珍稀克隆植物北極花有性繁殖及其對資源的分配和利用
2016-07-23 01:51:10翟雅芯王玥魏仙仙何爽張愛勤
新疆農業科學 2016年6期
關鍵詞:植物
翟雅芯,王玥,魏仙仙,何爽,張愛勤
(新疆大學生命科學與技術學院,烏魯木齊 830046)
?
珍稀克隆植物北極花有性繁殖及其對資源的分配和利用
翟雅芯,王玥,魏仙仙,何爽,張愛勤
(新疆大學生命科學與技術學院,烏魯木齊830046)
摘要:【目的】有性繁殖使克隆植物具有潛在適應環境變化的能力,對植物生活史具有重要影響。【方法】為了深入理解植物有性繁殖各構成因素之間的相互關系及對資源的分配和利用情況,通過野外觀測及室內測定,研究珍稀克隆植物北極花有性繁殖與相關構成因素。【結果】北極花分株長達(60.14 ± 5.70)cm,各分株的構件參數營養枝數量、有性枝數量和分株果實數分別為(8.23 ± 1.18)枝、(2.50 ± 0.30)枝和(1.47 ± 0.29)個。有性繁殖所占分株生物量比例為7.06% ± 1.30%。在分株水平上,有性繁殖適合度的直接影響因素為有性分枝數量,不受分株長度和生物量的影響;而營養枝作為克隆株的同化器官,可間接的影響有性繁殖。在居群水平上,斑塊內果實數與有性分枝數量、結實率的相關系數分別為0.849、0.847,均呈極顯著相關(P<0.01);而與斑塊大小(面積)無相關性。【結論】北極花具有相對低的有性繁殖狀態,有性繁殖的衡量參數果實數僅與有性分枝數量和結實率相關,而表現出與對繁殖適合度有較大貢獻的其他性狀不相關。依據遺傳不育進化的流行假說,如果北極花的有性繁殖對維持居群遺傳多樣性的貢獻率過低,那么其有性繁殖有通過不育突變而衰退的風險。
關鍵詞:北極花(Linnaea borealis L.);克隆植物;有性繁殖;構件參數;相關分析
0引 言
【研究意義】大多數克隆植物兼具有性和無性兩種繁殖方式,并對居群的遺傳結構產生重要影響。有性繁殖可通過配子體的有性融合、染色體分離和等位基因重組等產生遺傳變異[1],以增加植物潛在的適應環境變化的能力。但不同克隆植物之間有性繁殖和實生苗的更新能力差異很大,一些植物因克隆繁殖甚至放棄了有性過程,進而導致物種遺傳多樣性的降低和對環境適應力的下降[2-3]。隨著珍稀物種孤立小種群因有性繁殖衰退而導致瀕危的報道不斷增多[4-6],植物有性繁殖及其相關因素的研究成為許多克隆植物生活史研究的重要內容。植物的有性繁殖受多種因素的影響:如生境破碎化使物種以小居群的形式存在,居群間基因流中斷,近交比例過高,兩種繁殖方式間的資源權衡,及由此產生花粉輸出下降的雄性適合度代價和干擾結實產生的雌性適合度代價。尤其像高山、極地等一些特殊環境,傳粉者數量有限、結實率過低、或者結實后種子難以萌發等問題也成為植物有性繁殖的重要制約因素[4,7-10]。【前人研究進展】北極花(LinnaeaborealisL.)也叫林奈木,以分布在北半球高緯度地區而命名,也因優美的花型和作為植物界最小的灌木,被冠以著名分類學家的名字林奈,而深受喜愛。作為高緯度寒冷地區的一個常綠、匍匐小灌木,具克隆生長習性的一個珍稀物種,由于居群的片斷化導致有性繁殖受阻[12-15]。在蘇格蘭等地已被定為瀕危物種[16],我國目前將其作為珍稀物種記載[17]。目前針對克隆植物有性繁殖的研究主要集中在植物兩種繁殖方式的進化權衡、克隆生長在花粉傳遞與交配系統方面對有性繁殖所產生的影響[11]。【本研究切入點】自然狀態下,有性繁殖與其構成因素的相互作用及各因素對資源的利用和分配情況還缺乏研究。嚴酷的生境集有性、無性繁殖于一體,使北極花成為進行植物繁育系統多樣性研究和評估瀕危克隆植物有性繁殖狀態,分析瀕危機制的較好材料。【擬解決的關鍵問題】研究北極花的有性繁殖及相關構成因素如分株果實數、分株長度、有性分枝數、無性分枝數、分株生物量大小、結實率等,為揭示克隆植物的有性繁殖狀態和多樣的繁育系統類型積累新的數據。
1材料與方法
1.1材 料
觀測點位于86°55′E~87°03′E,48°36′N~48°42′N的喀納斯自然保護區海拔1 500~2 000 m的針葉林下。北極花為游擊型克隆構型,以每年產生6~8對對生葉片、長度30~50 cm的方式向前延伸,具較強的克隆生長力。有性和無性兩類垂直短枝著生在對生葉片的葉腋處。花序生于有性枝的頂端,具2花,雙懸垂,花果期6~8月。圖1
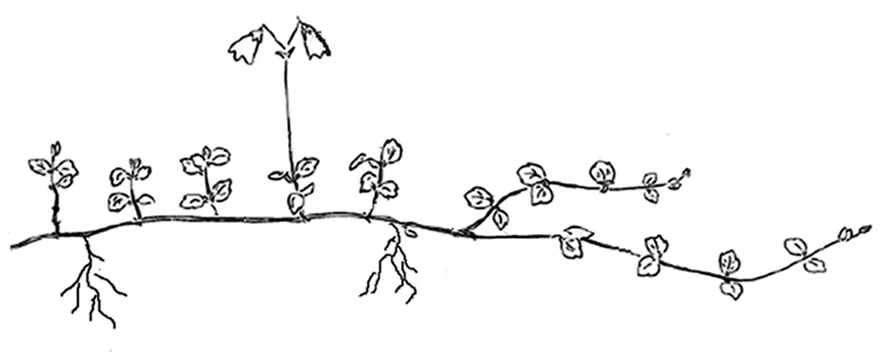
圖1 北極花游擊型克隆分株
1.2方 法
1.2.1分株水平上有性繁殖與構成因素的測定及各構成因素的相關分析
在北極花結實末期,以間隔5 m左右的距離為準,在30個不同的克隆斑塊內隨機取匍匐莖分株46個,對所有分株的長度分別進行測量。同時,隨機對30個分株進行有性枝、無性枝數量及分株果實數的統計。并對分株果實數與各構成因素進行相關分析。
1.2.2分株生物量及有性繁殖分配比例的測定及相關分析
在北極花結實末期,間隔一定距離在不同的克隆斑塊內隨機選取30個匍匐莖分株,清理干凈泥土后,將各分株分為有性繁殖構件(所有有性分枝及其上邊的果實)、無性分枝、及分株匍匐莖三部分,在80℃恒溫箱中烘72 h、分別稱干重,后進行各分株生物量和有性繁殖分配比例的統計。
1.2.3居群水平上有性繁殖格局及其與構成因素的相關分析
在結實期隨機選定不同面積大小的樣方(克隆斑塊)30個,對其面積進行測定;統計單位面積內有性分枝數量和果實數,計算結實率;并進行斑塊內果實數與面積大小等各構成因素的相關性分析。
1.2.4種子與幼苗的補充及更新
為了從后代適合度方面衡量有性繁殖的貢獻率,隨機選定不同面積大小的克隆斑塊10個,進行如下觀測:(1)對斑塊內實生苗數量進行統計;(2)對居群中當年新形成的果實及種子進行發育狀況的跟蹤觀察。(3)對斑塊內不同時期的結實率進行統計。結實率為克隆斑塊內所有果實數與單花數的比。
1.3數據處理
用SPSS 17.0,Excel 2007軟件進行數據處理、分析和繪制圖表。首先用One-Sample Kolmogorov-Smirnov Test進行正態性檢驗,經檢驗各組數據均符合正態分布后,再進行相關數據分析。所有統計數據均用平均值±標準誤表示。
2結果與分析
2.1分株水平上有性繁殖與構成因素
研究表明,北極花克隆分株的長度變化幅度較大,最長的達145.60 cm,最短的僅8.55 cm;平均長度為(60.14 ± 5.70 )cm,為典型的游擊型克隆構型。分株上有性和無性枝條從葉腋處伸出,隨機分布在匍匐莖上,平均每分株有營養枝(8.23 ± 1.18)枝(最大值為22枝,最小值為0);有性枝(2.50 ± 0.30)枝(最大值為8枝,最小值為0)。每分株上有效果實數最大值為6個,最小值為0,平均為(1.47 ± 0.29)個。
從各相關參數的測定值看,北極花分株較長,但各分株的構件參數如營養枝數量、有性枝數量、分株上果實數等較低,且各參數變異幅度較大。尤其是有性繁殖的構成因素如有性枝數量、果實數及生物量分配比例等顯得尤為低下。表1
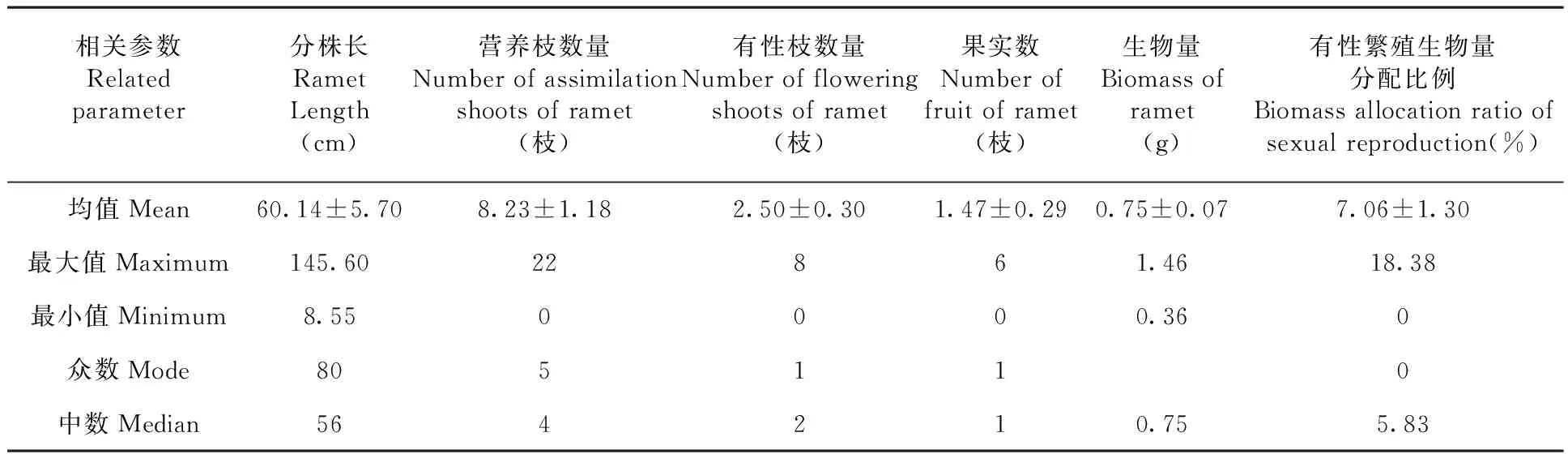
表1 分株水平上北極花果實數量與構成因素的測定值(平均值±標準誤)
2.2分株生物量及有性繁殖分配比例
研究表明,北極花分株生物量為(0.75 ± 0.07)g,分株上營養枝和分株匍匐莖所占的生物量比例分別為45.32% ± 3.56%,47.62% ± 3.33%;遠高于有性繁殖7.06% ± 1.30%的生物量分配比例。
北極花被稱為世界上最小的小灌木,從分株生物量的大小就可看出其細弱的株型。但較長的分株匍匐莖,說明了匍匐莖及其附著的根系在克隆株營養物質儲藏、資源的獲取、基株生長壽命的維持方面所具的重要功能。在分株各構件參數中,分株匍匐莖生物量最高也說明了這一現象。圖2
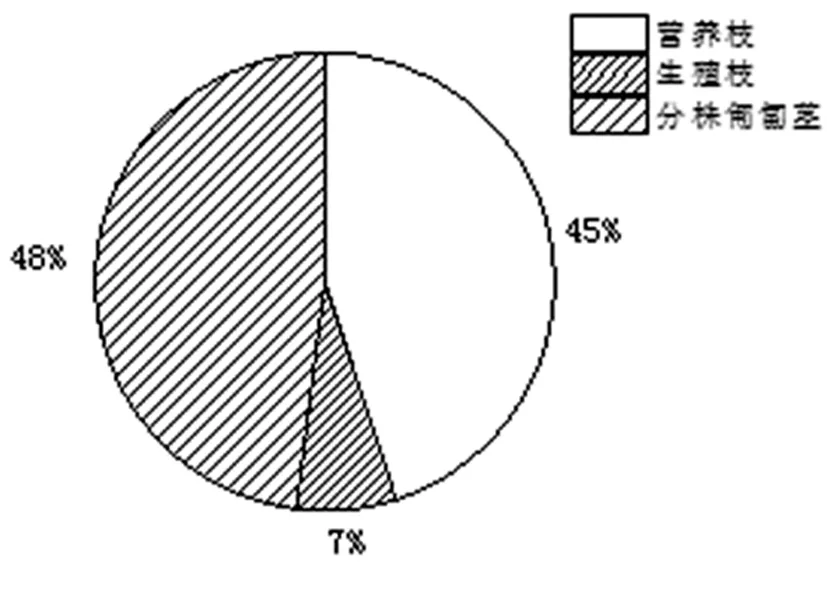
圖2 北極花分株各構件生物量分配比例
2.3分株水平上北極花有性繁殖與構成因素的相關性
研究表明,不同分株長度的果實數與有性枝數量的相關系數是0.389,呈顯著相關(P<0.05);與分株長度、分株上無性枝數量、分株的生物量無顯著相關性。而在各構成因素中,有性枝數量與營養枝數量的相關系數是0.422,為顯著相關(P<0.05);分株長度與營養枝數量的相關系數為0.821,呈極顯著正相關(P<0.01);與有性枝數量、生物量和果實數相關系數分別為0.291、0.032和0.017,相關性均不顯著。
從各構成因素的相關性看,有性繁殖適合度的直接影響因素為有性分枝數量,而營養枝為克隆株的同化器官,是分株光合產物的主要提供者,與有性枝的數量呈正相關,可間接的影響有性繁殖。表2
2.4克隆斑塊內有性繁殖格局及果實數與構成因素的相關性
研究表明,北極花斑塊內的果實數與有性分枝數量、結實率均呈極顯著正相關(P<0.01),其相關系數分別為0.849、0.847;而與斑塊面積的相關性不顯著,表明在斑塊內,北極花的果實數、有性分枝數呈明顯的非均勻分布,有性繁殖適合度主要受斑塊內有性枝數量和結實率影響。表3
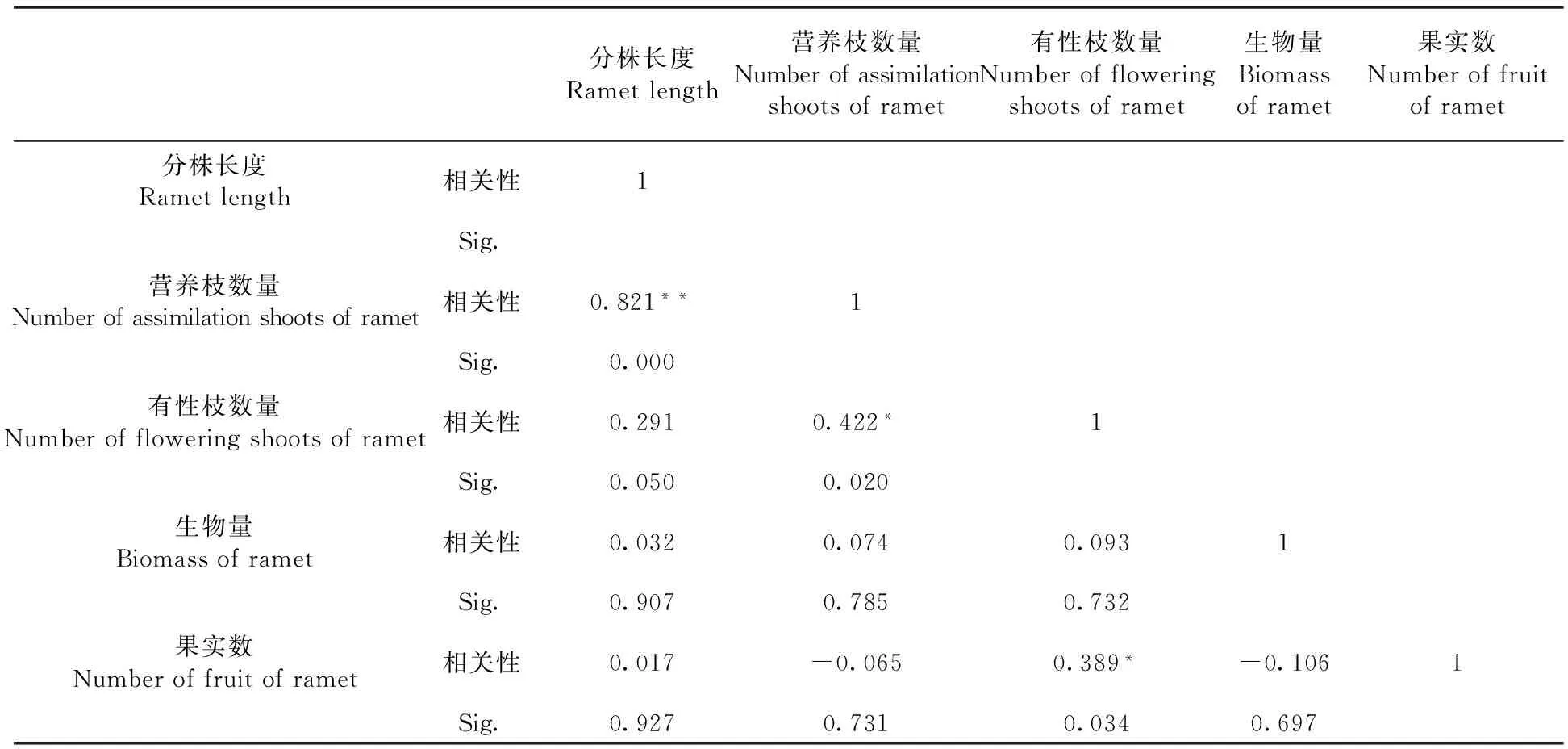
表2 分株水平上北極花果實數與構成因素的相關性
注:**在0.01水平(雙側)上顯著相關。*在0.05水平(雙側)上顯著相關
Note:**the 0.01 level. (bilateral). A significant correlation at 0.05 level (bilateral) significant correlation
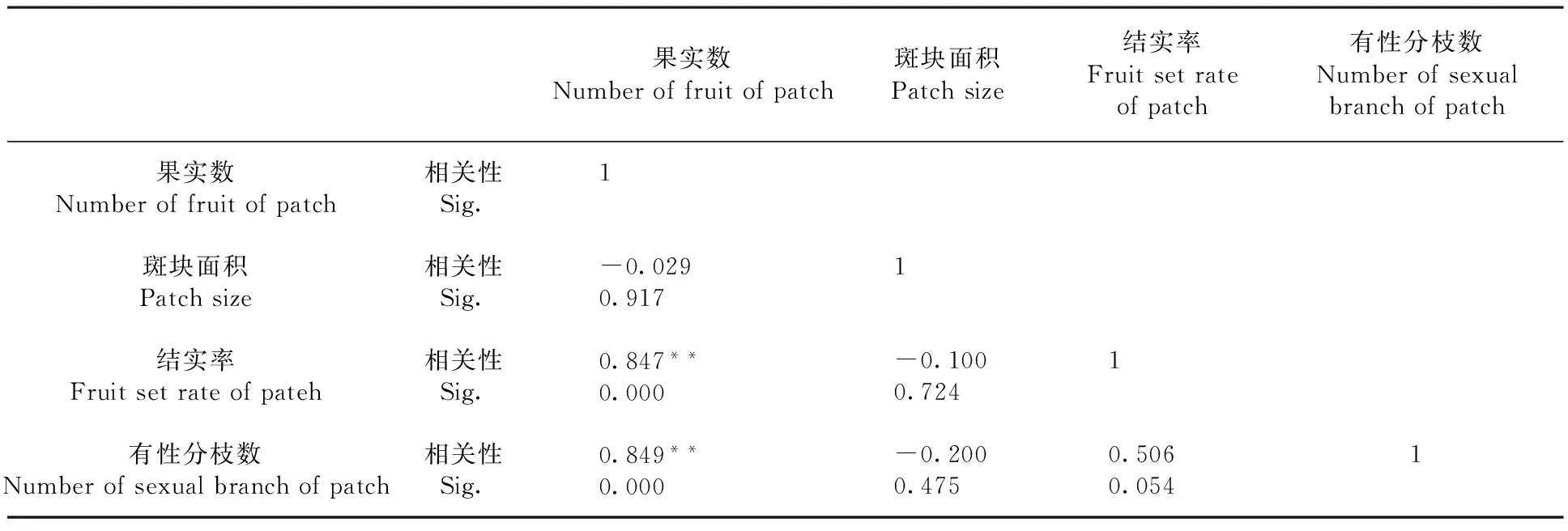
表3 北極花克隆斑塊內果實數與斑塊面積大小、結實率和有性枝數量的相關性
注:**在0.01水平(雙側)上顯著相關
Note:**at the 0.01 level (bilateral),a significant correlation
2.5種子與幼苗的補充及更新
自然狀態下北極花的胚珠有10多枚,但只有1枚能夠發育,授粉后10 d的胚珠在大小上僅略微膨大,幼果大小變化也不明顯。結實初期(自然授粉后半個月)結實率高達82.18% ± 1.48%,但后期逐步脫落。自然授粉后1個月觀測,果實及種子仍為綠色,結實率為35.64% ± 2.20%。當年果實完全脫落,無宿存現象。對10個大小不等的克隆斑塊進行幼苗補充狀況的調查,不見幼苗的更新。在居群的繁殖與更新中有性繁殖的后代適合度較低。
3討 論
3.1北極花的繁殖模式
克隆繁殖使得個體在有效攝取資源、規避風險方面具有重要作用,但是在資源有限的情況下,旺盛的克隆生長極易降低有性繁殖的資源投入。通常“游擊型”克隆植物能有效利用異質生境中的資源,具有較強的“覓食行為”,但有性繁殖能力較低[18-20]。北極花居群的繁殖模式支持了這一觀點。
北極花的分株長度達(60.14 ± 5.70)cm,但其分枝數量并不多,且在單株水平上有性枝數量(2.50 ± 0.30)枝明顯低于營養枝數量(8.23 ± 1.18)枝,在分株生物量統計中,有性繁殖的生物量分配比例僅占7.06% ± 1.30%,遠低于分株匍匐莖和營養枝的生物量,后者的生物量之和占90%以上。自然界中,植物的生殖分配比例是相對穩定的,通常一年生植物生殖分配比例達20%~40%,多年生植物占20%左右,人工選擇下的栽培植物可達40%左右[21]。相比之下,北極花的有性繁殖分配比例要低得多。
居群中實生苗的補充狀況是衡量有性繁殖適合度的主要參數之一,盡管統計顯示每分株有果實(1.47 ± 0.29)個,但在野外觀測中幾乎沒有看到實生苗的補充。從兩種繁殖方式的適合度看,北極花居群的擴展更多的依賴于分株匍匐莖的伸長和營養枝的分化。
3.2北極花的有性繁殖
植物繁殖與最適生活史格局的形成通常受植物資源的分配模式,尤其是繁殖過程中資源的分配模式影響[22]。生活史理論預測:不同生活史類型的植物各性狀之間的資源分配存在著“此消彼長”的權衡關系[11]。但這種權衡又受資源條件、生長時期和遺傳因素等的影響而表現各異。北極花在分株和居群水平上,其有性繁殖的衡量參數果實數僅與有性分枝數量和結實率相關,而表現出與對繁殖適合度有較大貢獻的其他性狀不相關,沒表現出明顯的權衡關系。權衡關系的出現取決于可利用的資源的量,只有當資源存在限制,而且依賴于相同的限制資源時,才能表現出來。
北極花過低的有性繁殖力可能在資源分配方面不足以對分株產生影響,或者與其他性狀之間依賴于不同的限制性資源,進而表現出與其他性狀的不相關。
有性繁殖對克隆植物居群遺傳結構貢獻的研究在不同類群中差異很大。一些研究表明較低的有性繁殖使得居群處于瀕危狀態[4-6];而又有一些研究則認為能夠產生大量種子并不意味著是以有性繁殖為主,如果幼苗成功更新的條件存在,不頻繁的種子生產也可能維持高的遺傳多樣性[23,24]。為此,多種機制被提出用于解釋有性繁殖對克隆植物遺傳結構的貢獻,其中種群幼苗補充機制假說認為:克隆植物居群遺傳多樣性的維持可通過建群早期的幼苗補充、發育過程中的重復幼苗補充和通過干擾發生的陣發式幼苗補充三種途徑來實現。其中,初期幼苗補充和陣發式幼苗補充在居群中都不會看到有高的幼苗補充[25,26]。這一假說較好的解釋了一些居群有著高的遺傳多樣性,卻沒有高的幼苗補充的現象。北極花屬于哪種情況還有待于對其進一步研究。
觀測點位于北極花亞歐地理分布區的邊緣,依據遺傳不育進化的流行假說:居群中一些對適合度不再有貢獻的性狀,往往可能會通過突變而逐步退化[27]。北極花居群過低的有性繁殖率如果對居群適合度不再有貢獻或貢獻率過低,那么其有性繁殖就有通過不育突變累積而退化的風險。
4結 論
4.1北極花具有較強的克隆生長力和相對低的有性繁殖狀態,居群的擴展更多依賴于分株匍匐莖的伸長和營養枝的分化。
4.2有性繁殖的衡量參數果實數僅與有性分枝數量和結實率相關,過低的有性繁殖力可能在資源分配方面不足以對分株產生影響,或者與其他性狀之間依賴于不同的限制性資源,進而表現出與對繁殖適合度有較大貢獻的其他性狀不相關。
4.3依據遺傳不育進化的流行假說,如果北極花的有性繁殖對維持居群遺傳多樣性的貢獻率過低,那么其有性繁殖有通過不育突變而衰退的風險。這也是很多物種瀕危的重要原因之一。
參考文獻(References)
[1]張大勇. 植物生活史進化與繁殖生態學[M]. 北京: 科學出版社, 2004.
ZHANG Da-yong. (2004).Lifehistoryevolutionandreproductionecologyinplants[M]. Beijing: Science Press. (in Chinese)
[2] Eriksson, O. (1997).Clonallifehistoriesandtheevolutionofseedrecruitment. [C]//. In: de Kroosn H & van Groenendael (eds). The ecology and evolution of clonal plants. Leiden: Backhuys Publishers. pp. 66.
[3] 張玉芬, 張大勇. 克隆植物的有性與無性繁殖對策[J]. 植物生態學報, 2006, 30(1): 174 - 183.
ZHANG Yu -fen, ZHANG Da-yong. (2006). Asexual and sexual reproductive strategies in clonal plants [J].JournalofPlantEcology, 30(1): 174 - 183. (in Chinese)
[4] Fischer, M., Hock, M., & Paschke, M. (2003). Low genetic variation reduces cross-compatibility and offspring fitness in populations of a narrow endemic plant with a self-incompatibility system.ConservationGenetics, 4(3):325-336.
[5] Aigner, P. A. (2004). Ecological and genetic effects on demographic processes: pollination, clonality and seed production in dithyrea maritima.BiologicalConservation, 116 (1):27-34.
[6] Scobie, A. R., & Wilcock, C. C. (2009). Limited mate availability decreases reproductive success of fragmented populations of linnaea borealis, a rare, clonal self-incompatible plant.AnnalsofBotany, 103 (6):835-846.
[7] Andrew, G. (1999). Paternal bottlenecks in fragmented populations of the grassland daisy rutidosis leptorrhynchoides.GeneticsResearch, 73:111-117.
[8] Willi, Y., Van, B. J., & Fischer, M. (2005). A threefold genetic allee effect: population size affects cross-compatibility, inbreeding depression and drift load in the self-incompatible ranunculus reptans.Genetics,169 (4):2,255-2,265.
[9] Willi, Y., Buskirk, J. V., & Hoffmann, A. A. (2006). Limits to the adaptive potential of small populations.AnnualReviewofEcologyEvolution&Systematics, 37:433-458.
[10] Totlsand. (2001). Environment-dependent pollen limitation and selection on floral traits in an alpine species.Ecology, (82): 2,233-2,244.
[11] 董鳴. 克隆植物生態學[M]. 科學出版社, 2011:67-68.
DONG Ming. (2011).ClonalPlantEcology[M]. Beijing: Science Press:67-68. (in Chinese)
[12] Wilcock, C. C., & Jennings, S. B. (1999). Partner limitation and restoration of sexual reproduction in the clonal dwarf shrub linnaea borealis, l. (caprifoliaceae).UniversityCollegeLondon, 208 (1):76-86.
[13] Scobie, A. R., & Wilcock, C. C. (2009). Limited mate availability decreases reproductive success of fragmented populations of linnaea borealis, a rare, clonal self-incompatible plant.AnnalsofBotany, 103 (6):835-846.
[14] Zhang, A. Q., Xiong, Y. Z., Huang, S. Q. (2014). Maintenance of self-incompatibility in peripheral populations of a circumboreal woodland subshrub.AoBPlants, 6(3):141-145
[15]侯真珍, 何爽, 王風雷, 等. 珍稀克隆植物北極花有性繁殖動態及影響因素[J]. 生態學雜志,2013,(12):3 167-3 172.
HOU Zhen-zhen, HE Shuang, WANG Feng-lei, et al. (2013). Dynamics of rare clonal plant Linnaea borealis L. sexual reproduction and related affecting factors [J].ChineseJournalofEcology, (12):3,167-3,172. (in Chinese)
[16] Neiland, M. R. M., Wilcock, C. C. (1997). Reproductive ecology and conservation of rare plant species in Scotland [J].BotanySocial.ScotlandNews, 68:18-19.
[17] 尹林克. 新疆珍稀瀕危植物[M]. 烏魯木齊: 新疆科學技術出版社, 2006.
YIN Lin-ke. (2006).RareendangeredendemicplantsinXinjiangofChina[M]. Urumqi: The Publish House of Xinjiang Science Technology. (in Chinese)
[18] Sculthorpe, C. D. (1967). The biology of aquatic vascular plants [M].EdwardArnold, London.
[19] Philbrick, C. T., & Les, D. H. (1996). Evolution of aquatic angiosperm reproductive systems. Bioscience, 46(11):813-826.
[20] Marcel E. Dorken +, & Eckert, C. G. (2001). Severely reduced sexual reproduction in northern populations of a clonal plant, decodon verticillatus, (lythraceae).JournalofEcology, 89 (3):339-350.
[21] 周紀倫. 植物種群生態學[M]. 北京: 高等教育出版社, 1992.
Zhou Ji-lun. (1992).PlantPopulationEcology[M]. Beijing: Higher Education Press. (in Chinese)
[22] 鐘章成. 植物種群的繁殖對策[J]. 生態學雜志, 1995,14(1):37 - 42.
ZHONG Zhang-cheng. (1995). Reproductive strategies of plant populations [J].ChineseJournalofEcology, 14(1):37-42. (in Chinese)
[23] Bengtsson, B. O. (2003). Genetic variation in organisms with sexual and asexual reproduction.JournalofEvolutionaryBiology, 16(2):189-199
[24] Silvertown ,J., Franco, M., Menges, E. (1996). Interpretation of elasticity matrices as an aid to the management of plant population for conservation [J].ConservationBiology, 10:591-597.
[25] Eriksson, O. (1989). Seedling dynamics and life histories in clonal plants.Oikos, 55(2):231-238.
[26] Eriksson, O. (1993). Dynamics of genets in clonal plants.TrendsinEcologyandEvolution, 8(9):313-316.
[27] Klekowski, E. J. (1988). Progressive cross- and self-sterility associated with aging in fern clones and perhaps other plants.Heredity, 61 (2):247-253.
Fund project:NSFC- Xinjiang joint fund project (U1203102); Key Laboratory of genetic engineering of biological resources in Xinjiang (XJDX0201-2011-08); Innovative Training Program for Undergraduate of Xinjiang University(XJU-SRT-14067)
doi:10.6048/j.issn.1001-4330.2016.06.020
收稿日期(Received):2016-01-19
基金項目:NSFC-新疆聯合基金項目(U1203102); 新疆生物資源基因工程重點實驗室項目(XJDX0201-2011-08); 新疆大學大學生創新訓練項目(XJU-SRT-14067)
作者簡介:翟雅芯(1990 - ), 女, 河南郾城人, 碩士研究生, 研究方向為繁殖生態學,(E-mail)1435605732@qq.com 通訊作者(Cotresponding author):何爽(1970 - ), 女, 講師, 研究方向為繁殖生態學, (E-mail)heshuang18@yahoo.com 張愛勤(1970 - ), 女, 河南南陽人, 教授, 碩士生導師, 研究方向為繁殖生態學, ( E-mail)zaql@sohu.com
中圖分類號:Q94-1
文獻標識碼:A
文章編號:1001-4330(2016)06-1122-07
The Sexual Reproduction and Plant Strategies for Resource Acquisition and Mass Partitioning in Rare Clonal PlantLinnaeaborealisL.
ZHAI Ya-xin, WANG Yue, WEI Xian-xian, HE Shuang, ZHANG Ai-qin
(CollegeofLifeScienceandTechnology,XinjiangUniversity,Urumqi830046,China)
Abstract:【Objective】 Sexual reproduction has an important influence on clonal plant life history, for it increases the potential ability of clonal plant adapting to the changeable environment.【Method】To understand interactions among component factors contributing to plant sexual reproduction, and resource condition in terms of distribution and partitioning, we studied sexual reproduction effort and some related agronomical traits such as sizes of clonal ramets, no. of fruit, assimilation shoots and flowering shoots per clonal ramet, biomass allocation ratio of clonal ramets, no. of flowering shoots, fruit set ratio of different size clonal patch and so on in Linnaea borealis by observation in the wild and measurement indoor. 【Result】The results showed the clonal ramets was (60.14 ± 5.70) cm length, with assimilation shoots of (8.23 ± 1.18),flowering shoots of (2.50 ± 0.30) and fruits of (1.47 ± 0.29) per clonal ramet. The sexual reproduction fitness was affected directly by sexual branch number, and not affected by ramet length and biomass at ramet level by correlation analysis. The assimilation shoots, the assimilation organs of cloned ramets, affected indirectly sexual reproduction fitness of population by impacting on the number of flowering shoots. At population level, the number of fruit was correlated significantly with the sexual branch number and fruit set rate with the correlation coefficient of 0.849, 0.847 respectively (P<0.01), and no correlation with clonal patch size. 【Conclusion】L. borealis had relatively low sexual reproduction effort. The fruit number, the measure parameters of sexual reproduction, was only related to sexual branch number and fruit set rate, and no correlation with other traits which have greater contribution to offspring fitness such as sizes of clonal ramets, no. of assimilation shoots, ramets biomass and clonal patch size. On the basis of the evolution hypothesis of genetic sterility, the sexual reproduction would have the risk of degradation by sterility mutations in L. borealis natural populations, if the genetic diversity of population were at a lower level due to the low contribution rate of sexual reproduction.
Key words:Linnaea borealis L.; clonal plant; sexual reproduction; component parameters; correlation analysis
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
