3]?gren G I. The C∶N∶P stoichiometry of autotrophs-theory and observations. Ecology Letters, 2004, 7(3): 158- 191.
[4]Koerselman W, Meuleman A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441- 1450.
[5]Rong Q Q, Liu J T, Cai Y P, Lu Z H, Zhao Z Z, Yue W C, Xia J B. Leaf carbon, nitrogen and phosphorus stoichiometry ofTamarixchinensisLour in the Laizhou Bay coastal wetland, China. Ecological Engineering, 2015, 76: 57- 65.
[6]Chadwick O A, Derry L A, Vitousek P M, Huebert B J, Hedin L O. Changing sources of nutrients during four million years of ecosystem development. Nature, 1999, 397(6719): 491- 497.
[7]Cleveland C C, Liptzin D. C∶N∶P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 2007, 85(3): 235- 252.
[8]Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C∶N∶P ratios in China′s soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139- 151.
[9]He M Z, Dijkstra F A. Drought effect on plant nitrogen and phosphorus: a meta-analysis. New Phytologist, 2014, 204(4): 924- 931.
[10]Drenovsky R E, Richards J H. Critical N∶P values∶Predicting nutrient deficiencies in desert shrublands. Plant and Soil, 2004, 259(1/2): 59- 69.
[11]Delgado-Baquerizo M, Maestre F T, Gallardo A, Bowker M A, Wallenstein M D, Quero J L, Ochoa V, Gozalo B, Garcia-Gómez M, Soliveres S, García-Palacios P, Berdugo M, Valencia E, Escolar C, Arredondo T, Barraza-Zepeda C, Bran D, Carreira J A, Chaieb M, Concei??o A A, Derak M, Eldridge D J, Escudero A, Espinosa C I, Gaitán J, Gabriel Gatica M, Gómez-González S, Guzman E, Gutiérrez J R, Florentino A, Hepper E, Hernández R M, Huber-Sannwald E, Jankju M, Liu J S, Mau R I, Miriti M, Monerris J, Naseri K, Noumi Z, Polo V, Prina A, Pucheta E, Ramírez E, Ramírez-Collantes D A, Rom?o R, Tighe M, Torres D, Torres-Díaz C, Ungar E D, Val J, Wamiti W, Wang D L, Zaady E. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature, 2013, 502(7473): 672- 676.
[12]李新榮, 趙洋, 回嶸, 蘇潔瓊, 高艷紅. 中國干旱區恢復生態學研究進展及趨勢評述. 地理科學進展, 2014, 33(11): 1435- 1443.
[13]李新榮. 干旱沙區土壤空間異質性變化對植被恢復的影響. 中國科學(D輯:地球科學), 2005, 35(4): 361- 370.
[14]蘇永中, 趙哈林, 張銅會, 李玉霖. 科爾沁沙地不同年代小葉錦雞兒人工林植物群落特征及其土壤特性. 植物生態學報, 2004, 28(1): 93- 100.
[15]翁伯琦, 鄭祥洲, 丁洪, 王煌平. 植被恢復對土壤碳氮循環的影響研究進展. 應用生態學報, 2013, 24(12): 3610- 3616.
[16]Su Y Z, Zhao W Z, Su P X, Zhang Z H, Wang T, Ram R. Ecological effects of desertification control and desertified land reclamation in an oasis-desert ecotone in an arid region: A case study in Hexi Corridor, northwest China. Ecological Engineering, 2007, 29(2): 117- 124.
[17]Ma Q L, Wang J H, Zhu S J. Effects of precipitation, soil water content and soil crust on artificialHaloxylonammodendronforest. Acta Ecologica Sinica, 2007, 27(12): 5057- 5067.
[18]Zhu Y J, Jia Z Q. Soil water utilization characteristics ofHaloxylonammodendronplantation with different age during summer. Acta Ecologica Sinica, 2011, 31(6): 341- 346.
[19]Song J, Feng G, Tian C Y, Zhang F S. Strategies for adaptation ofSuaedaphysophora,HaloxylonammodendronandHaloxylonpersicumto a saline environment during seed-germination stage. Annals of Botany, 2005, 96(3): 399- 405.
[20]von Wandruszka R. Phosphorus retention in calcareous soils and the effect of organic matter on its mobility. Geochemical Transactions, 2006, 7: 6.
[21]Cross A F, Schlesinger W H. Biological and geochemical controls on phosphorus fractions in semiarid soils. Biogeochemistry, 2001, 52(2): 155- 172.
[22]陳隆亨, 李福興, 邸醒民, 張繼賢. 中國風沙土. 北京: 科學出版社, 1998.
[23]鮑士旦. 土壤農化分析(第三版). 北京: 中國農業出版社, 2000.
[24]Borrelli P, Ballabio C, Panagos P, Montanarella L. Wind erosion susceptibility of European soils. Geoderma, 2014, 232- 234: 471- 478.
[25]Fryrear D W, Krammers C A, Williamson D L, Zobeck T M. Computing the wind erodible fraction of soils. Journal of Soil and Water Conservation, 1994, 49(2): 183- 188.
[26]McGroddy M E, Daufresne T, Hedin L O. Scaling of C∶N∶P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios. Ecology, 2004, 85(9): 2390- 2401.
[27]賈曉紅, 李新榮, 王新平, 樊恒文, 趙金龍. 流沙固定過程中土壤性質變異初步研究. 水土保持學報, 2003, 17(4): 46- 50.
[28]蘇永中, 趙哈林, 張銅會. 幾種灌木、半灌木對沙地土壤肥力影響機制的研究. 應用生態學報, 2002, 13(7): 802- 806.
[29]李從娟, 雷加強, 高培, 徐新文, 王永東, 范敬龍. 人工防護林作用下風沙土成土過程的研究進展. 土壤學報, 2012, 49(6): 1227- 1234.
[30]Jobbágy E G, Jackson R B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry, 2001, 53(1): 51- 77.
[31]He M Z, Dijkstra F A, Zhang K, Li X R, Tan H J, Gao Y H, Li G. Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Scientific Reports, 2014, 4: 6923.
[32]Schlesinger W H, Reynolds J F, Cunningham G L, Huenneke L F, Jarrell W M, Virginia R A, Whitford W G. Biological feedbacks in global desertification. Science, 1990, 247(4946): 1043- 1048.
[33]楊陽, 劉秉儒, 楊新國, 韓叢叢. 荒漠草原中不同密度人工檸條灌叢土壤化學計量特征. 水土保持通報, 2014, 34(5): 67- 73.
[34]王維奇, 王純, 曾從盛, 仝川. 閩江河口不同河段蘆葦濕地土壤碳氮磷生態化學計量學特征. 生態學報, 2012, 32(13): 4087- 4093.
[35]周正虎, 王傳寬, 張全智. 土地利用變化對東北溫帶幼齡林土壤碳氮磷含量及其化學計量特征的影響. 生態學報, 2015, 35(20): 6694- 6702.
[36]Larkindale J, Knight M R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiology, 2002, 128(2): 682- 695.
[37]Kawano T, Sahashi N, Takahashi K, Uozumi N, Muto S. Salicylic acid induces extracellular superoxide generation followed by an increase in cytosolic calcium ion in tobacco suspension culture: The earliest events in salicylic acid signal transduction. Plant and Cell Physiology, 1998, 39(7): 721- 730.
[38]趙雪萊, 何興東, 薛蘋蘋, 張寧, 鄔畏, 李榮, 慈華聰, 徐靜靜, 高玉葆, 趙哈林. 土壤碳酸鈣/有效磷化學
[39]南嶺, 杜靈通, 展秀麗. 土壤風蝕可蝕性研究進展. 土壤, 2014, 46(2): 204- 211.
重慶縉云山兩種林分土壤呼吸對模擬氮沉降的季節響應差異性
基金項目:國家自然科學基金項目(41271291);國家林業局“948”項目(2015- 4- 42);重慶市林業重點科技攻關項目(渝林科研2015- 7);重慶市應用開發計劃重點項目(CSTC2014yykfB80015);西南大學資源環境學院“光炯”創新實驗項目(2計量特征對油蒿群落植物密度的影響. 科學通報, 2012, 57(1): 80- 87.
0150104)
曾清蘋,何丙輝,毛巧芝,秦華軍,李源,黃祺.重慶縉云山兩種林分土壤呼吸對模擬氮沉降的季節響應差異性.生態學報,2016,36(11):3244- 3252.
Zeng Q P, He B H, Mao Q Z, Qin H J, Li Y, Huang Q.Seasonal responses of soil respiration to simulated nitrogen deposition in a citrus plantation and masson pine forest in Mt. Jinyun, Chongqing, China.Acta Ecologica Sinica,2016,36(11):3244- 3252.
重慶縉云山兩種林分土壤呼吸對模擬氮沉降的季節響應差異性
曾清蘋1,何丙輝1,*,毛巧芝1,秦華軍2,李源3,黃祺4
1 西南大學資源環境學院,三峽庫區生態環境教育部重點實驗室,重慶400715 2 貴州省生物研究所,貴陽550009 3 重慶市涪陵區林業局,重慶408000 4 中國電建集團貴陽勘測設計研究院有限公司,貴陽550009
摘要:氮沉降對土壤呼吸的影響仍然存在著爭論,需要進一步研究。選擇重慶縉云山的馬尾松林和柑橘林開展了氮添加實驗,分別設置3個氮添加水平(低氮T5:20 gN m-2a-1,中氮T10:40 gN m-2a-1和高氮T15:60 gN m-2a-1)和對照(T0:0 gN m-2a-1)共4個水平的處理,各林分每個處理各9次重復,每個處理量分4次,在每個季度開始各施1次。采用ACE(Automated Soil CO2Exchange Station, UK)自動土壤呼吸監測系統測定兩林分土壤表層(0—10 cm)的呼吸、溫度和濕度,分別在當年的7月、9月、11月、第2年的1月、2月、3月、5月、6月各連續測定4d,每天(8:00—18:00)4次,以揭示兩種林分土壤呼吸對模擬氮沉降的季節動態響應及其差異性。結果表明:(1)柑橘林與馬尾松林林下土壤表層呼吸表現出一致的季節變化動態趨勢:夏季>春季>秋季>冬季,但柑橘林土壤呼吸顯著高于馬尾松林(P<0.05)。(2)總體上氮沉降抑制了2種林分土壤表層呼吸,而N沉降量大抑制程度越高。只在冬季土壤濕度低的馬尾松林下氮沉降促進了土壤呼吸。(3)土壤溫度與土壤呼吸有極顯著的正相關指數關系(P<0.01),而土壤水分與土壤呼吸有顯著的二次模型擬合關系,但均受到氮沉降量處理的影響。綜合分析表明,在亞熱帶山區2類森林下的典型案例結果支持氮沉降抑制土壤呼吸的認識。
關鍵詞:氮沉降;土壤呼吸;土壤溫度;土壤濕度;柑橘林;馬尾松林
Abstract: Disagreements over the effects of nitrogen deposition on soil respiration still exist, thereby necessitating further studies. In the present study, we designed an experiment to explore the seasonal responses of soil respiration to various levels of nitrogen deposition (T0: 0 g N m-2a-1as the control; T5: 5 g N m-2a-1as the low N treatment; T10: 10 g N m-2a-1as the moderate nitrogen treatment; and T15: 15 g N m-2a-1as the high nitrogen treatment. All treatments were performed with nine replicates both in the Masson pine forest and in the Citrus plantation at Mt. Jinyun, Chongqing, China. The different levels of N addition were applied four times at the beginning of each season from May 2014. We measured soil respiration, soil temperature, and soil moisture simultaneously by using the Automated Soil CO2Exchange Station (ACE, UK) for four consecutive days in July, September, November, January, February, March, May, and June, respectively, at four times each day between 8:00 and 6:00. The results showed that:(1) the two forests underwent similar seasonal changes in soil respiration, with the most change being observed during the summer, followed by spring, autumn, and winter, which yielded the least change. However, the citrus plantation presented significantly higher levels of soil respiration than the Masson pine forest across all seasons (P< 0.05). (2) Generally, nitrogen deposition suppressed soil respiration in both forests and these inhibitory effects were strengthened with increasing levels of nitrogen deposition. The only exception in which nitrogen deposition enhanced soil respiration was in the Masson pine forest during the winter, while soil moisture was very low. (3) Soil respiration showed significantly positive exponential relationships with soil temperature (P< 0.01) and significant quadratic relationship with soil moisture (P< 0.05). Both factors (soil temperature and moisture) are influenced by the levels of nitrogen deposition in subtropical forests. Thus, the results of our study on subtropical forests confirm the theory that nitrogen deposition inhibits soil respiration.
Key Words: nitrogen deposition; soil respiration; soil temperature; soil moisture; citrus plantation; Masson pine forest
近年來,全球變暖逐漸成為各界關注的熱點問題,IPCC第5次評估報告(AR5)指出2003—2012年平均溫度較1850—1900年上升0.78℃,并表現出加劇的趨勢。CO2作為導致全球變暖最主要的氣體之一,2011年其濃度達到391μL/L比1750年高40%[1- 2]。而土壤呼吸(Rs)是大氣CO2的重要來源,約占整個生態系統呼吸的50%—80%,主要由土壤動物、微生物和根系等呼吸產生[3- 4],是陸地生態系統中最重要的碳循環過程[5],其產生速度受自然、生物以及各種干擾等因子影響[6]。
隨著人類社會發展,氮肥施用、畜牧業快速發展及礦物燃料燃燒等活動使大氣含氮化合物增多,導致大氣氮沉降量加劇,且變化量有增無減[7]。氮素作為林木生長的養分限制因子,它直接或間接影響著森林土壤呼吸。目前,氮沉降引起的土壤呼吸改變逐漸成為各領域學者研究的重點,其中最常用的方法為外源氮添加模擬氮沉降試驗,然而研究區域、研究時間不同所得結論各異。吳迪等[8]、劉益君等[9]、孫素琪等[10]及Sun等[11]模擬氮沉降試驗表明氮沉降對土壤呼吸有抑制作用,Tu等[12]的研究結果表明氮沉降對土壤呼吸有促進作用,Hasselquist等[13]則發現氮沉降并沒有明顯改變林地土壤呼吸。可見,氮沉降對土壤呼吸的影響存在很大的不確定性,需要進一步的深入研究。氮沉降的土壤效應深受植被性質、土壤類型與氣候差異的影響[8-13],不同的土壤水熱環境也會使土壤呼吸有較大差異。北美[14]、歐洲[15]關于氮沉降試驗開展較早,而國內氮沉降對森林土壤呼吸影響的試驗起步較晚,且大部分集中在溫帶森林系統,涉及熱帶、亞熱帶森林相關研究相對較少。
本文選取我國亞熱帶地區廣泛分布的馬尾松和柑橘林為研究對象,開展人工氮添加模擬氮沉降試驗。探討不同氮沉降水平對馬尾松林和柑橘林土壤呼吸及水熱環境的影響,探索該區域馬尾松林和柑橘林土壤呼吸的動態特征以及不同林分對氮沉降的響應,為不同林地利用途徑的土壤呼吸研究提供基礎數據。
1材料與方法
1.1研究區概況
研究區位于重慶市北碚區境內縉云山國家級自然保護區(106°22′E,29°45′N)內,距市中心60 km。縉云山占地面積76 km2,海拔350—951.5 m,屬典型中亞熱帶溫暖濕潤季風氣候。年平均溫度13.6℃,最熱月(8月)平均溫度24.3℃,最高溫43℃,最冷月(1月)平均溫度3.1℃,最低溫-1℃;年平均相對濕度87%,年平均降雨量1161.8 mm,年均日照1293.9 h,干濕季分明。雨季是4—9月(降雨量1243.8 mm),其中7—8月是伏旱期(蒸發量255.4 mm),降雨量小,蒸發量大。土壤以酸性黃壤及水稻土為主,伴有少量零星分布的紫色土。區內植物資源豐富,植被種類繁多,森林覆蓋率達96.6%。試驗地建立在海拔為350—500 m的柑橘林和馬尾松林內,其中柑橘林林下無灌木層,草本層為優勢群落,主要優勢草本為:兔兒酸(PolygonumamphibiumL.)、糯米團(Gonostegiahirta(Bl.) Miq.)、九層塔(OcimumbasilicumL.)、加拿大飛蓬(Conyzacanadensis(L.) Cronq.)、杠板歸(PolygonumperfoliatumL.)、稗草(Echinochloacrusgalli(L.) Beauv.);馬尾松林主要優勢灌木為:杉木(Cunninghamialanceolata(Lamb.) Hook.)、鹽膚木(RhuschinensisMill.)、四川山礬(SymplocossetchuensisBrand)、異葉榕(FicusheteromorphaHemsl.)、青岡(Cyclobalanopsisglauca(Thunb) Oersted)、野桐(Mallotusjaponicus(Thunb.) Muell. Arg.)、毛桐(Mallotusbarbatus(Wall.) Muell. Arg.)、樸樹(CeltissinensisPers.)、黃牛奶樹(Symplocoslaurina(Retz.) Wall.)、鈍葉柃(EuryaobtusifoliaH. T. Chang)、白櫟(QuercusfabriHance)、慈竹(Neosinocalamusaffinis(Rendle) Keng f.)等,主要優勢草本為:邊緣鱗蓋蕨(Microlepiamarginata(Houtt.) C. Chr.)、中華里白(Hicriopterischinensis(Ros.)Ching)、粽葉狗尾草(Setariapalmifolia(Koen.) Stapf)、菝葜(SmilaxchinaLinn.)、山莓(RubuscorchorifoliusLinn. f.)、尖葉清風藤(SabiaswinhoneiHemsl.)、香花崖豆藤(MillettiadielsianaHarms)等。
1.2實驗設計
2014年5月,在海拔相近的馬尾松林和柑橘林內分別選取3塊面積為20 m×20 m的代表性樣地,基本情況如表1所示。在每個樣地沿對角線確定3個點,于每個點各設置4個1 m×1 m的小樣地(間隔>1 m防相互干擾),作為氮添加處理(不考慮大氣氮沉降)樣方。氮添加處理設置了4個水平:對照(T0,0 gN m-2a-1)、低氮(T5,20 gN m-2a-1),中氮(T10,40 gN m-2a-1)和高氮(T15,60 gN m-2a-1)。為消除流水等對樣地氮含量造成影響,施氮時保證高濃度樣方在低濃度樣方之下。同時將長為16 cm內徑為23 cm的PVC管隨機埋入小樣地中并確保管頂距土壤表面3 cm,去除管內植物和凋落物。在每個季度初期選擇晴天將不同濃度NH4NO3溶于1L水中,采用手提式噴霧器均勻噴灑在每個小樣地內,為排除試驗中溶解NH4NO3所需水分對土壤呼吸的影響,在對照樣地噴灑了等體積水。
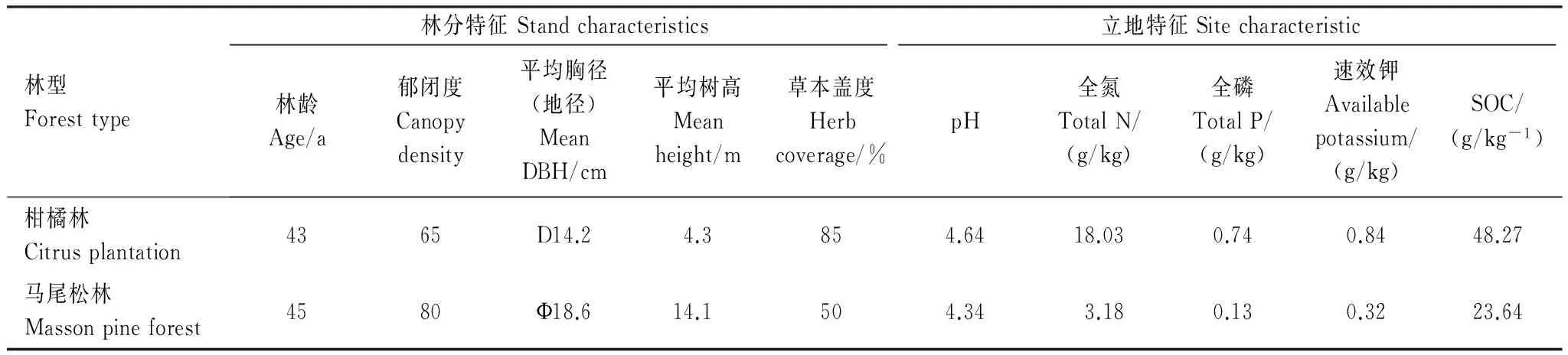
表1 重慶縉云山2種實驗林(馬尾松林和柑橘林)的林分與土壤特征基本信息
1.3土壤呼吸的測定
2014年7月至2015年7月,夏、秋、冬、春4個季節(夏季用7月和9月所得數據平均值表示,秋季用11月和1月所得數據平均值表示,冬季用2月和3月所得數據平均值表示,春季用5月和6月所得數據平均值表示),在施肥2d后(所施氮溶液完全滲入土壤后)選擇晴朗天氣,采用ACE自動土壤呼吸監測系統(Automated Soil CO2 Exchange Station, UK)于7月、9月、11月、1月、2月、3月、5月、6月對兩林地土壤呼吸進行測定,各測定4d(連續),各個點每天共測定4次,測定時間為8:00—18:00(每次測定間隔2.5 h)。為了減小安放PVC管對土壤呼吸的影響,每個季度埋管與測定土壤呼吸時間間隔保持一致,并在每次測定前一天,將測定管內的地表植被自土壤表層徹底剪除,盡量不破壞土壤,以減少土壤擾動和根系損傷對測量結果的影響。呼吸圈埋好后固定1a。
在測定土壤呼吸的同時用ACE自動土壤呼吸監測系統配帶的2個土壤溫度熱電阻探頭和2個土壤水分探頭SM300分別對土壤10cm深處的溫度和濕度進行同步測量。
1.4數據處理與統計分析
以每個點每天測定的4個數據通過算術平均來代表該林地的當日平均土壤呼吸、土壤溫度和土壤濕度。數據處理和統計分析在SPSS18.0、office Excel 2003、OriginPro9.0與MATLAB R2012B等軟件中進行,采用單因素方差分析法(LSD)比較土壤呼吸在不同氮處理、不同季節間的差異水平,采用回歸分析方法描述土壤呼吸與溫度(T)及濕度(W)的關系。
土壤呼吸與土壤溫度之間的關系采用指數模型[16]:Rs=R0eβT;Q10值通過下式計算[17- 18]:Q10=e10β;采用二次項模型[19]:Rs=a+b1W+b2W2與冪函數模型[20]:Rs=aWb研究土壤呼吸與土壤濕度的相關性;利用線性模型[21]:Rs=a+bT+cW和指數冪函數模型[22]:Rs=aebTWc研究土壤溫度和土壤濕度交互作用對土壤呼吸的影響。式中Rs為土壤呼吸μmol m-2s-1,T為地溫,R0是溫度為0℃時的土壤呼吸,β為溫度反應系數,Q10為土壤呼吸溫度敏感性指數,W為土壤濕度,a,b,c為待定參數。
2結果
2.1兩種林分土壤呼吸、土壤溫度和土壤濕度季節特征
實驗期內兩林分土壤呼吸和溫濕度隨季節均表現出先減后增的變化趨勢,且差異顯著(圖1)。T0為空白對照,其值反映了兩森林生態系統內土壤呼吸特征。柑橘林和馬尾松林土壤呼吸最大值出現在夏季,分別為(4.947±0.642)、(3.930±0.378)μmol m-2s-1,相應的地表土壤溫度最大值也出現在夏季,分別為(31.10±6.287)℃、(27.77±1.575)℃;而最小值則出現在冬季,此時兩林分的土壤呼吸分別為(2.868±0.695)μmol m-2s-1和(2.634±0.413)μmol m-2s-1,溫度則為(14.87±1.600)℃和(13.77±0.429)℃;柑橘林(43.902±3.985)%和馬尾松林(33.259±4.985)%土壤濕度最大值均出現在春季,最小值出現在秋季(20.087±1.590)%、(15.866±1.250)%(圖1,圖2)。
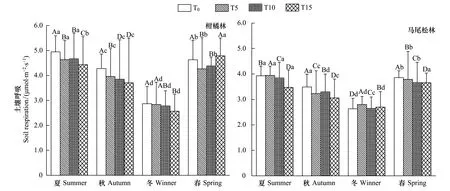
圖1 重慶縉云山馬尾松林和柑橘林在不同施氮處理下的土壤呼吸的季節動態變化Fig.1 Seasonal variation of soil respiration rate under the Masson pine forest and Citrus plantation with various nitrogen addition amount treatments at Mt. Jinyun, ChongqingP<0.05,大寫字母表示同一季節不同處理間差異性水平,小寫字母表示同一處理不同季節間差異性水平;T0:control check,對照組;T5:low nitrogen,低氮水平;T10:middle nitrogen,中氮水平;T15:high nitrogen,高氮水平
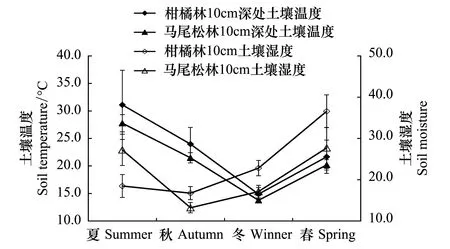
圖2 重慶縉云山柑橘林和馬尾松林下土壤的表層溫度及濕度的季節動態變化Fig.2 Seasonal variation of soil temperature and soil moisture under the Masson pine forest and nearby Citrus plantation at Mt. Jinyun, Chongqing
2.2兩種林分土壤呼吸對模擬氮沉降的響應
不同氮沉降處理的土壤呼吸具有一致的變化規律,表現為夏季最高,其次是春季和秋季,冬季最低,且各季度間差異顯著(圖1)。在春季,馬尾松林土壤呼吸隨著模擬的氮沉降量增加呈直線下降趨勢,T5、T10和T15處理分別比對照T0降低了1.76%、4.86%及5.11%,但柑橘林則表現為先降低后升高的變化趨勢,與對照T0相比,T5、T10和T15分別降低了7.87%、降低了5.36%、升高了3.38%;在夏季,馬尾松林土壤呼吸隨氮沉降量增加而表現為先升高后降低,T5、T10和T15處理分別比對照T0升高了0.44%、降低了2.11%和11.62%,但柑橘林土壤呼吸基本上呈降低趨勢,T5、T10和T15處理后的土壤呼吸均比對照T0低,且分別降低6.34%、5.68%及10.30%;在秋季,較對照T0而言,兩林分土壤呼吸在氮沉降處理后均降低,馬尾松林T5、T10和T15處理分別降低了7.44%、5.50%、12.36%,而柑橘林T5、T10和T15處理則分別降低7.43%、10.04%、12.34%;在冬季,氮沉降處理后馬尾松林T5、T10和T15處理土壤呼吸比對照T0高6.32%、0.43%和2.51%,而柑橘林T5、T10和T15處理土壤呼吸則比對照T0低0.96%、3.11%和10.52%。
2.3兩種林分土壤呼吸與土壤溫度的關系模型對模擬氮沉降的響應
各處理土壤呼吸與土壤溫度進行回歸分析(圖3)結果可知,土壤呼吸與土壤溫度存在極顯著的指數關系(P<0.01),柑橘林T0、T5、T10和T15處理與10 cm深處土壤溫度之間擬合方程的決定系數R2分別為0.613、0.358、0.358和0.377,解釋了柑橘林土壤呼吸35%—62%的變異,Q10值分別為1.36、1.35、1.33、1.40;馬尾松林擬合方程的決定系數R2分別為0.558、0.238、0.428和0.165,可解釋馬尾松林土壤呼吸16%—56%的變異,Q10值分別為1.32、1.25、1.35、1.19。
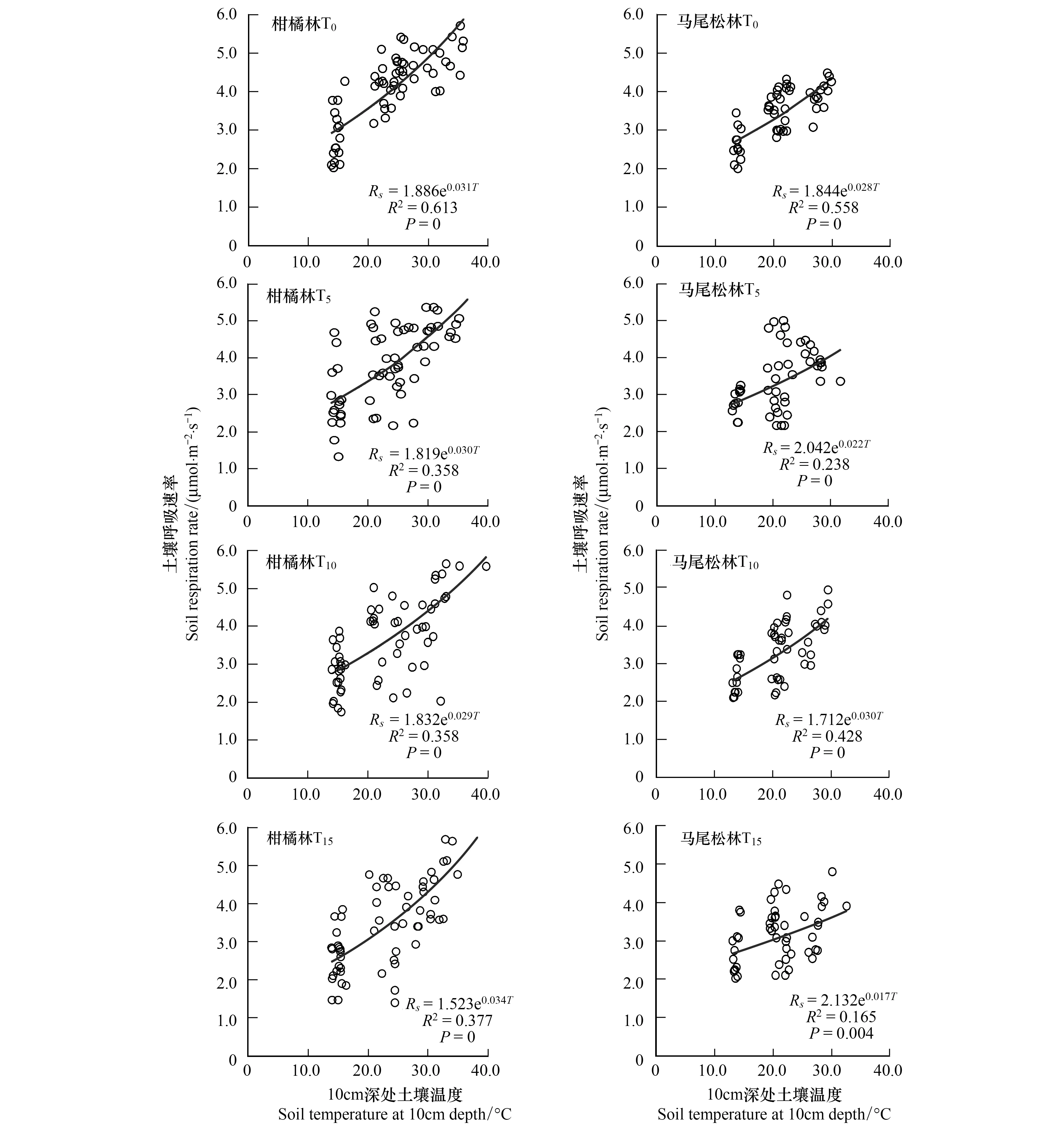
圖3 重慶縉云山柑橘林和馬尾松林在不同氮沉降量模擬條件下土壤10cm深處的呼吸與溫度的關系Fig.3 Relationship between soil respiration and soil temperature at 10 cm depth with various nitrogen addition amount treatments under the Masson pine forest and Citrus plantation from Mt. Jinyun, Chongqing
2.4兩種林分土壤呼吸與土壤濕度的關系模型對模擬氮沉降的響應
將不同處理的土壤呼吸與10 cm深處土壤濕度用二次模型和冪函數模型進行擬合,擬合度R2在0.052—0.233(表2),明顯低于土壤呼吸與溫度關系方程的R2值,且二次模型擬合效果優于冪函數模型。這些結果也說明,2種林分條件下的土壤濕度變化顯著影響著土壤的呼吸。
表2重慶縉云山柑橘林和馬尾松林下土壤表層在模擬不同氮沉降處理條件下的呼吸與濕度的關系方程
Table 2Correlation equations between soil respiration rate and soil moisture at 10cm depth across various nitrogen addition treatments under the Masson pine forest and Citrus plantation at Mt. Jinyun, Chongqing

林型Foresttype處理TreatmentRs=a+b1W+b2W2Rs=aWbab1b2PR2abPR2柑橘林T05.018-0.014-0.0010.0040.16611.110-0.3410.0020.147CitrusplantationT56.036-0.1610.0030.1060.0717.828-0.2480.0470.062T104.1160.019-0.0010.1370.0638.723-0.2930.0690.052T151.1570.156-0.0010.0140.1110.5040.6400.0000.172馬尾松林T03.543-0.0460.0020.0250.1482.2020.1450.0510.078MassonpineforestT54.104-0.1350.0050.0040.2161.5090.2680.0090.136T102.8420.0010.0010.0020.2331.5700.2510.0020.186T151.9380.086-0.0010.0060.1991.4140.2700.0010.204
2.5兩種林分土壤呼吸與土壤溫度、土壤濕度的復合關系模型對模擬氮沉降的響應
分別利用線性模型和指數冪函數模型對土壤呼吸與土壤溫度和土壤濕度的復合關系進行分析(表3)。兩林分T0處理的指數冪函數模型擬合效果不如線性模型,其他處理的指數冪函數模型效果均優于線性模型。從指數冪函數模型擬合結果可知,土壤溫度和濕度能夠共同解釋兩林分土壤呼吸季節變化的56.6%—61.4%。不同氮沉降處理下馬尾松和柑橘林土壤呼吸受土壤溫度和土壤濕度影響程度不一樣,兩林分T0擬合度R2均高于氮沉降T5、T10和T15組,氮沉降處理下土壤溫度和土壤濕度僅能解釋土壤呼吸28.0%—47.8%的變異。
3討論
土壤呼吸與植被類型、林下溫濕度、pH、土壤質地及有機質含量等密切相關[23- 25]。不同植被的凋落物質量、數量和纖維素含量各異,這些均可影響土壤中微生物和酶等分解凋落物的強度和時間[26]。植被不同,其葉面積指數、根生物量等均有差異,葉面積大小可影響植被覆蓋下土壤微型氣候,Sims等[27]發現土壤CO2通量值和同時測量的葉面積指數值有顯著的相關性。Althea等[28]在研究本寧堡松樹林時發現土壤呼吸與距離最近的松樹成負相關,與根生物量成正相關。本實驗中柑橘林土壤呼吸顯著高于馬尾松林,原因可能是柑橘林郁閉度低、凋落物輸入量大,柑橘樹根細、量多且分布廣,同時有機肥添加和采果、剪枝等人為活動促進了土壤養分積累與轉化,加之林下良好的光、熱、水等環境條件促進了土壤微生物和根系生長與代謝,增加了微生物活性與數量,加快了土壤有機質分解速度,因此產生CO2的速率加快。
表3重慶縉云山柑橘林和馬尾松林林下模擬氮沉降各處理情況下土壤呼吸與10cm深度溫度(T)及濕度(W)的復合關系方程
Table 3Combined relationships between soil respiration and soil temperature as well soil moisture at 10 cm depth across different nitrogen addition amount treatments under the Masson pine forest and Citrus plantation at Mt. Jinyun, Chongqing
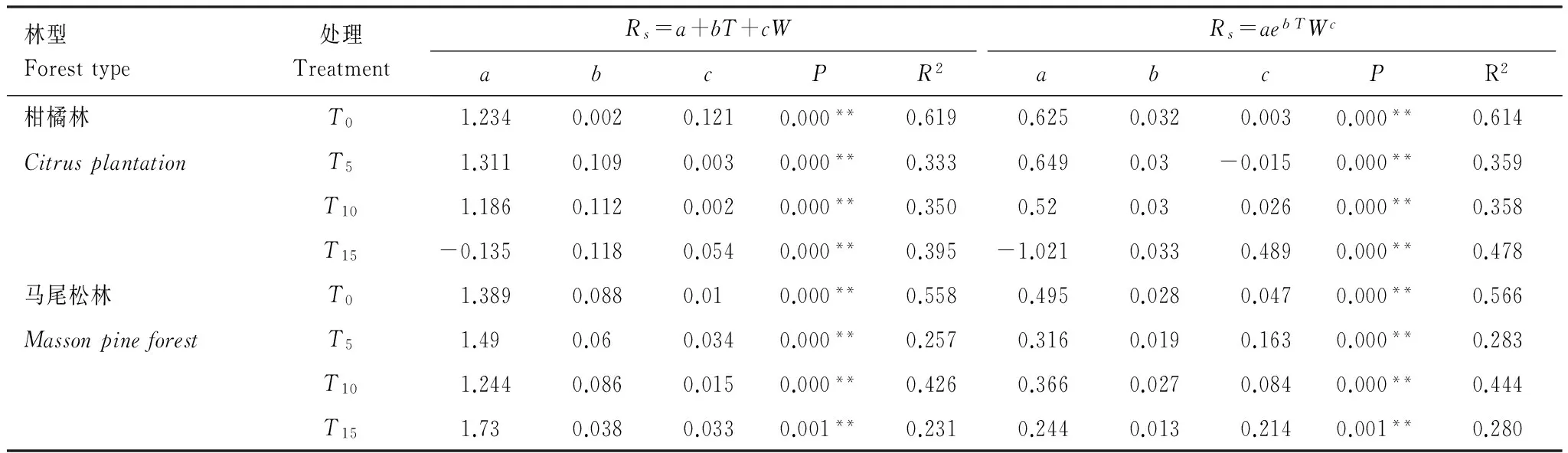
林型Foresttype處理TreatmentRs=a+bT+cWRs=aebTWcabcPR2abcPR2柑橘林T01.2340.0020.1210.000**0.6190.6250.0320.0030.000**0.614CitrusplantationT51.3110.1090.0030.000**0.3330.6490.03-0.0150.000**0.359T101.1860.1120.0020.000**0.3500.520.030.0260.000**0.358T15-0.1350.1180.0540.000**0.395-1.0210.0330.4890.000**0.478馬尾松林T01.3890.0880.010.000**0.5580.4950.0280.0470.000**0.566MassonpineforestT51.490.060.0340.000**0.2570.3160.0190.1630.000**0.283T101.2440.0860.0150.000**0.4260.3660.0270.0840.000**0.444T151.730.0380.0330.001**0.2310.2440.0130.2140.001**0.280
**表示在0.01顯著水平下的差異性;*表示在0.05顯著水平下的差異性
土壤呼吸有明顯的季節性變化,丁訪軍等[29]在研究喀斯特地區的灌木林、闊葉混交林和針闊混交林土壤呼吸時發現:土壤呼吸呈夏季>秋季>春季>冬季的變化規律。這與劉春霞等[30]研究結論一致。而本實驗結果為夏季>春季>秋季>冬季,春季>秋季的結論與丁訪軍、劉春霞等研究不一致,原因可能是研究對象、處理空間和測定時間的異質性導致的。實驗區夏季土壤呼吸最大,夏季光照充足、溫度較高、降雨量也最豐富,良好環境條件有利于植物生長發育,促進根系呼吸,同時加快凋落物分解,增加微生物種群數量及其代謝活性并使土壤中聚集較高的土壤酶活性,進而使土壤微生物呼吸、土壤動物呼吸及土壤中氧化還原反應產生CO2的速度加快。
氮沉降對森林土壤呼吸的影響主要有3種:促進、抑制和不顯著。總的來說,本實驗除冬季氮沉降促進馬尾松林土壤呼吸外,其余均表明氮沉降抑制土壤呼吸。這與國內外很多研究結果一致。比如,吳迪等[8]在研究長江中下游歐美楊(Populus dettoides)人工林時發現低氮、中氮和高氮處理使CO2排放量分別減少12.35%、14.45%和15.73%。劉益君等[9]發現,氮沉降顯著抑制了濕地松林分的土壤呼吸且抑制作用第1年顯著大于第2年,施氮處理后第1年低、中、高處理土壤呼吸分別比對照降低了31.3%、27.4%和24.5%,第2年則分別降低24.0%、16.5%和25.3%。孫素琪等[10]于2012年研究縉云山常綠闊葉林對模擬氮沉降的響應時發現低、中、高氮處理分別使土壤呼吸降低了20.88%,34.80%和42.23%。Sun等[11]在研究河北塞罕壩國家森林公園落葉松林土壤呼吸時發現低氮處理使土壤呼吸降低10.0%,高氮則降低12.5%,與本文結果基本一致但與冬季馬尾松林處理后所得結果相異。也有研究表明氮沉降對土壤呼吸有促進作用,Tu等[12]在研究竹林生態系統時發現氮沉降顯著增加了土壤呼吸。Hasselquist等[13]研究結果則表明氮沉降對土壤呼吸無影響。這可能跟植被、土壤性質、施氮處理方式、處理與測定時間間隔長度以及氣候條件等不同有關。
氮沉降使土壤呼吸降低的原因可能是:①氮沉降增加了土壤銨態氮和硝態氮含量,高濃度氨氮含量降低微生物活性和數量,產生抑制物質或影響土壤酶系統,從而阻礙凋落物分解和養分釋放,進而減慢土壤碳釋放速度。②氮沉降加強土壤硝化作用,使森林土壤pH值降低,加速土壤酸化[31],降低植物根系生長和根生物量,致使根際分泌物減少,進而抑制植物根系呼吸[10]。③在氮素豐富的地區,當森林土壤氮含量達到飽和狀態后,過量的氮沉降就會對土壤呼吸產生抑制的作用[32- 33]。
縉云山兩林分土壤表層呼吸受土壤溫度和濕度共同影響。圖4是基于指數冪函數模型的土壤呼吸對溫濕度協同變化的響應趨勢面。該響應趨勢面表明:第一,土壤呼吸隨溫度升高而加強;第二,低溫下,土壤呼吸受土壤濕度影響較小,高溫時,土壤呼吸則受溫度和濕度協同控制;第三,土壤濕度對馬尾松林土壤呼吸影響較明顯,但對柑橘林土壤呼吸影響較小。Zeng等[34]在研究太行山洋槐林、黃荊林及鐵桿蒿地時發現土壤呼吸主要受土壤溫度驅動而與土壤濕度僅有微弱的關系。Chen等[35]研究北京奧林匹克公園時發現土壤呼吸年周期主要受溫度控制,而夏季土壤呼吸主要受濕度控制,這均與本研究結論一致。說明土壤呼吸受土壤溫度影響更大,而多數情況下土壤呼吸受土壤溫度及濕度共同控制。

圖4 柑橘林和馬尾松林土壤呼吸的溫濕度響應趨勢面Fig.4 Response surface of soil respiration to soil temperature and soil moisture in Masson pine forest and Citrus plantation
4結論
(1)兩林分土壤呼吸均具有明顯的季節變化規律,其中夏季土壤呼吸最高,其次是春季和秋季,最低為冬季,且柑橘林土壤呼吸明顯高于馬尾松林,不同季度觀測的土壤呼吸平均值之間差異顯著(P<0.05)。
(2)總體而言,氮沉降抑制了兩林分土壤表層呼吸,抑制程度隨氮沉降增加而升高,只在土壤濕度低的冬季,氮沉降對馬尾松林土壤呼吸有促進作用。
(3)土壤呼吸與土壤溫度存在極顯著的指數關系,與土壤呼吸有顯著的二次模型擬合關系,但均受氮沉降量處理的影響。
[1]IPCC.ClimateChange2013:ThePhysicalScienceBasis:WorkingGroupIContributiontotheIPCCFifthAssessmentReport(AR5),FinalDraftUnderlyingScientific-TechnicalAssessment. [2015-06- 28].http://www.climatechange2013.org/images/report/WG1AR5_SPM_FINAL.pdf
[2]唐秀萍. 碳匯 拓展林業外部空間. 中國林業, 2005, (22): 11- 17.
[3]SchlesingerWH,AndrewsJA.Soilrespirationandtheglobalcarboncycle.Biogeochemistry, 2000, 48(1): 7- 20.
[4]SehimelDS.Terrestrialecosystemsandthecarboncycle.GlobalChangeBiology, 1995, 1(1): 77- 91.
[5]ChenST,ZouJW,HuZH,ChenHS,LuYY.Globalannualsoilrespirationinrelationtoclimate,soilpropertiesandvegetationcharacteristics:Summaryofavailabledata.AgriculturalandForestMeteorology, 2014, 198- 199: 335- 346.
[6]魏書精, 羅碧珍, 孫龍, 魏書威, 劉芳芳, 胡海清. 森林生態系統土壤呼吸時空異質性及影響因子研究進展. 生態環境學報, 2013, 22(4): 689- 704.
[7]LuCQ,TianHQ,LiuML,RenW,XuXF,ChenGS,ZhangC.EffectofnitrogendepositiononChina′sterrestrialcarbonuptakeinthecontextofmultifactorenvironmentalchanges.EcologicalApplications, 2012, 22(1): 53- 75.
[8]吳迪, 張蕊, 高升華, 付曉, 鄧紅兵, 邵國凡, 張旭東. 模擬氮沉降對長江中下游灘地楊樹林土壤呼吸各組分的影響. 生態學報, 2015, 35(3): 717- 724.
[9]閆文德, 徐旺明, 鄭威, 王光軍, 張徐源, 梁小翠, 高超. 施氮對濕地松(Pinus elliottii)林土壤呼吸及其相關因子的消減效應. 生態學報, 2016, 36(2),doi:10.5846/stxb201311192767.
[10]孫素琪, 王玉杰, 王云琦, 張會蘭, 李云霞, 于雷, 胡波, 劉婕. 縉云山常綠闊葉林土壤呼吸對模擬氮沉降的響應. 林業科學, 2014, 50(1): 1- 8.
[11]SunZZ,LiuLL,MaYC,YinGD,ZhaoC,ZhangY,PiaoSL.Theeffectofnitrogenadditiononsoilrespirationfromanitrogen-limitedforestsoil.AgriculturalandForestMeteorology, 2014, 197: 103- 110.
[12]TuLH,HuTX,ZhangJ,LiXW,HuHL,LiuL,XiaoYL.Nitrogenadditionstimulatesdifferentcomponentsofsoilrespirationinasubtropicalbambooecosystem.SoilBiologyandBiochemistry, 2013, 58: 255- 264.
[13]HasselquistNJ,MetcalfeDB,H?gbergP.ContrastingeffectsoflowandhighnitrogenadditionsonsoilCO2fluxcomponentsandectomycorrhizalfungalsporocarpproductioninaborealforest.GlobalChangeBiology, 2012, 18(12): 3596- 3605.
[14]GundersenP,EmmettBA,Kj?naasOJ,KoopmansCJ,TietemaA.Impactofnitrogendepositiononnitrogencyclinginforests:asynthesisofNITREXdata.ForestEcologyandManagement, 1998, 101(1/3): 37- 55.
[15]FennME,PothMA,AberJD,BaronJS,BormannBT,JohnsonDW,LemlyAD,McNultySG,RyanDF,StottlemyerR.NitrogenexcessinNorthAmericanecosystems:predisposingfactors,ecosystemresponses,andmanagementstrategies.EcologicalApplications, 1998, 8(3): 706- 733.
[16]FangC,MoncriefJB.ThedependenceofsoilC02effluxontemperature.SoilBiologyandBiochemistry, 2001, 33(2): 155- 165.
[17]BowdenRD,NadelhofferKJ,BooneRD,MelilloJM,GarrisonJB.Contributionsofabovegroundlitter,belowgroundlitter,androotrespirationtototalsoilrespirationinatemperatemixedhardwoodforest.CanadianJournalofForestResearch, 1993, 23(7): 1402- 1407.
[18]ReyA,PegoraroE,TedeschiV,DeParriI,JarvisPG,ValentiniR.AnnualvariationinsoilrespirationanditscomponentsinacoppiceoakforestincentralItaly.GlobalChangeBiology, 2002, 8(9): 851- 866.
[19]SottaED,MeirP,MalhiY,NobreAD,HodnettM,GraceJ.SoilCO2effluxinatropicalforestinthecentralAmazon.GlobalChangeBiology, 2004, 10(5): 601- 617.
[20]姜艷, 王兵, 汪玉如, 楊清培. 亞熱帶林分土壤呼吸及其與土壤溫濕度關系的模型模擬. 應用生態學報, 2010, 21(7): 1641- 1648.
[21]WildungRE,GarlandTR,BuschbomRL.Theinterdependenteffectsofsoiltemperatureandwatercontentonsoilrespirationrateandplantrootdecompositioninaridgrasslandsoils.SoilBiologyandBiochemistry, 1975, 7(6): 373- 378.
[22]李化山, 汪金松, 劉星, 蔣思思, 張春雨, 趙秀海. 模擬氮沉降對太岳山油松林土壤呼吸的影響及其持續效應. 環境科學學報, 2014, 34(1): 238- 249.
[23]SongXZ,PengCH,ZhaoZY,ZhangZT,GuoBH,WangWF,JiangH,ZhuQ.QuantificationofsoilrespirationinforestecosystemsacrossChina.AtmosphericEnvironment, 2014, 94: 546- 551.
[24]ShabagaJA,BasilikoN,CaspersenJP,JonesTA.Seasonalcontrolsonpatternsofsoilrespirationandtemperaturesensitivityinanorthernmixeddeciduousforestfollowingpartial-harvesting.ForestEcologyandManagement, 2015, 348: 208- 219.
[25]李凱, 江洪, 由美娜, 曾波. 模擬氮沉降對石櫟和苦櫧幼苗土壤呼吸的影響. 生態學報, 2011, 31(1): 82- 89.
[26]張東秋, 石培禮, 張憲洲. 土壤呼吸主要影響因素的研究進展. 地球科學進展, 2005, 20(7): 778- 785.
[27]SimsPL,BradfordJA.Carbondioxidefluxesinasouthernplainsprairie.AgriculturalandForestMeteorology, 2001, 109(2): 117- 134.
[28]ArchMillerAA,SamuelsonLJ.Intra-annualvariationofsoilrespirationacrossfourheterogeneouslongleafpineforestsinthesoutheasternUnitedStates.ForestEcologyandManagement, 2015,doi:10.1016/j.foreco.2015.05.016.
[29]丁訪軍, 高艷平, 吳鵬, 崔迎春, 胡蕖. 喀斯特地區3種林型土壤呼吸及其影響因子. 水土保持學報, 2010, 24(3): 217- 221, 237- 237.
[30]劉春霞, 王玉杰, 王云琦, 馮華敏, 胡波. 重慶縉云山3種林型土壤呼吸及其影響因子. 土壤通報, 2013, 44(3): 587- 593.
[31]肖輝林. 大氣氮沉降對森林土壤酸化的影響. 林業科學, 2001, 37(4): 111- 116.
[32]FranklinO,H?gbergP,EkbladA, ?grenGI.PineforestfloorcarbonaccumulationinresponsetoNandPKadditions:bomb14Cmodellingandrespirationstudies.Ecosystems, 2003, 6(7): 644- 658.
[33]MaierCA,KressLW.SoilCO2evolutionandrootrespirationin11year-oldloblollypine(Pinus taedaL.)plantationsasaffectedbymoistureandnutrientavailability.CanadianJournalofForestResearch, 2000, 30(3): 347- 359.
[34]ZengXH,ZhangWJ,ShenHT,CaoJS,ZhaoX.SoilrespirationresponseindifferentvegetationtypesatMountTaihang,China.Catena, 2014, 116: 78- 85.
[35]ChenWJ,JiaX,ZhaTS,WuB,ZhangYQ,LiCY,WangXP,HeGM,YuHQ,ChenGP.SoilrespirationinamixedurbanforestinChinainrelationtosoiltemperatureandwatercontent.EuropeanJournalofSoilBiology, 2013, 54: 63- 68.
基金項目:國家自然科學基金項目(41401337)
收稿日期:2015- 06- 30; 2015- 06- 30;
修訂日期:2015- 11- 27 2015- 10- 29
*通訊作者
Corresponding author.E-mail: suyzh@lzb.ac.cn .E-mail: hebinghui@swu.edu.cn
DOI:10.5846/stxb201506301362 10.5846/stxb201506301367
Soil stoichiometry characteristics ofHaloxylonammodendronwith different plantation age in the desert-oasis ecotone, north China
ZHANG Ke1,2, SU Yongzhong1,*, WANG Ting1,2, LIU Tingna1
1KeyLaboratoryofEco-HydrologyinInlandRiverBasin,LinzeInlandRiverBasinResearchStation,ColdandAridRegionsEnvironmentalandEngineeringResearchInstitute,ChineseAcademyofSciences,Lanzhou730000,China2UniversityofChineseAcademyofSciences,Beijing100049,China
Abstract:Vegetation construction is one of the most important measures of ecological restoration in arid area and also one of the most effective methods of desertification control. Haloxylon ammodendron, one important sand-fixation plant in desert region of China with remarkable tolerate to drought, hot environmental stress, and can grow rapidly after planting on sand lands, was widely used for restoring vegetation on the peripheries of the oasis to fix the sand dunes in Linze Station since 1975. To clarify the effect of H. ammodendron on soil nutrient stoichiometric characteristics and explore the indicative significance of nutrient stoichiometric ratios on the recovery of soil, this study conducted the research on the artificial forest of H. ammodendron in an desert-oasis ecotone by analyzing soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), Available P and CaCO3 in an age sequence of 0-, 2- , 5- , 9- , 13- , 16- , 31- , and 39- year-old H. ammodendron plantations. Results showed that (1) distribution of SOC, TN and TP presented significant layered phenomenon along with the increase of plantation age, even though this feature in TP was relatively backward. In different soil depth, SOC and TN increased significantly with the increase of plantation age, but TP has no significant variation. (2) C∶P and N∶P also displayed obvious stratification trait and C∶P and N∶P in different layer also increased with the increase of plantation age. However, C∶N presented at a relatively constant variation. (3) The lower content of Available P expressed significant stratification feature after 2-year-old plantation and Available P and CaCO3 in 0—5 cm increased with the increase of plantation age. Nevertheless, CaCO3∶Available P also has no remarkable changes. (4) With the increase of plantation age, soil erodible fraction was highly significant decreased and has remarkably negative correlated with soil nutrients. These results exhibited significant increase of SOC and TN with the increase of establishment time. C∶N exhibited a stable trend, compared with C∶P and N∶P, indicating the formation of nitrogen need a relative fixation ratio of carbon and also stating C∶N that less susceptible to the effects of climate factors is quite stable. The small variation of TP mainly derived from the weathering of rock was strongly restricted by time. Moreover, the lower content of Available P and the narrow change of CaCO3∶ Available P manifested that the growth of H. ammodendron may limited by soil P and the limited Available P was more fixed by CaCO3. In addition, the negative relationship between soil nutrients and soil erodible fraction further showed that the establishment of H. ammodendron has improved soil texture and enhance soil nutrients contents. Our findings aimed at soil environment has enriched the study on H. ammodendron. However, further study is needed to clarify the response of soil nutrient stoichiometric characteristics to H. ammodendron and herbaceous, so as to provide more knowledge for the restoration effect of H. ammodendron on arid environmental and maintain sustainable development under the limited water condition.
Key Words:Haloxylon ammodendron; plantation age; soil stoichiometry characteristics; desert-oasis ecotone
Seasonal responses of soil respiration to simulated nitrogen deposition in a citrus plantation and masson pine forest in Mt. Jinyun, Chongqing, China
ZENG Qingping1, HE Binghui1,*, MAO Qiaozhi1, QIN Huajun2, LI Yuan3, HUANG Qi4
1KeyLaboratoryofThreeGorgesRegionEco-Environments,MinistryofEducation,CollegeofResourcesandEnvironment,SouthwestUniversity,Chongqing400715,China2GuizhouInstituteofBiology,Guiyang550009,China3ForestryAdministrationofFuling,Chongqing408000,China4HydroChinaGuiyangEngineeringCorporationLimited,Guiyang550009,China
張珂,蘇永中,王婷,劉婷娜.荒漠綠洲區不同種植年限人工梭梭林土壤化學計量特征.生態學報,2016,36(11):3235- 3243.
Zhang K, Su Y Z, Wang T, Liu T N.Soil stoichiometry characteristics ofHaloxylonammodendronwith different plantation age in the desert-oasis ecotone, north China.Acta Ecologica Sinica,2016,36(11):3235- 3243.
