8個新疆扁桃S-基因多態性與生物信息學研究
2016-07-23 01:58:54郭春苗楊波吐迪麥麥提李寧唐亞萍龔鵬王繼勛
新疆農業科學 2016年6期
郭春苗,楊波,吐迪·麥麥提,李寧,唐亞萍,龔鵬,王繼勛
(1新疆農業科學院園藝作物研究所/農業部新疆地區果樹科學觀測試驗站,烏魯木齊 830091;2新疆莎車縣林業局,新疆莎車 844700)
?
8個新疆扁桃S-基因多態性與生物信息學研究
郭春苗1,楊波1,吐迪·麥麥提2,李寧1,唐亞萍1,龔鵬1,王繼勛1
(1新疆農業科學院園藝作物研究所/農業部新疆地區果樹科學觀測試驗站,烏魯木齊830091;2新疆莎車縣林業局,新疆莎車844700)
摘要:【目的】自交不親和基因(S-基因)在果樹及授粉樹選配和自交親和性品種選育等方面具有重要意義,是影響扁桃繁殖性狀的重要候選基因,對8個新疆扁桃S-基因序列、基因結構、基因多態性進行研究。【方法】采用Blast、ProtParam和SignalP等生物信息學軟件,對8個新疆扁桃S-基因的理化性質、多態性、系統發生樹和蛋白結構等進行分析。【結果】新疆扁桃S氨基酸存在信號肽,不是分泌性蛋白,具有跨膜結構,推測S氨基酸可能定位于細胞膜中發揮功能;通過進化樹分析,分為兩大類群;二級結構由無規則卷曲、α螺旋和延伸鏈等結構元件組成。【結論】8個新疆扁桃S-基因的理化性質、多態性、系統發生樹和蛋白結構等生物信息學分析結果將為S-基因的酶學特性及自交不親和基因分子機理研究提供理論參考。
關鍵詞:扁桃;S-基因;多態性;生物信息學分析
0引 言
【研究意義】扁桃(AmygdaluscommunisL.),又名巴旦杏,屬于薔薇科(Rosaceae)李亞科桃屬扁亞屬落葉喬木,為世界著名干果及木本油料樹種。新疆南部具有扁桃生長發育的氣候條件和資源優勢,是我國扁桃的主產區[1]。自交不親和性是高等植物在進化過程中為保持遺傳變異特性逐漸形成的一種重要機制,而扁桃普遍具有配子體自交不親和性,品種間S基因型的差異性導致授粉親和力不穩定、結實率低,生產上需合理配置授粉樹以提高產量[2]。因此,扁桃S-基因的研究,對栽培生產、遺傳改良具有重要的理論意義和實踐價值。【前人研究進展】依據控制因子不同,自交不親和類型分為孢子體自交不親和(Sporophytie self-incompatibility, SSI)和配子體自交不親和類型(Gametophytic self-incompatibility,GSI)[3]。蘋果、扁桃、梨等薔薇科果樹通常屬于配子體型自交不親和性植物[4],該類型是由復等位基因的單一位點即S-基因決定。GSI花粉管常常在花柱傳遞組織,特別是花柱中上部1/3處停止生長,這些表型由花粉本身的單倍體基因所決定[5]。基于S基因型不相同的品種之間授粉親和力強、結實性高的原理,運用科學鑒定的S-基因型來確定合理的授粉樹品種是科學高效的方法,因此研究自交不親和性果樹的S-基因對栽培中品種搭配和遺傳育種具有重要的指導意義[6-7]。【本研究切入點】利用生物信息學技術對目的基因進行序列分析,是初步確定基因結構及預測其生物學功能的有效方法,對新疆喀什當地品種進行S-基因多態性與生物信息學分析還鮮見報道。【擬解決的關鍵問題】研究采用生物信息學分析方法,對8個新疆扁桃S-基因理化性質、結構特征和功能域等進行分析,揭示該物種S-基因的結構,并對其生物學功能進行預測分析,為新疆扁桃分子生物學研究及自交親和性育種奠定理論基礎。
1材料與方法
1.1材 料
以本課題前期克隆得到的8個新疆扁桃品種的S-基因序列為基礎,結合從Genbank數據庫(http://www.ncbi.nlm.nih.gov/genbank/)中下載櫻桃、歐洲李、矮扁桃等植物的S-基因序列進行序列分析。表1
1.2方 法
對8個克隆、測序比對正確的扁桃S-基因序列及其編碼蛋白采用表1中所列軟件進行生物信息學分析,對其結構、理化特性和生物學功能進行分析和預測。

表1 生物信息學分析軟件信息
2結果與分析
2.1扁桃S-基因序列理化性質
用Prot Param在線工具分析8個新疆扁桃品種S-基因的核苷酸及其氨基酸序列。研究表明,氨基酸序列長度在198~257 aa,分子量在23 412.9~30 720.8 Da,理論等電點在8.62~10.30,均為不穩定性蛋白。亞細胞定位分析表明除了白薄殼S蛋白在細胞膜上,其余主要定位在質膜上。利用SMART分析S蛋白的功能域,表明S蛋白均具有一個RNase-T2功能域,屬于RNase蛋白家族。
對S蛋白的親水性/疏水性分析顯示,親水區域顯著大于疏水區域,說明S蛋白是親水性蛋白,其親水性較強。該蛋白均存在1~3個跨膜區,屬于跨膜蛋白,均不存在信號肽,不是分泌性蛋白。蛋白磷酸化位點分析表明,多數S蛋白具有1~4個絲氨酸位點,其中雙薄、葉爾羌、白長巴旦S蛋白不具有相對保守的絲氨酸位點。表2
2.2S蛋白同源性比較
利用DNAman對8個新疆扁桃S-基因氨基酸序列進行同源性比對。研究表明,葉爾羌和雙軟同源性最高,達到99.5%,苦巴旦和黃雙的同源性為99.2%,白長巴旦與葉爾羌的同源性高達99%,白長巴旦與雙軟的同源性高達98.5%;雙軟和黃雙,白長巴旦和黃雙,白長巴旦和苦巴旦的同源性較低,僅為21.3%。其余品種之間同源性也比較集中,一部分在23%~31%,另一小部分為63%左右。說明同源性之間有高度相似的部分,但差異也是顯著的。觀察圖形化的多重序列比對結果,證實了上述同源性比對的結果。通過Weblogo圖可以看出,不同品種扁桃的S蛋白具有保守結構域YFQFVQQWPPT。圖2,圖3,表3

表2 8個新疆扁桃品種的S蛋白理化性質
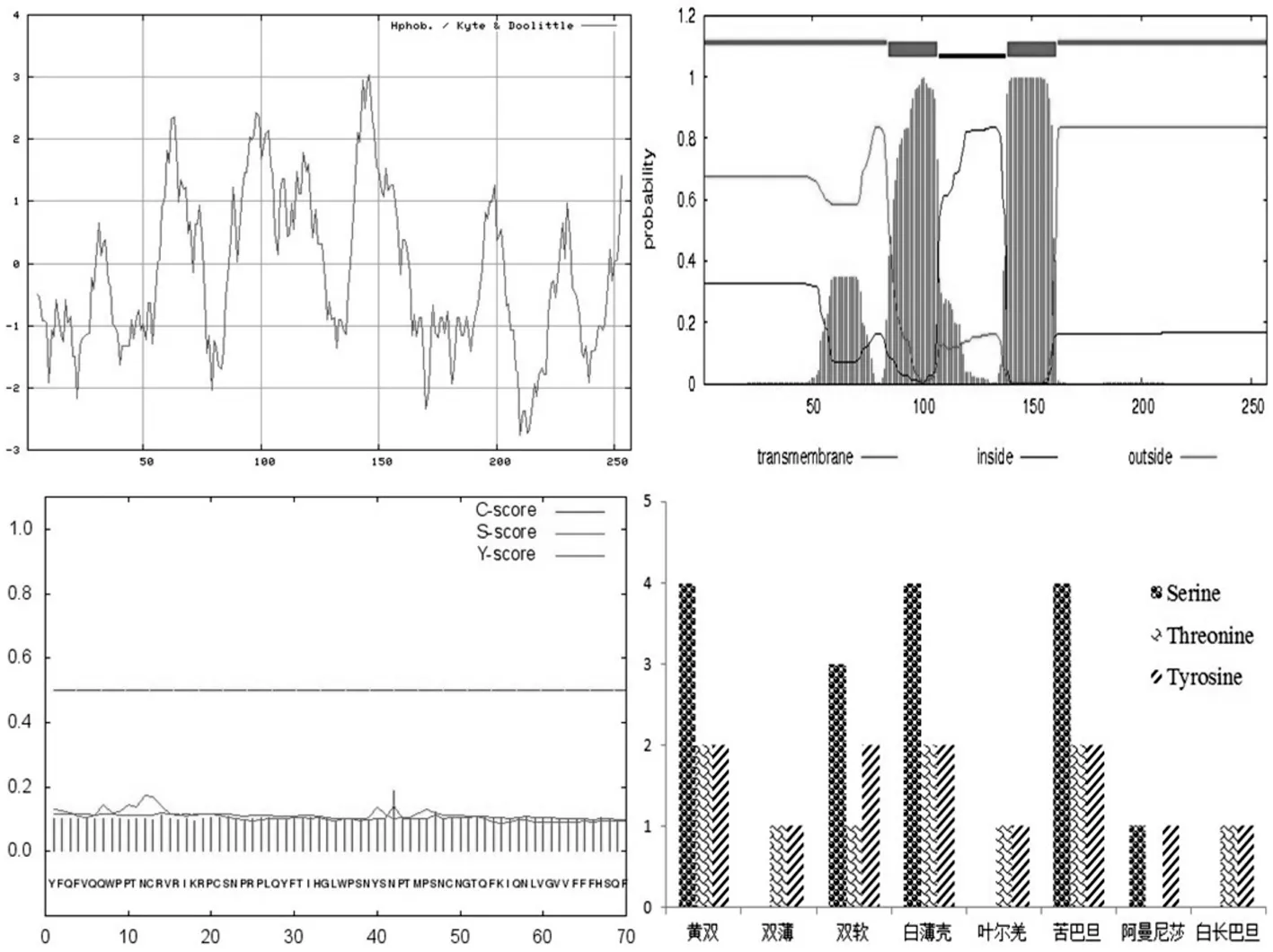
圖1 黃雙扁桃S-基因序列理化性質
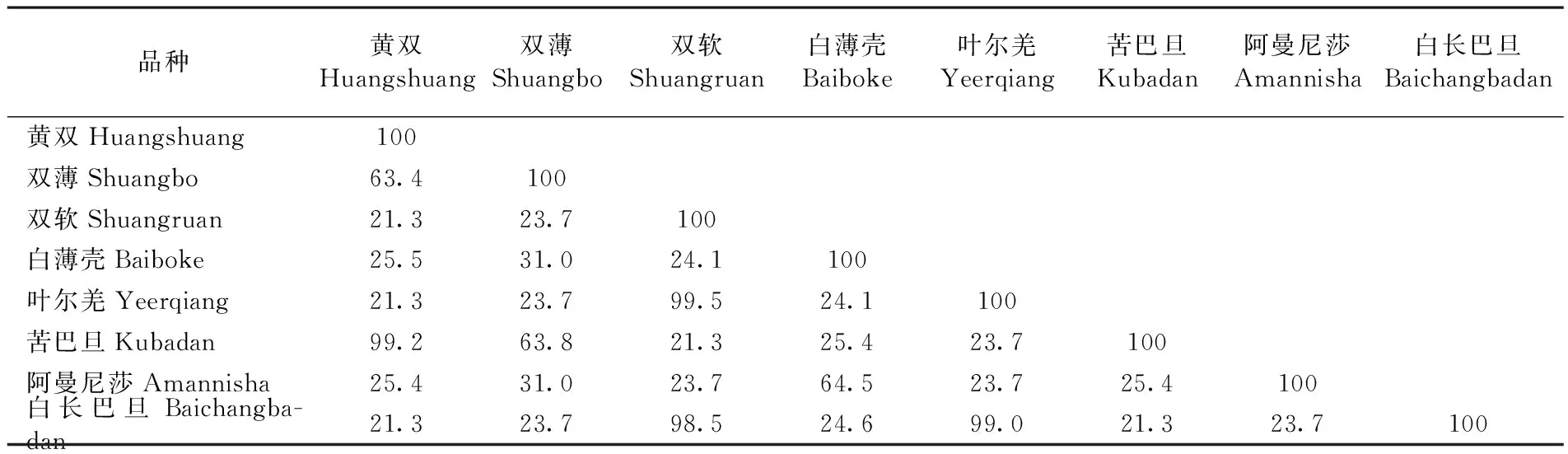
品種黃雙Huangshuang雙薄Shuangbo雙軟Shuangruan白薄殼Baiboke葉爾羌Yeerqiang苦巴旦Kubadan阿曼尼莎Amannisha白長巴旦Baichangbadan黃雙Huangshuang100雙薄Shuangbo63.4100雙軟Shuangruan21.323.7100白薄殼Baiboke25.531.024.1100葉爾羌Yeerqiang21.323.799.524.1100苦巴旦Kubadan99.263.821.325.423.7100阿曼尼莎Amannisha25.431.023.764.523.725.4100白長巴旦Baichangba-dan21.323.798.524.699.021.323.7100

圖2 8種扁桃推導S氨基酸序列比對

圖3 8種扁桃S氨基酸序列Logo圖譜
2.3S蛋白系統進化樹
用ClustalX1.83和MEGA6.0.5軟件對上述18種植物的S氨基酸序列構建系統進化樹,采用默認參數,自檢舉1 000次,對生成的系統樹進行Bootstrap校正。研究表明,在同一個進化分支上,18個物種分為三大類,新疆本土扁桃S基因分為兩個類群。其中,阿曼尼沙、白薄殼、雙軟、黃雙、苦巴旦在一枝。白長巴旦、雙薄和葉爾羌在另一枝,與巴旦杏距離較近。剩余的物種為一類,這與同源性比較的結果相一致。圖4
2.4S蛋白二級結構
通過NPS程序對8種扁桃S蛋白序列進行二級結構分析表明,S蛋白均由α螺旋、β折疊、無規則卷曲和延伸鏈等結構元件組成,其中無規則卷曲所占比例最高,在39.6%~51.75%,其次為α螺旋或延伸鏈,β折疊所占比例最小。在不同扁桃品種的S蛋白中,4種結構元件的百分比和分布均不同,這可能與不同品種中的S蛋白所具有的具體生物學功能不同有關。表4

圖4 18種植物S蛋白的系統進化

植物種類PlantSpecies二級結構元件比例Theproportionofsecondarystructureelements(%)α螺旋Alphahelix延伸鏈Extendedstrand無規則卷曲Randomcoil二級結構元件分布Secondarystructureelementsdistribution黃雙Huanghuang30.3525.6843.97雙薄Shuangbo40.9112.1246.97雙軟Shuangruan24.7123.5351.76白薄殼Baiboke34.5115.0450.44葉爾羌Yerqiang40.4012.6346.97苦巴旦Kubadan33.8522.5743.58阿曼尼莎Amannisha47.612.839.6白長巴旦Baichangbadan37.3712.6350
2.5S蛋白三級結構分析
利用Swiss-Model采用同源建模的方法預測不同扁桃品種S蛋白的三級結構。S蛋白三級結構主要是由α螺旋、延伸鏈和無規則卷曲等二級結構元件構成,不同扁桃品種的S蛋白三級空間結構相似度高,保守性強。圖5
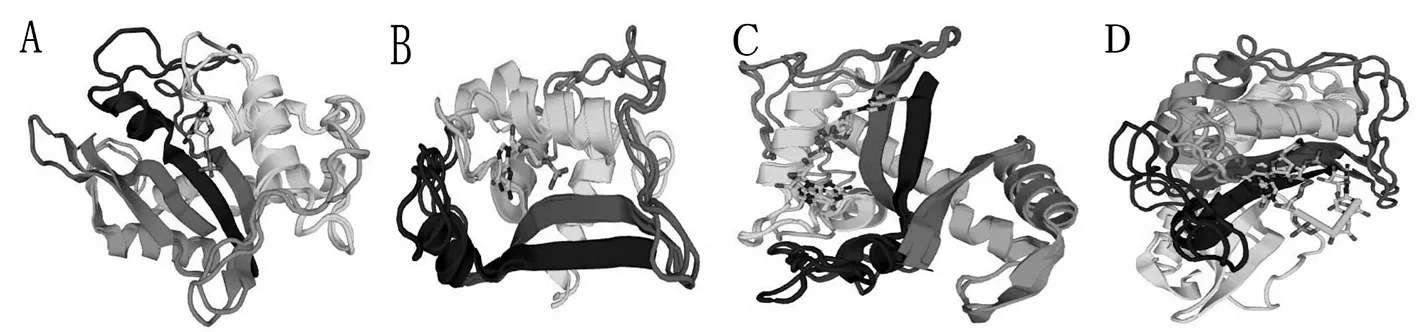
A.阿曼尼沙;B.雙軟;C.黃雙;D.白長巴旦
3討 論
S-RNase是一種S糖蛋白,具有核酸酶活性,又稱為S-核酸酶,其氨基酸同源性為38%~98%,其不同S-單倍型核酸酶具有高度多態性,而高變區的多態性為不同S-RNase的鑒定依據[8]。對櫻桃[9]、蘋果[10]梨[11]和杏等[12]果樹S-RNase的編碼cDNA序列進行對比分析發現,果樹S-RNase基因的結構特征存在極高的相似性。研究通過課題RT-PCR獲得了8個新疆扁桃S-基因的cDNA序列,對該基因編碼的蛋白氨基酸序列進行分析,所獲得cDNA序列編碼合成的S-基因氨基酸序列包含氨基酸數目在198~257 aa,自成兩個類群,白長巴旦,雙薄、葉爾羌為一個類群,黃雙、雙薄、雙軟、白薄殼、苦巴旦、阿曼尼莎為另一個類群。
對8個新疆扁桃S-基因進行一系列生物信息學分析表明,S-基因均具有一個典型的RNase-T2功能域,屬于RNase蛋白家族[13]。序列比對發現扁桃S-基因與其他植物物種的S-基因家族具有極高的相似性,其中該類蛋白的N端保守結構區域相似性極高,而其C端的質體信號肽序列區差異性則較大,這表明不同品種扁桃間S蛋白功能的差異性可能通過其特異的C端序列發揮作用。
4結 論
S氨基酸序列理化性質分析顯示,這8個S-基因理論等電點pI在8.62~10.30,脂肪指數基本上比較凌亂,最小的65.09,最大的86.31,均為較不穩定蛋白,均富含絲氨酸;系統進化分為2枝,黃雙、雙薄、雙軟、白薄殼、苦巴旦、阿曼尼莎聚在一分枝上,其他的聚在另一分枝上;均不存在信號肽,不是分泌性蛋白,均存在1~3個跨膜區,屬于跨膜蛋白,定位在在細胞膜上;S氨基酸序列的二級結構中,α-螺旋和無規則卷曲是S多肽鏈中的大量結構元件;整個肽鏈中,呈無規則卷曲>α-螺旋>延伸鏈>β-轉角的規律,其中無規則卷曲占主導地位。
參考文獻(References)
[1]李疆,曾斌,羅淑萍,等.中國野扁桃資源的保護及引種繁育[J].新疆農業學,2006,43(1):61-62.
LI Jiang,ZENG Bin,LUO Shu-ping,et al.(2006).Protection and Propagation of Amygdalus Ledebouriana Schleche in China [J].XinjiangAgriculturalSciences,43(1):61-62.(in Chinese)
[2]呂志江,李疆,吾買爾夏提·塔漢,等.新疆野扁桃種質資源遺傳多樣性的ISSR分析[J].果樹學報,2010,27(6):918-923.
LV Zhi-jiang,LI Jiang,Wumaierxiati Tahan,LUO Shu-ping,et al.(2010).ISSR analysis for genetic diversity of Amygdalus ledebouriana germplasm from Xinjiang,China [J].JournalofFruitScience,27(6):918-923.(in Chinese)
[3] Ken-ichi, Kubo, Tetsuyuki, Entani, Akie, Takara, Ning, Wang, Allison M, Fields, & Zhihua, Hua. (2010). Collaborative non-self recognition system in s-rnase-based self-incompatibility.Science, 330(6005):796-799.
[4] Ivanov, R., Fobis-Loisy, I., & Gaude, T. (2010). When no means no: guide to brassicaceae self-incompatibility.TrendsinPlantScience, 15(7):387-394.
[5] Scalone, R., & Albach, D. (2014). Cytological evidence for gametophytic self-incompatibility in the genus veronica.TurkishJournalofBotany, 38(1):197-201.
[6] Certal, A. C., Sanchez, A. M., Kokko, H., Broothaerts, W., Oliveira, M. M., & Feijó, J. A. (1999). S-rnases in apple are expressed in the pistil along the pollen tube growth path.SexualPlantReproduction, 12(2):94-98.
[7]郭長奎,李疆,羅淑萍,等.扁桃花粉SFB 基因的鑒定和序列分析[J].經濟林研究,2009,27(6):18-23.
GUO Chang-kui,LI Jiang,LUO Shu-ping,et al.(2009).Identification and Sequence Analysis of Pollen SFB Gene in Almond [J].NonwoodForestResearch,27(6):18-23.(in Chinese)
[8] Meng, D., Gu, Z., Wang, A., Yuan, H., Li, W., & Yang, Q., et al. (2014). Screening and characterization of apple rho-like gtpase (mdrops) genes related to s-rnase mediated self-incompatibility.PlantCellTissue&OrganCulture, 117(3):465-476.
[9]李曉,張紹鈴,吳俊,等.櫻桃品種S基因型及自交不親和性分子機制研究進展[J].生物技術通報,2006,(6):28-33.
LI Xiao,ZHANG Shao-ling,WU Jun,et al.(2006).Studies on S Genotypes and Molecular Mechanism of self-incompatibility in Cherry[J].BiotechnologyBulletin,(6):28-33.(in Chinese)
[10] Dong, M., Zhaoyu, G., Wei, L., Aide, W., Hui, Y., & Qing, Y., et al. (2014). Apple mdabcf assists in the transportation of s-rnase into pollen tubes.PlantJournal, 78(6):990-1,002.
[11]烏云塔娜,譚曉風,李秀根,等.梨自交不親和新基因S12-RNase的分離鑒定及序列分析[J].林業科學,2006,42(4):117-121.
WU Yun-tana,TAN Xiao-feng,LI Xiu-geng,et al.(2006).Isolation and Identification of S12-RNase Gene of Pyrus bretschneideri[J].ScientiaSilvaeSinicae,42(4):117-121.(in Chinese)
[12] Zhang, L., Chen, X., Chen, X., Zhang, C., Liu, X., & Ci, Z., et al. (2008). Identification of self-incompatibility ( s- ) genotypes of chinese apricot cultivars.Euphytica, 160(2):241-248.
[13] Collani, S., Alagna, F., Caceres, E. M., Galla, G., Ramina, A., & Baldoni, G., et al. (2012). Self-incompatibility in olive: a new hypothesis on the s-locus genes controlling pollen-pistil interaction.ActaHorticulture, 967(1):133-140.
Fund project:Supported by the Basic Science and Technology Research Support Funds of Non-profit Research Institutions of Xinjiang Uygur Autonomous Region "germplasm identification of S-genotypes based on local almond in Xinjiang."(KY2015072); NSFC "Studies on the relationship between physiological fruit drop and sucrose metabolism in "Zhipi" almond of Xinjiang"
doi:10.6048/j.issn.1001-4330.2016.06.013
收稿日期(Received):2016-02-06
基金項目:自治區公益性科研院所基本科研業務經費資助項目“基于S-基因型鑒定的新疆本地扁桃種質資源鑒定(KY2015072)”;國家自然科學基金項目“新疆‘紙皮’扁桃幼果生理脫落與蔗糖代謝關系的生理與分子機制研究”
作者簡介:郭春苗(1982-),女,助理研究員,碩士,研究方向為新疆特色果樹生物技術改良,(E-mail)chunmiaoguo@126.com 通訊作者(Cotresponding author):王繼勛(1965-),男,研究員,研究方向為果樹資源收集與種質創新,(E-mail)ee_wjx@163.com
中圖分類號:S662.9
文獻標識碼:A
文章編號:1001-4330(2016)06-1067-07
Studies on Polymorphism and Bioinformatics of 8 Xinjiang Almonds Self-incompatibility Gene (S-gene)
GUO Chun-miao1, YANG Bo1, Tudi Maimaiti2, LI Ning1, TANG Ya-ping1,GONG Peng1, WANG Ji-xun1
(1ResearchInstituteofHorticulturalCrops,XinjiangAcademyofAgriculturalSciences/XinjiangFruitScienceExperimentStation,MinistryofAgriculture,Urumqi830091,China;2.ShacheCountyForestryBureau,ShacheXinjiang844700,China)
Abstract:【Objective】 Self incompatibility gene (S-gene) is an important candidate gene affecting reproductive traits of almond. It has important significance in the selection of self compatible varieties of fruit tree and the selection of pollination tree. This paper is preliminary studies on S-gene sequence, gene structure, gene polymorphism of 8 Xinjiang almond.【Method】This experiment used the Blast, Protparam and Signalp bioinformatics software to analyze the physical and chemical properties, polymorphism, phylogenetic tree and protein structure of 8 Xinjiang almond S-gene.【Result】The S amino acid of Xinjiang almond contained signal peptide and had a membrane structure, but it wasn't a secretory protein. It has been assumed that S amino acid may be located in the cell membrane to play a function. Through phylogenetic tree analysis,they were divided into two groups;The secondary structure was composed of irregular coil, alpha helix, extended chain, etc.【Conclusion】The analysis results of S-gene sequence, gene structure, gene polymorphism of 8 Xinjiang almond will provide a theoretical reference for the enzymatic properties of S- gene and the molecular mechanism of self incompatibility gene.
Key words:almond;self-incompatibility gene;polymorphism;bioinformatics analysis
