兩系不育系Y58S的花藥培養與花培后代的鑒定篩選
2016-08-06 03:58:20黃翠紅楊瑰麗周丹華劉永柱陳志強
廣東農業科學 2016年5期
黃翠紅,楊瑰麗,黃 明,周丹華,劉永柱,郭 濤,陳志強,王 慧
(華南農業大學國家植物航天育種工程技術研究中心,廣東 廣州 510642)
兩系不育系Y58S的花藥培養與花培后代的鑒定篩選
黃翠紅,楊瑰麗,黃 明,周丹華,劉永柱,郭 濤,陳志強,王 慧
(華南農業大學國家植物航天育種工程技術研究中心,廣東 廣州 510642)
花藥培養技術與傳統不育系育種的結合能有效加快新的光溫敏不育系材料的選育進程。為了篩選出適合兩系不育系Y58S的花藥培養條件,利用10種不同培養基對兩系不育系Y58S不同雜交組合F2代進行花藥培養。培養基E(NMB+2 mg/L 2,4-D+2 mg/L NAA+1 mg/L KT+0.5 g/L脯氨酸+0.5 g/L谷氨酰胺+0.5 g/L水解酪蛋白+25 g/L蔗糖+25 g/L麥芽糖+10 g/L瓊脂粉)較為適合兩系不育系Y58S的花藥培養;同時,材料的基因類型對花藥培養力有著顯著的影響,粳稻血緣的滲入有利于兩系不育系花藥培養力的提高。花藥培養后代田間性狀觀察結果表明,在當代花藥培養植株中發現較多的二倍體植株,少部分為單倍體、四倍體植株,尚未發現非整倍體;篩選出一個表現較好的不育株系。
水稻;兩系不育系;Y58S;花藥培養;花培后代
黃翠紅,楊瑰麗,黃明,等.兩系不育系Y58S的花藥培養與花培后代的鑒定篩選[J].廣東農業科學,2016,43(5):31-36.
不斷提純改良不育系是兩系法雜交水稻研究和超級雜交水稻組合選育的關鍵。鑒于花培育種具有純合快、選育效率高、可使農藝性狀和育性同步穩定等優點,花藥培養技術可在較短的周期內使包括溫光反應特性在內的各種性狀不再分離,大大縮短育種進程,也可用于育成不育系的提純[1]。周元昌等對培矮 64 S 的核心種子進行花藥培養,結果在H后代株系間發生育性分離,篩選出了完全不育的不育系培矮 64 HS,在不育期間其花粉不育度達99.8%以上,且不育期較長,育性和后代遺傳均穩定。證明了花藥培養在培矮64S的遺傳純化上是有效的[2]。馬鎮榮等也用花藥培養對兩用核不育系培矮64S進行提純與改良,結果表明花藥培養是提純及改良兩用核不育系的有效途徑[3]。陳遠孟等對3個光(溫)敏核不育系培選S、農墾58S、以及培矮64S進行花藥培養,其花粉植株均有育性轉換的特性,且在H3代花粉植株的株系中,得到花粉不育度>99.5%的植株比對照的株系多,表明了應用花培對光(溫)敏核不育系提純的可行性[4]。但在水稻花藥培養中,基因型是影響花藥培養里的一個重要因素[5],一般來說,花藥培養力的大小為:糯稻>粳稻>粳/秈稻>秈型雜交稻>秈稻[6]。而光溫敏核不育系和非光溫敏核不育系在花藥培養效率上不存在顯著差異,花藥培養里僅與供試材料的基因型有關[7]。利用光溫敏核不育材料進行花藥培養雖然可以加速不育系的選育,但可能由于培養條件不合適,或者花粉的敗育而影響愈傷的誘導和分化。因此,需要尋找新的培養基或調整現有配方,還包括附加成分的增減、培養條件的改善等,目的在于提高愈傷組織的誘導率和綠苗的分化率[8]。
Y58S 是湖南雜交水稻研究中心新選育而成的光溫敏不育系,2005年2月通過湖南省農作物品種審定。它具有配合力高、抗逆性強、綜合性狀優良等特點,適合配制廣適性超級稻組合。我們通過對光溫敏核不育系Y58S不同雜交后代進行花藥培養研究,探究出對光溫敏核不育系材料適宜的花培條件,同時通過對花培后代材料篩選,以期獲得新的兩系不育系材料,為花培育種提供理論基礎。
1 材料與方法
1.1 試驗材料
兩系不育系材料:Y58S/02428、Y58S/合豐占,F2代(F1代中選擇不育株系割兜留禾頭,經人工氣候箱處理轉育后留種)。
1.2 試驗方法
1.2.1 取材與低溫預處理 試驗在華南農業大學教學試驗場農學分場進行,供試材料于2013年早造開始種植,常規田間管理。當植株處于孕穗期時,在晴天下午從田間選取劍葉與下一葉的葉枕距約5~10cm的帶苞葉稻穗。將多數花粉發育時期處于單核中晚期的稻穗用濕潤紗布包裹,置于7~8℃下低溫預處理7~10 d。
1.2.2 培養基 誘導、繼代培養基:分別選用N6 和NMB為基本培養基,并在激素水平與有機添加物上進行調控(表1)、瓊脂10 g/L、蔗糖和麥芽糖各25g/L。
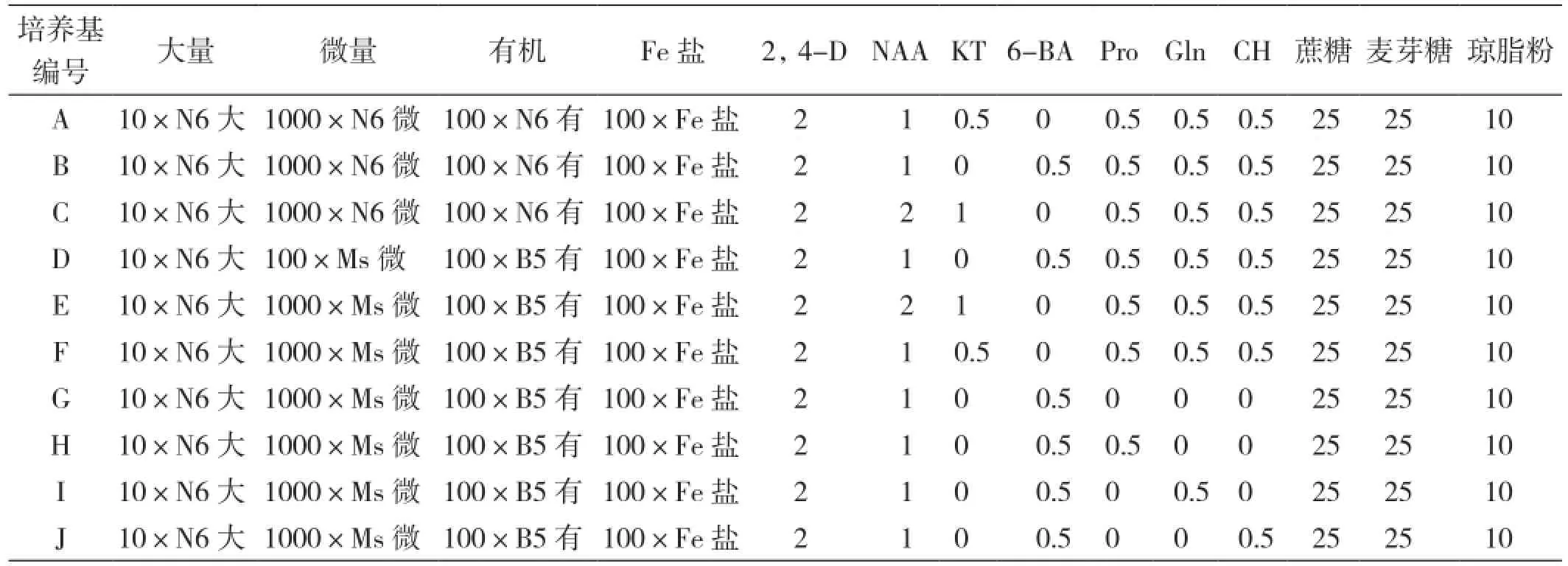
表1 不同培養基水平的篩選
分化培養基:統一使用MS為基本培養基,附加6-BA 3 mg/L、NAA 1 mg/L、蔗糖30 g/L、瓊脂10 g/L。
生根壯苗培養基:采用l/2MS培養基,不附加激素,蔗糖25 g/L。
所有培養基pH值均調至5.8左右。
1.2.3 接種條件 用75%酒精對稻穗進行表面消毒,剝除葉鞘,置于燒杯中用2%次氯酸鈉消毒10~15 min,期間不時震蕩攪拌,以保證消毒完全,然后用無菌水沖洗3遍,用無菌紗布吸干水分,在無菌條件下用“剪穎抖藥法”把花藥接種至培養瓶中。將供試材料接種在不同誘導培養基中,每種培養基上都分別接種10瓶,每瓶約100~120枚花藥,每個處理3次重復。
當愈傷長出至2~3 mm時,挑選乳白色、表面光滑、結構致密的愈傷組織轉入分化培養基中培養。每個供試材料分別接種8瓶,每瓶接種8塊愈傷組織塊。
1.2.4 培養方法 花藥培養分為誘導培養階段和分化培養階段,誘導階段于26~28℃下暗培養,于接種后2~3 d統計各培養瓶中所接種的花藥枚數,接種后30~60 d統計愈傷組織塊數;分化階段先暗培養3 d,后轉至光照培養,使其有一個過渡適應階段,更快適應轉換了的環境,進而提高分化力。一般光培養的光照時間為10~12 h/d,光強1 000~1 500 lx,室溫保持26~28℃,分化培養45 d進行統計綠苗叢數、白苗叢數等。

整理數據并進行方差分析,確定各材料在不同培養基中對愈傷組織誘導率和后續分化效果的影響。
利用SAS數據處理系統軟件進行數據統計分析。一般的數據處理用Excel,統計分析用SAS 9.1.3軟件,差異顯著性用鄧肯氏新復極差法測驗。
2 結果與分析
2.1 Y58S不同雜交組合F2代在不同培養基中的花藥培養結果
2.1.1 Y58S不同雜交F2代的愈傷誘導情況 對不同培養基中的水稻材料的出愈率(愈傷組織誘導率)進行統計及分析結果(表2)表明,Y58S/02428的雜交F2代和Y58S/合豐占F2代在不同的誘導培養基上其誘導率有所不同。 可能由于02428是粳稻材料,而合豐占是秈稻材料,所以含有粳稻血緣的Y58S/02428組合的誘導率會高一些。其中,Y58S/02428組合在G培養基上的誘導率最高,為14.33%,而Y58S/合豐占組合反而最低,只有1.57%。而在同一個雜交組合內,誘導率也存在差異,Y58S/02428組合在G培養基上誘導率可達到14.33%,其次分別為12.55%(D培養基)、11.71%(I培養基)、11.12%(E培養基),而在B培養基上才4.27%;Y58S/合豐占組合在E培養基上誘導率表現最高,為8.38%,培養基D (7.18%)、B(6.30%)次之,而在G培養基上的誘導率表現最低,僅為1.57%,說明不同的培養基對花藥培養的誘導率有一定影響,而不同血緣的材料對培養基的要求也有所不同。

表2 Y58S不同雜交組合在不同培養基的花藥誘導情況
對兩種不同的雜交組合在10種培養基上的誘導率進行雙因素有重復的方差分析,結果(表3)表明,材料間與培養基間的差異均達到極顯著水平,這證明了不同雜交組合的花藥來源以及培養基的類型對愈傷組織誘導率的影響顯著。進一步對誘導率進行多重比較(統一使用Duncan新復極差測驗)的結果表明,在不同雜交組合方面,Y58S/02428組合的平均誘導率為9.37%,顯著高于Y58S/合豐占組合(4.54%);而在培養基方面(表4),使用C培養基進行組織培養,所得到的誘導率是最低的,僅為4.40%;而D培養基的誘導率最高,為9.95%,E培養基(9.82%)次之,但兩者差異不顯著,因而可以將這兩種培養基視為較為適合于花藥培養的誘導培養基。
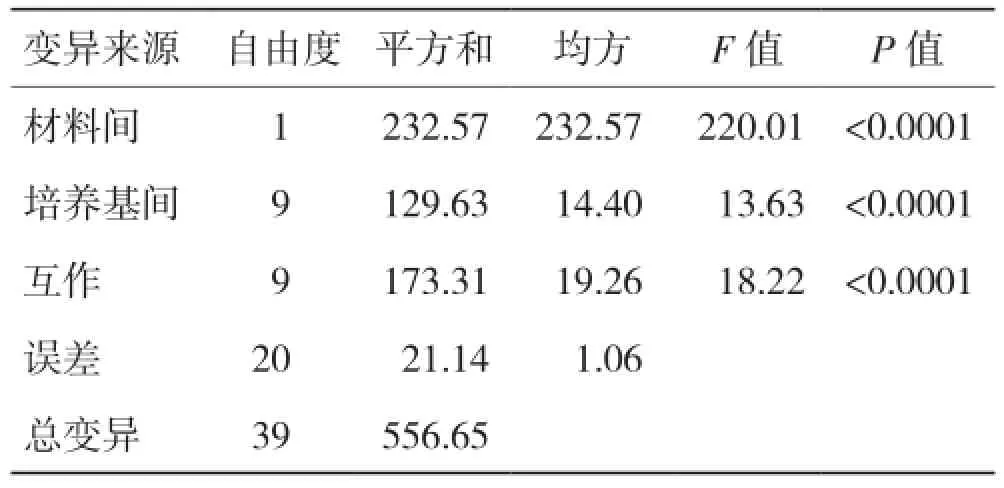
表3 Y58S不同雜交組合在不同培養基花藥誘導率的方差分析

表4 不同培養基對花藥誘導率的多重比較分析
2.1.2 Y58S不同雜交F2代的愈傷分化情況 將10個不同雜交組合F2花藥誘導出來的愈傷組織轉移到相同的分化培養基上進行后續分化培養,并對其愈傷組織分化結果進行統計,結果(表5)顯示,不同誘導培養基誘導出來的愈傷組織因質量水平不同,分化情況各異,部分出現零分化。綠苗分化率方面,最高值表現在F培養基上(75.00%),但只Y58S/合豐占組合有分化,而 Y58S/02428雜交組合卻是零分化;對于Y58S/02428雜交組合,最高綠苗分化率出現在培養基E上(25.00%),對應Y58S/合豐占的分化率為15.38%(圖1)。總的來說,E培養基對于不同材料的綠苗分化率較為有利,廣譜性較強。
在花藥培養力(綠苗產率)方面,兩個雜交組合在誘導培養基E和誘導培養基B上表現稍好,且培養基E(2.78%、1.29%)更優于培養基B (0.36%、0.35%)。誘導培養基E較為適合Y58S不同血緣材料的花藥培養,且具有一定的廣適性。Y58S/02428組合的花藥培養力均高于Y58S/合豐占雜交組合,表明轉入粳稻血緣有利于花培效力的提高,這與前人研究結果相同[6]。

表5 Y58S不同雜交組合在不同培養基的愈傷分化率
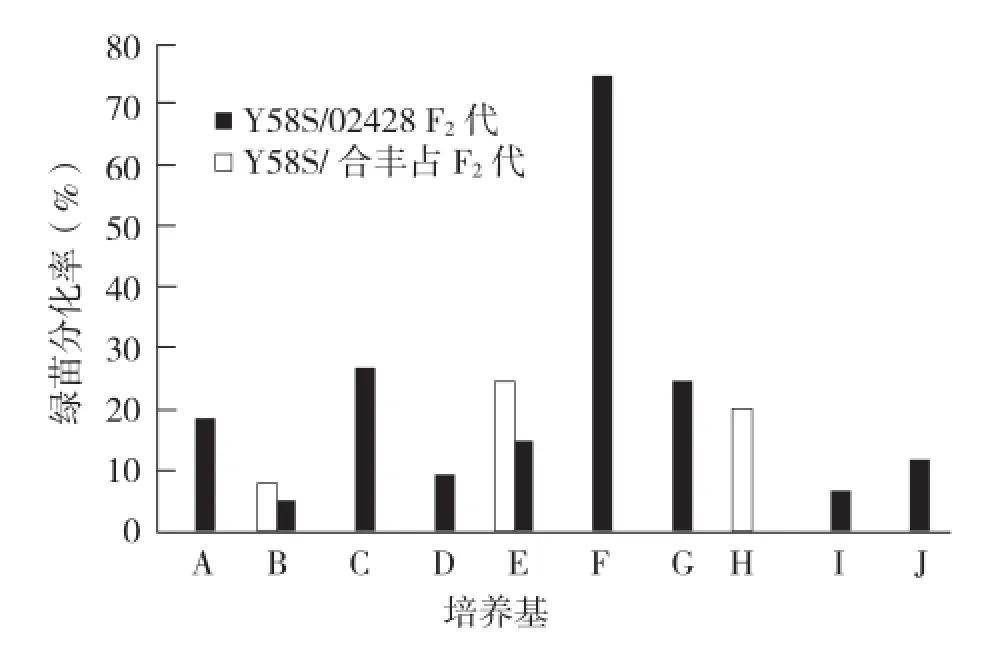
圖1 Y58S不同雜交組合在不同培養基的綠苗分化率
白苗分化率方面,除了出現零分化的培養基,A培養基對于兩個雜交組合的白苗分化率都比較低,在Y58S/02428為1.04%、Y58S/合豐占為4.76%,其次是E培養基,Y58S/02428與Y58S/合豐占雜交組合的白苗分化率分別為8.33%、7.69%。表明誘導培養基A與誘導培養基E在控制白苗分化的水平上表現較好。
2.2 Y58S不同雜交組合F2花藥培養后代的田間表現
2.2.1 花藥培養后代的倍性觀察 經過對花藥培養后代植株利用流式細胞儀進行初步倍性鑒定并結合田間種植表現觀察,結果(表6)表明:在當代花藥培養植株中發現較多的二倍體植株(48.00%),少部分為單倍體(23.05%)、四倍體植株(7.19%),尚未發現非整倍體。其中,單倍體表現為植株矮小,葉片較小,根細小;四倍體表現為植株較高、粗壯,葉片較寬,根較粗壯;而二倍體則處于這兩者間,并與原生種相似。

表6 花藥培養植株當代的倍性表現
2.2.2 花藥培養后代的育性觀察 Y58S/02428與Y58S/合豐占兩個 F2代組合獲得的花培后代的田間表現以偏粳型較多,結合花藥培養后代的育性觀察與鏡檢鑒定,同時從花培后代篩選出花粉不育且柱葉型表現較好株系6株,割兜留禾頭,待其孕穗、育性轉育敏感期移至人工氣候箱處理,成功轉育并收獲株系2株。2015早造繼續種植,其中一個來自Y58S/02428 F2代組合的花培后代株系的株葉型表現較好,育性觀察與鏡檢結果(圖2、圖3,封二)顯示為典敗類型,其育性穩定性有待于進一步的研究觀察。
3 結論與討論
一般情況下,秈稻品種其花藥培養力較低,粳稻培養效率較高,秈粳雜交后代居中[9]。本研究以光溫敏核不育系Y58S為材料,對其不同雜交組合F2代進行花藥培養也發現,不同基因型材料對花藥培養力影響極為顯著。不同雜交組合其花藥培養力也有所不同,與其雜交導入的外源基因有一定的關系,對于光溫敏核不育系Y58S來說,雜交組合Y58S/02428培養效果優于雜交組合Y58S/合豐占,說明導入粳稻血緣能提高不育系的花藥培養力,并與前人研究結果相同,但導入粳稻血緣含量高低對秈稻花藥培養力影響大小還有待于進一步研究。另外,培養基類型是影響花藥培養力的重要影響因素。對于不同的基因型材料,需要的培養基成分組成不同。一般情況下,誘導培養基中,N6適于粳稻、SK3適于秈粳雜交、合5適于秈稻、通用和M8對秈稻和粳稻均較適用。馮雙華等選用合5、M8和改良M8為基本培養基對培兩優288、P88S/0293、秈型雜交稻兩優培九進行花培,結果表明,改良M8培養基比較適合不同的基因型材料[10]。王伍梅等通過多年的試驗結果表明,N6培養基對多數秈粳交后代和粳稻的誘導效果都較好[11]。黃慧君等對不同激素種類以及濃度配比進行多次實驗研究表明,秈稻品種要求更高的生長素濃度,并且配合一定濃度的KT,有利于愈傷組織誘導率和綠苗的提高[12]。也有人認為,適當的降低生長素的濃度,對愈傷組織的誘導率有促進作用。本研究通過改變培養基配方(包括基本培養基、激素配比以及有機添加物進行綜合調配),探索到培養基E是適合光溫敏不育系的培養基,其中誘導培養基選用NMB培養基和激素配比2,4-D∶NAA∶KT為2∶2∶1較為適合,但秈稻花藥培養力與粳稻相比仍有一段距離,還需要進一步探究最佳的培養基成分。
愈傷組織在誘導階段往往會發生變異,出現染色體成倍增加、減少或缺失等,從而使花藥培養后代出現多倍體、單倍體或非整倍體等,因此對花藥培養后代進行倍性鑒定尤為重要。
水稻花藥組織培養后產生的植株除單倍體外,還會有二倍體、三倍體以及各種異倍體[13],花藥植株倍性發生變異主要是由于離體培養細胞染色體的數目和結構不穩定,在花藥培養中發生異常有絲分裂。如要獲得單倍體或純合二倍體,不但要做好倍性鑒定工作,前期培養條件也很重要。花藥培養易受到體細胞的干擾,在產生單倍體的同時也產生二倍體或四倍體;同時單倍體細胞的生長很容易被生長旺盛的多倍體細胞所掩蓋[14]。本試驗采用流式細胞分析法鑒定花藥組織培養后代的倍性為輔助手段,并結合田間觀察方法,抽樣花藥培養苗為185株,其中二倍體的平均頻率最高;其次為單倍體;而多倍體出現的頻率,未發現非整倍體、嵌合體等現象。表明光溫敏不育系Y58S/02428與Y58S/合豐占兩個 F2代與其他水稻材料一樣,經過花藥組織培養后會發生自然加倍,并且受到多重因素的影響,其中兩個組合獲得的花培后代的田間表現以偏粳型居多,通過育性與田間表現篩選,其中一株來自Y58S/02428組合的不育單株表現較好將留作進一步深入的研究,期望獲得具有一定育種價值的育性穩定純合的新兩系不育系材料。本研究結果進一步表明利用光溫敏核不育材料進行花藥培養可以加速兩系不育系的選育,同時開展的花藥培養條件的優化研究,提高并改善花藥培養力,能有效服務于光溫敏核不育系水稻的提純以及水稻育種工作。
[1]梁曼玲,陳彩虹.低溫敏核不育系培矮64S的花藥培養[J].廣西農業科學,2002(4):171-173.
[2]周元昌,林荔輝,江樹業,等.花藥培養遺傳純化溫敏核不育系培矮64S的效果初步分析[J].中國水稻科學,2000(2):56-58.
[3]馬鎮榮,凌定厚,王昌虎.花藥培養對秈稻兩用核不育系培矮64S的提純與改良[J].熱帶亞熱帶植物學報,2000(4):308-314.
[4]陳遠孟,呂志仁,韋鵬霄,等.花藥培養對光(溫)敏核不育系親和力的影響[J].廣西農業科學,2002 (3):112-113.
[5]孫宗修,程式華,閔紹楷,等.光敏核不育水稻的光溫反應研究Ⅲ.減數分裂期溫度對兩個秈稻光敏不育系育性轉換的影響[J].作物學報,1993(1):83-87.
[6]沈錦驊,李梅芳,陳銀全等.花藥培養在水稻品種改良上的應用[J].中國農業科學,1982,4(2):15-19.
[7]蔡得田,陳冬玲,祝虹等.湖北光溫敏核小育水稻的未受精子房和花藥培養出單倍體[J].實驗生物學報,1988,2(4):401-407.
[8]李艷萍.水稻花藥培養與花培育種研究[J].天津農業科學,2003,9(4):36-38.
[9]Dewi I S,Purwoko B S,Aswidinnoor H,et al.Plant regeneration from anther cultures of several genotypes of indica rice tolerant to aluminum toxicity[J].Indones J Agric Sci,2009,2:1-5.
[10]馮雙華,趙森,郭家源,等.不同培養基和激素對超級雜交稻花藥培養力的影響[J].西南農業大學學報(自然科學版),2006(4):523-525.
[11]王伍梅,臺德衛,張效忠,等.水稻高效花藥培養技術體系的構建[J].中國農學通報,2009(16):65-68.
[12]黃慧君.外源激素、培養基對水稻幼穗培養直接分化和脫分化的作用[J].廣東農業科學,1993(3):5-7.
[13]Nishi T S,SuokaMit.Occurrence of various ploidy plants from anther culture and ovary culture of rice plant[J].Japan J Genetici,1969,44:341-246.
[14]周旭紅,莫錫君.花藥培養的研究進展[N].江西農業學報,2007,19(8):74-76.
(責任編輯 楊賢智)
Primary research on anther culture of photothermosensitive genic male sterile rice “Y58S”and selection of descendant of anther culture
HUANG Cui-hong,YANG Gui-li,HUANG Ming,ZHOU Dan-hua,LIU Yong-zhu,GUO Tao,CHEN Zhi-qiang,WANG Hui
(National Engineering Research Center of Plant Space Breeding,South China Agricultural University,Guangzhou 510642,China)
The combination of anther culture techniques and traditional sterile lines breeding can effectively speed up the improvement of photo-thermosensitive genic male sterile(PTGMS) lines.To screen for the optimal culture medium for the two-line male sterile line,Y58S,we studied the effects of 10 different culture mediums on the F2generation of different cross combination between Y58S and selected male parents.The results showed that medium E(NMB+2 mg/L 2,4-D +2 mg/L NAA+1 mg/L KT+0.5 g/L Pro+0.5 g/L Gln+0.5 g/L CH+25 g/L sugar+25 g/L Maltose+10 g/L Agar)was more effective for anther culture of PTGMS lines.And introducing japonica rice by cross breeding improved anther culture ability of PTGMS lines,indicating that genetype played an important role in rice anther culture.The examination on traits in field of descendants displayed that most of the plants were diploid,some were haploid and tetraploid,and no non-integer ploidy,and a new PTGMS lines was screened.
rice;PTGMS lines;Y58S;anther culture;descendant of anther culture
S511.0353
A
1004-874X(2016)05-0031-06
10.16768/j.issn.1004-874X.2016.05.007
2016-01-28
