黃土高原先鋒種豬毛蒿葉片形態解剖與生理特征對立地的適應性
2016-07-21 01:10:38杜華棟焦菊英
生態學報 2016年10期
杜華棟, 焦菊英, 寇 萌, 苗 芳
1 西安科技大學地質與環境學院,西安 710054 2 中國科學院水利部水土保持研究所,楊陵 712100 3 西北農林科技大學生命科學學院,楊陵 712100
?
黃土高原先鋒種豬毛蒿葉片形態解剖與生理特征對立地的適應性
杜華棟1,2, 焦菊英2,*, 寇萌2, 苗芳3
1 西安科技大學地質與環境學院,西安710054 2 中國科學院水利部水土保持研究所,楊陵712100 3 西北農林科技大學生命科學學院,楊陵712100
摘要:為了揭示黃土丘陵溝壑區撂荒地植被演替前期優勢種豬毛蒿(Artemisia scoparia)對該區立地環境的適應性,探討豬毛蒿演替生態位的變化,研究了陜北黃土丘陵溝壑區3種立地環境下(陽峁坡、峁頂、陰峁坡)豬毛蒿葉片形態解剖和生理特征的變化,以及這些變化與生態因子之間的相互關系。結果表明:(1)豬毛蒿葉片具有適應該區半干旱環境的形態及解剖結構:葉片針形化、具表皮毛、環柵型葉肉組織、海綿組織特化為貯水組織、維管束退化、具裂生分泌腔,C3 植物呈現類似CAM 植物的葉片特性;(2)在土壤、空氣濕度相對干燥和強光生境的陽峁坡與峁頂,豬毛蒿具有較小的葉面積、發達的柵欄組織、致密的表皮毛和緊密的細胞間隙,而在生境條件較好的陰峁坡則呈相反趨勢;(3)陽峁坡豬毛蒿葉片相對含水量和葉綠素含量較小,超氧陰離子自由基增加,但植物體內超氧化物歧化酶和抗壞血酸含量增加以清除植物體內產生的活性氧;(4)冗余及相關性分析表明,豬毛蒿葉片形態、解剖和生理指標的可塑性對立地光照強度、土壤水分和有機質含量較為敏感,同時其形態解剖與生理可塑性可共同調節來適應生境。綜合分析,豬毛蒿對陜北黃土丘陵溝壑區撂荒初期光照強度大、土壤貧瘠但土層干化現象尚未出現的立地環境有較好的適應性,使其成為黃土高原植被自然演替過程中的先鋒物種。
關鍵詞:黃土高原;生態適應;葉片形態解剖;抗逆生理;豬毛蒿
豬毛蒿(Artemisiascoparia)是菊科蒿屬的一年生草本,是陜北黃土丘陵溝壑區退耕地撂荒后植被自然演替的主要先鋒物種[1],單個樣地覆蓋度可達50%以上[2],同時也可與苦荬菜 (Ixerispolycephala)、豬毛菜 (Salsolacollina)、阿爾泰狗娃花 (Heteropappusaltaicus)等構成共優群落,因此其在退耕初期控制坡面水土流失強度具有重要作用[3],同時其生長對土壤性質的改善對演替后期物種定植也有一定意義[4]。
植物在適應環境的過程中不但葉片內部外部形態特征發生了變化[5],而且還表現出一定的植物生理功能的可塑性[6]。這些可塑性使得植物在新的環境中能調節個體適應能力從而占有一定的生態位,顯示植物的生態適應和競爭能力[7],進而影響了植物群落的演替與更新[8]。黃土丘陵溝壑區嚴重的土壤侵蝕形成了復雜多變的立地環境,使得植物生長生態因子(光照、溫度、養分等)在空間上存在異質性[9],加之土壤侵蝕產生的土壤擾動[10],必然會對該地區植物的生長發育和植被演替更新產生影響。
目前學者對豬毛蒿水分脅迫下葉片解剖結構和生理變化[11]、種內種間競爭及其演替生態位變化[12]等方面進行了相關研究,然而在黃土丘陵溝壑區自然立地環境下,豬毛蒿在各生態因子的綜合效應下形態解剖特征與生理指標的可塑性研究報道不多[13],同時從植物形態、解剖與生理特征綜合研究植物適應環境策略及其生態學意義研究較為薄弱[14- 15]。因此,本研究選擇黃土丘陵溝壑區坡溝自然系統,分析不同立地環境各生態因子綜合作用下豬毛蒿葉片形態、解剖與生理指標的變化特征,并探索測定指標之間及其與環境生態因子之間的相互關系,并試圖探討豬毛蒿形態與生理可塑性在其生態位構建過程中的意義,為充實黃土丘陵溝壑區植物對環境的適應機制及土壤侵蝕區植被恢復提供科學依據。
1材料與方法
1.1研究區自然概況
研究區位于陜西省安塞縣紙坊溝小流域(圖1)。流域面積8.26 km2,氣候區劃上屬暖溫帶、半濕潤氣候向溫帶、半干旱氣候過渡的區域,年日照時間2415.6 h,年輻射量552.7 kJ/cm2,年均氣溫8.8 ℃,干燥度1.5。年平均降水量為542.5 mm且分布極不均勻,7、8、9月平均降雨量占年降雨量的61.1%,且多為暴雨,加上流域土質疏松的黃綿土,造成該流域嚴重的水土流失,溝壑密度高達8.06 km/ km2,屬典型的黃土丘陵溝壑地貌。
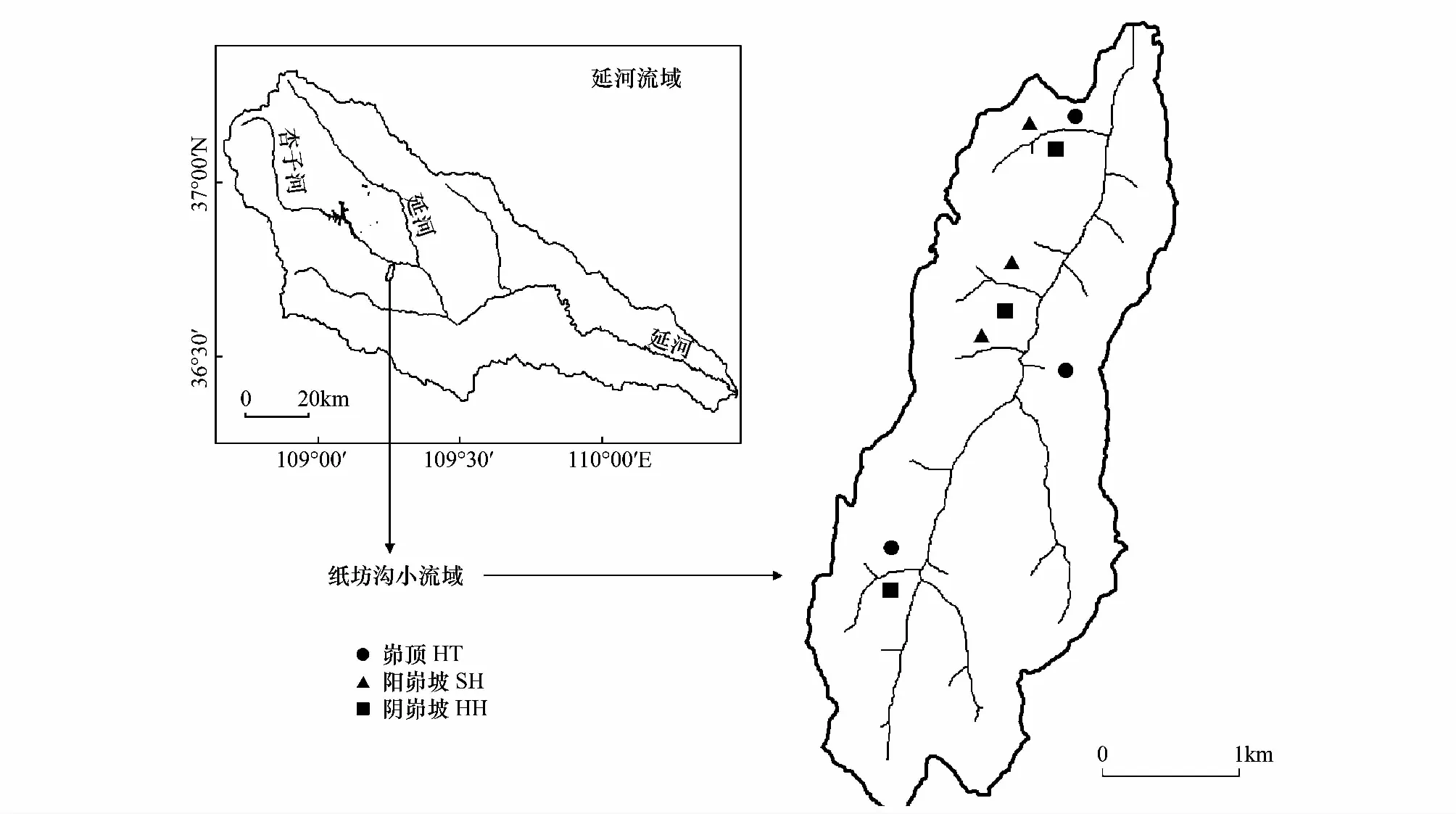
圖1 研究區位置圖Fig.1 Location of the experimental study area
1.2樣地選擇
坡溝系統作為黃土丘陵溝壑區小流域侵蝕產沙的基本單元,由于長期的侵蝕,其地貌形態、坡度及土壤侵蝕程度存在垂直變化。依據黃土丘陵溝壑區坡溝侵蝕類型和豬毛蒿的分布,在坡溝系統的陽峁坡(SH)、峁頂(HT)和陰峁坡(HH)3種不同立地環境進行調查取樣,且每種立地環境有3個退耕年限相似的豬毛蒿群落作為樣地重復,共9樣地(圖1)。首先每個樣地設置3個2 m×2 m樣方進行植被調查,調查樣方內出現的物種、數量及其蓋度,并剪取豬毛蒿地上生物量在60 ℃下24 h烘干稱重,豬毛蒿重要值=相對密度+相對頻度+相對蓋度,群落調查結果見表1。同時測定群落演替初期植物根系主要分布的0—30 cm土層的土壤養分和0—100 cm的土壤水分(垂直分布差異較大因此測量0—100 cm土層土壤水分),用上海嘉定學聯儀表廠生產的JD- 3型照度計測定豬毛蒿群落中的光照強度,樣地環境因子結果見表2。
1.3研究方法
每個樣地內在豬毛蒿群落中選擇植物個體作為取樣植株,選取樣株時盡量選擇大小一致的植株,10株重復。在每個植株上選擇從頂上第一枝向下第4枝上枝中的葉片,用剪刀剪下,置于兩片濕潤的濾紙之間,放入塑料袋內后封口,然后儲藏于黑暗的容器內(有冰袋,內部溫度<4 ℃)。回到室內,與已知面積的參照物同時照相,之后用Image pro-plus軟件分析照片得出葉面積。最后將葉片放入60 ℃烘箱內烘干48 h后取出稱重,計算用式(1)計算豬毛蒿比葉面積。

表1 樣地植被概況
數據以平均值(標準偏差)表示;不同字母表示不同樣地間差異顯著(0.05水平);相同的字母表示差異性不顯著(0.05水平); SH:sunny hill slope,陽峁坡;HT:hilltop,峁頂;HH:shady hill slope,陰峁坡
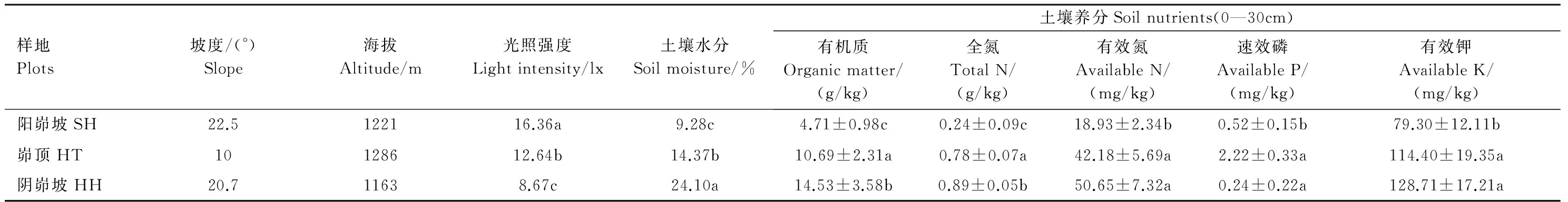
表2 樣地環境因子概況
葉片解剖在每株植物上葉片采樣時選取原則同葉片形態采樣,樣片采下后迅速放入FAA固定液中。實驗室采用常規石蠟切片方法,切片在Axio Scope A1數碼顯微圖像系統照相,后在Image pro-plus 軟件支持下分析,測量6種植物葉片角質層厚度、表皮細胞大小、葉片厚度、柵欄組織厚度、貯水組織厚度、木質部面積、韌皮部面積等,每個指標隨機選取50個進行測量,依據式(2、3、4、5)分別計算氣孔指數[16]、氣孔可塑性[17]、木質部韌皮部面積比、細胞間隙率[5]。
比葉面積
(1)
氣孔指數
(2)
氣孔可塑性
(3)
木質部韌皮部面積比
(4)
細胞間隙率
(5)
式中,SLA為比葉面積(m2/kg);LA為葉面積(m2);LM為干葉片質量(kg);SI為氣孔指數(%);STP為氣孔可塑性(%);STD為氣孔密度(個/μm2);CD為表皮細胞密度(ind/μm2);SWmax為氣孔完全張開寬度(μm);SWmin為氣孔閉合時寬度(μm);X/P為木質部韌皮部面積比(%);XA為木質部面積(μm2);PA為韌皮部面積(μm2);fias為細胞間隙率 (%);Am為葉肉組織橫切面面積(μm2);I為葉肉組織厚(μm);W為測量橫切面寬(μm)。
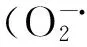
1.4數據處理
論文所有數據采用SPSS16.0軟件對數據進行統計檢驗,結果用平均值±標準偏差(SD)表示,用單因素方差分析(one-way ANOVA)和最小顯著差異法(LSD)比較不同數據組間的差異,差異顯著性水平設定為α=0.05;用 CANOCO4.5 軟件基于線性的冗余分析(RDA)分析豬毛蒿葉片形態、解剖和生理特征與環境因子的關系,相關顯著性用Monte Carlo 法進行檢驗,同時為使豬毛蒿葉片測量參數和環境因子的數據獲得正態分布,對所分析數據均進行lg(x+1) 轉換;用皮爾遜相關分析方法(Pearson Correlation Coefficient)分析葉片形態解剖特征與生理指標之間的相互關系。
2結果與分析
2.1不同立地環境下豬毛蒿葉片形態解剖特征
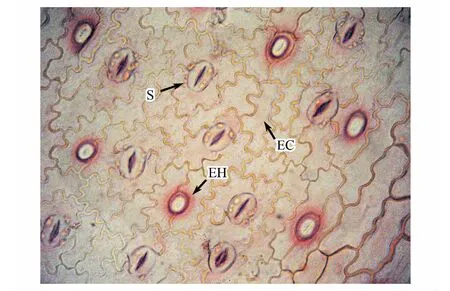
圖2 豬毛蒿葉片表皮特征Fig.2 Chacteristics of leaf epidermis of A. scoparia EC:表皮細胞 epidermal cell;EH:表皮毛著生位點 site of epidermal hair; S:氣孔 stomatal apparatus
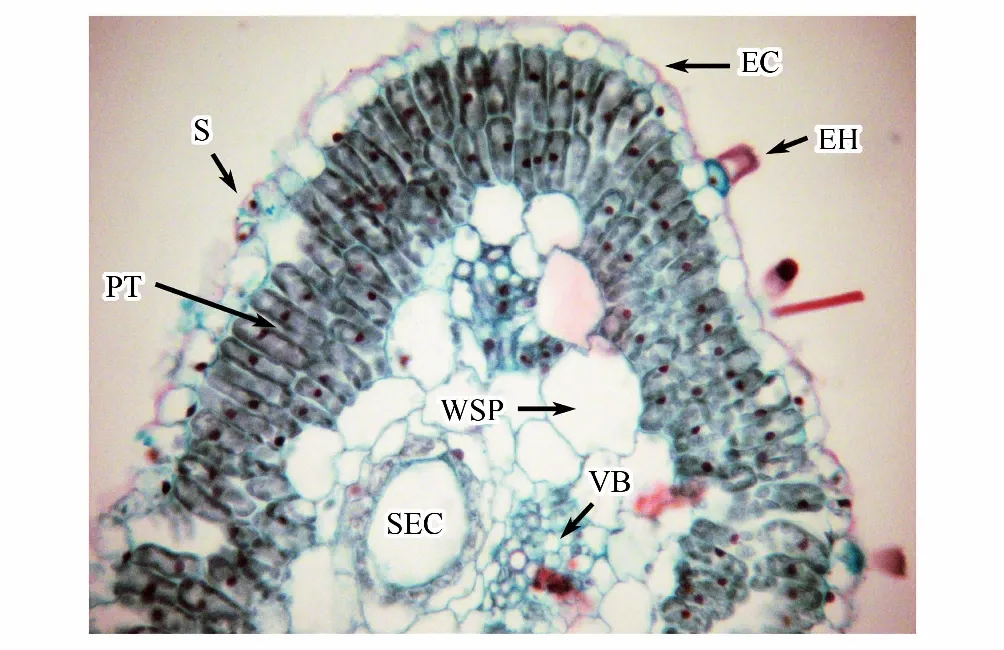
圖3 豬毛蒿葉片解剖特征Fig.3 Chacteristics of leaf anatomy of A. scoparia EC:表皮細胞 epidermal cell;EH:表皮毛著生位點 site of epidermal hair; PT:柵欄組織 palisade tissue;S:氣孔 stomatal apparatus;SEC:分泌腔 secretory cavity;VB:維管束 vascular bundle;WSP:貯水組織 water storage parenchyma
圖2、圖3分別顯示豬毛蒿葉片表皮及解剖特征(不同立地條件下豬毛蒿葉片表皮及解剖特征僅為測量統計上差異,并無特殊結構發育,因此文中僅選取了其陰峁坡表皮及解剖圖片作為示例),可以看出豬毛蒿葉表皮具表皮毛;表皮細胞形狀不規則,垂周壁為波狀彎曲,相互嵌合;氣孔復合體只有保衛細胞而無副衛細胞,普通表皮細胞不規則的圍繞在氣孔周圍,表皮細胞橫切面緊密排列;葉肉為環柵型,具有2—3層柵欄組織;海綿組織特化為貯水組織;葉肉細胞具裂生分泌腔;維管束外韌維管束但表現出一定的退化特征。
由圖4不同立地環境下豬毛蒿葉片形態與解剖變化統計可知,豬毛蒿葉面積、比葉面積、氣孔指數、表皮細胞大小、細胞間隙率、貯水組織厚、主維管束面積都呈現陰峁坡>峁頂>陽峁坡的趨勢,并表現出顯著性差異(P<0.05);而表皮毛密度、表皮角質層厚度、柵欄組織厚則表現出相反的趨勢,但陽峁坡和峁頂差異不顯著(P>0.05);氣孔可塑性、主維管束面積和木質部韌皮部面積比在3種立地環境下差異不顯著(P>0.05)。豬毛蒿葉片形態解剖測量的12個指標中,不同立地環境下變化量較大的前5個指標依次是細胞間隙、表皮毛密度、葉面積、氣孔指數和表皮角質層厚,其中細胞間隙、葉面積和氣孔指數在陰峁坡較陽峁坡分別增加了3.82、2.50倍和2.48倍,分別達到8.5%、3.82 cm2和26.52 m2/kg;而表皮毛密度和表皮角質層厚在陽峁坡較陰峁坡增加了3.06倍 和2.00倍,分別達到211.66 根/mm2和2.36 μm。
2.2不同立地環境下豬毛蒿生理特征
由圖5不同立地環境下豬毛蒿葉片生理指標變化特征可知,相對含水量和葉綠素含量在陰峁坡>峁頂、陰溝坡,并表現出顯著性差異(P<0.05);而相對可溶性蛋白含量、超氧陰離子自由基、超氧化物歧化酶和抗壞血酸則表現出相反的趨勢;所測豬毛蒿生理指標中,除可溶性蛋白和超氧化物歧化酶外,其他生理指標在陽峁坡和峁頂差異不顯著(P>0.05)。
由圖5統計結果可知,豬毛蒿葉片相對含水量由陽峁坡和峁頂的平均82.8%增加至陰峁坡的93.0%;豬毛蒿體內滲透調節物質可溶性蛋白含量由陽峁坡和峁頂的平均31.33 mg/g降低至陰峁坡的28.46 mg/g,雖然表現出差異顯著性,但其在立地環境的變化較其他測定生理指標小;表示植物細胞受傷害程度的超氧陰離子自由基在陽峁坡和峁頂平均達到17.15 μg/g,其在陰峁坡下降了3.2倍,迅速減小至5.35 μg/g;豬毛蒿體內抗逆酶SOD活性在陽峁坡最大為314.03 U g-1h-1,較峁頂和陰溝坡分別提高了24.4%和59.9%;非酶抗氧化物質抗壞血酸在由陽峁坡和峁頂的平均267.30 mg·100g-1減小至陰峁坡的182.40 mg/100 g;在陽峁坡和峁頂,豬毛蒿葉內葉綠素含量迅速由陰峁坡的6.09 mg/g減少了52.8%,下降至3.22 mg/g。
2.3豬毛蒿葉片形態解剖和生理指標與環境因子關系的RDA分析

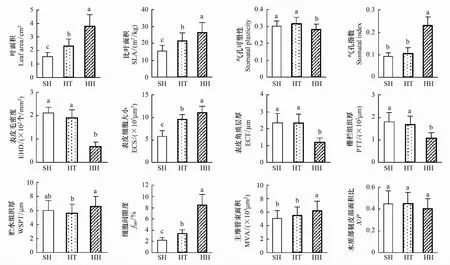
圖4 不同立地環境下豬毛蒿葉片形態與解剖變化特征Fig.4 Morphological and anatomical characteristics of A. scoparia at different sitesSH:陽峁坡 sunny hill slope;HT:峁頂 hilltop;HH:陰峁坡 shady hill slope;EHD: 表皮毛密度 epidermal hair density;ECS:表皮細胞大小 epidermal cells size;ECT:表皮角質層厚度 epidermal cuticular thickness;PTT:柵欄組織厚 palisade thickness;WSPT:貯水組織厚 water storage parenchyma thickness;MVA:主維管束面積 main vascular bundles
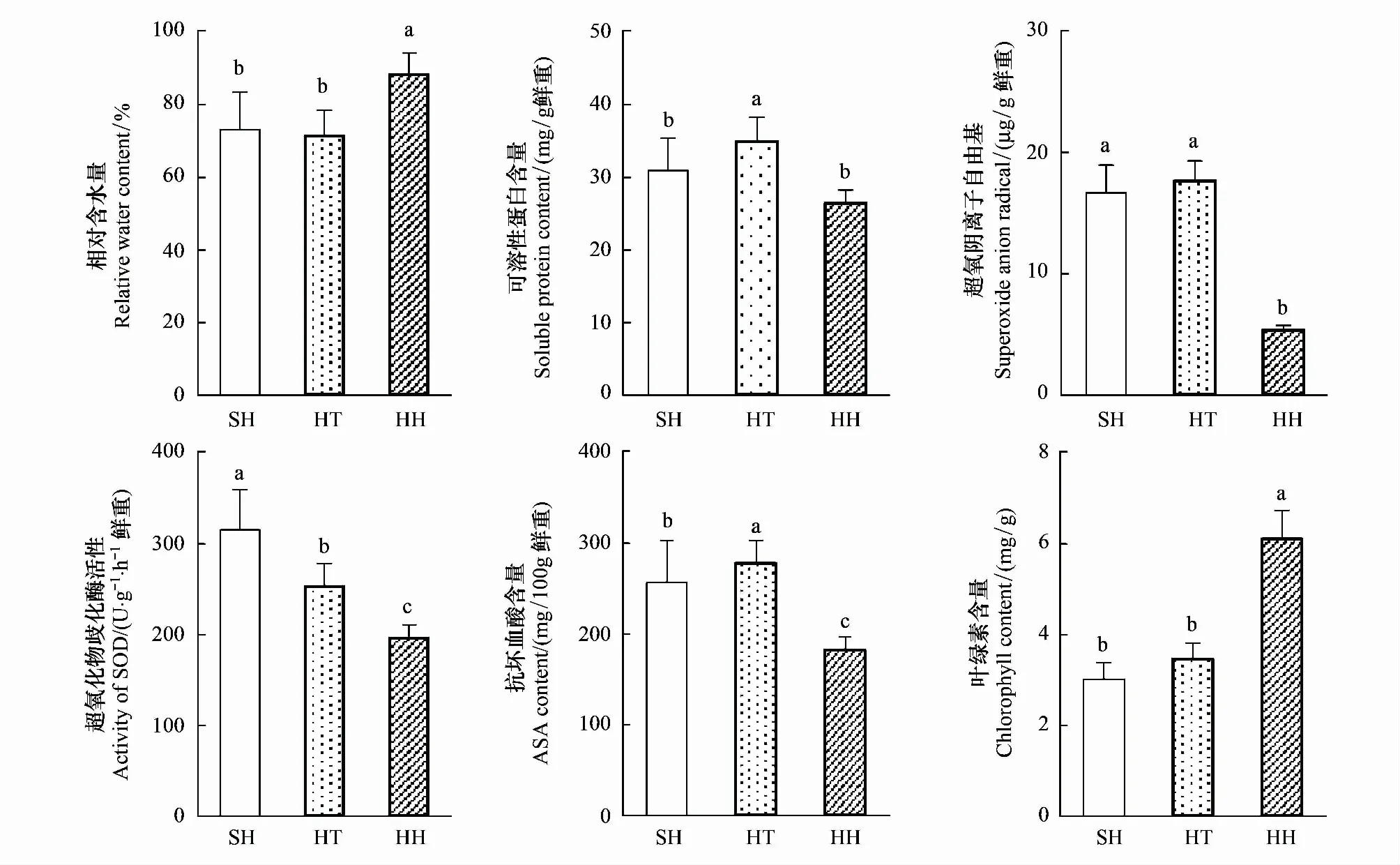
圖5 不同立地環境下豬毛蒿葉片生理指標變化特征Fig.5 Physiolgical characteristics of A. scoparia at different sites
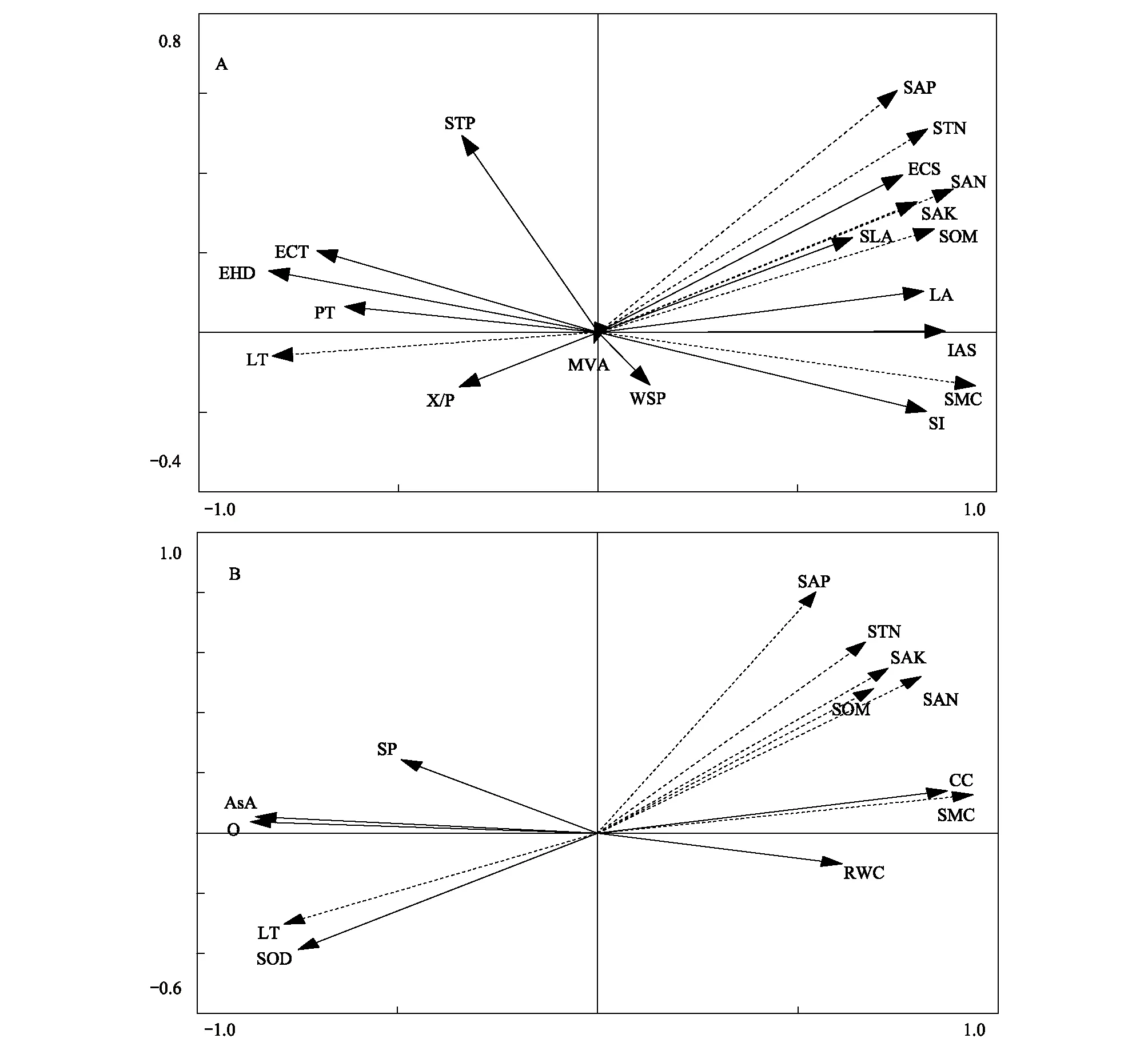
圖6 豬毛蒿葉片形態解剖指標(A)和生理特征(B)與立地環境因子相關性Fig.6 The correlation between foliar morphological, anatomical index (A), physiolgical characteristics (B) and environmental factors of A. scoparia實線表示豬毛蒿相關測量指標, 虛線表示環境因;LT:光照強度 light intensity;SMC:土壤水分 soil moisturecontent;SOM:土壤有機質 soil organic matter;STN:土壤全氮 soil total N;SAN:土壤有效氮 soil available N; SAP:土壤有效磷 soil available P;SAK:土壤有效鉀 soil available K;RWC:相對含水量 relative water content;SP:可溶性蛋白 soluble protein;O:超氧陰離子自由基 superoxide anion radical;SOD:超氧化物歧化酶 superoxide dismutase;AsA:抗壞血酸 ascorbic acid;CC:葉綠素含量 chlorophyll content
2.4豬毛蒿葉片形態解剖和生理指標的相互關系分析
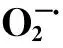
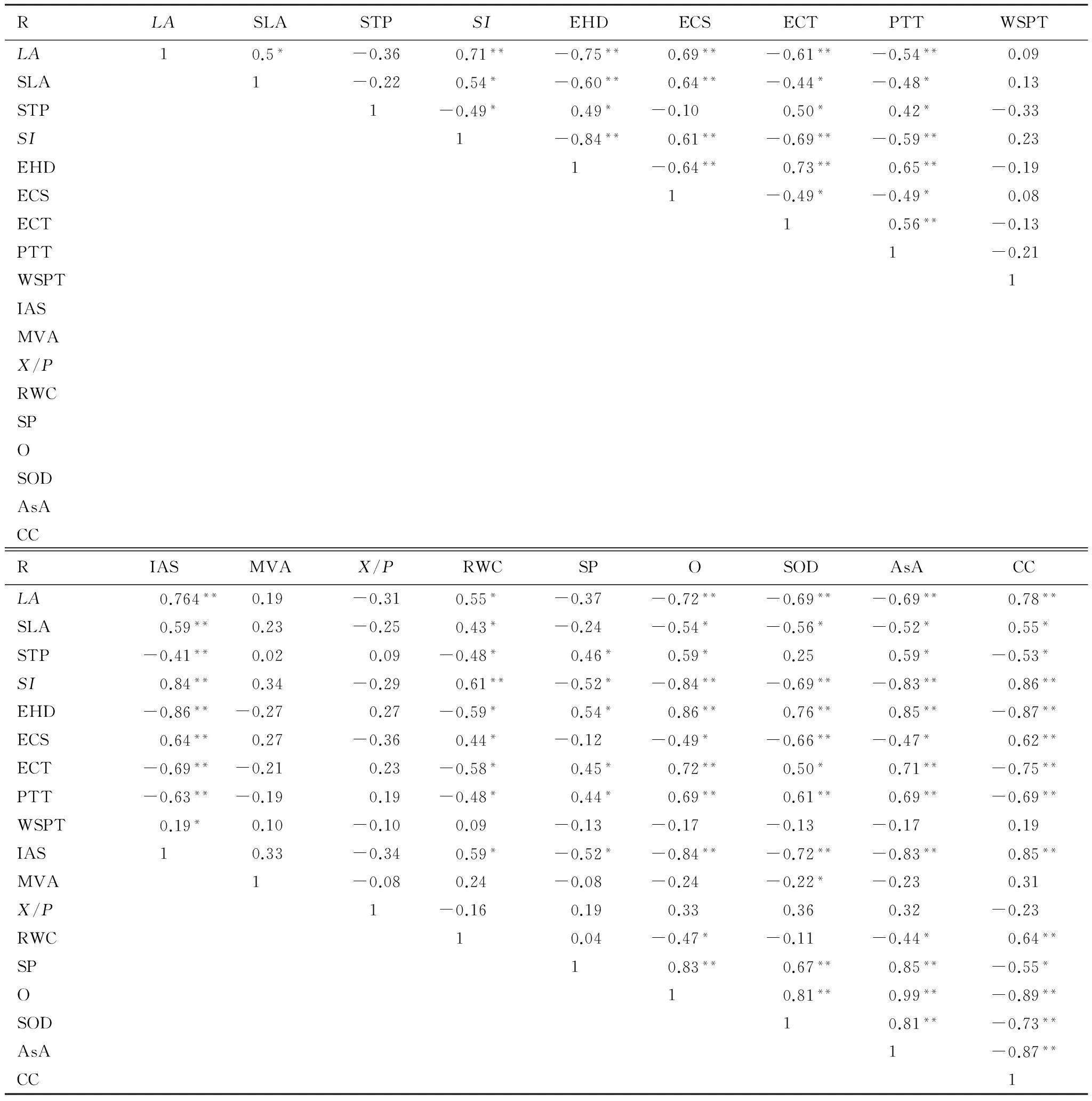
表3 豬毛蒿葉片形態解剖特征及生理指標相關性
**表示極顯著水平P<0.01; *表示顯著水平P<0.05;LA:葉面積 leaf area;SLA:比葉面積 specific leaf area;STP:氣孔可塑性 stomatal plasticity;SI:氣孔指數 stomatal index;STD:氣孔密度 stomatal density;EHD:表皮毛密度 epidermal hair density;ECS:表皮細胞大小 epidermal cells size;ECT:表皮角質層厚度 epidermal cuticular thickness;PTT:柵欄組織厚 palisade thickness;WSPT:貯水組織厚 water storage parenchyma thickness;IAS:細胞間隙率 intercellular air space;MVA:主維管束面積 main vascular bundles;X/P:木質部韌皮部面積比 area ratio of xylem to phloem;RWC:相對含水量 relative water content;SP:可溶性蛋白 soluble protein;O:超氧陰離子自由基 superoxide anion radical;SOD:超氧化物歧化酶 superoxide dismutase;AsA:抗壞血酸 ascorbic acid;CC:葉綠素含量 chlorophyll content
3討論
3.1豬毛蒿葉片形態解剖特征對立地環境的響應
陜北黃土丘陵溝壑區土地撂荒后,其生態系統的明顯特征是土壤貧瘠,生物多樣性降低,小氣候的各項指標變幅大[21],加之地區嚴重的土壤侵蝕產生的土壤擾動也直接干擾著植物的發育。黃土高原先鋒物種豬毛蒿進化出了適應該環境的葉片特征,如葉片針形化、具表皮毛、環柵型葉肉、貯水組織發達、維管束退化、具有裂生分泌腔等,豬毛蒿葉片的這些形態和解剖特征都使得豬毛蒿對撂荒后土壤貧瘠但含水量相對較大[22]、強光照的立地環境下具有較高的適應特征。不同立地環境下,在土壤、空氣濕度相對干燥和強光生境的陽峁坡與峁頂,豬毛蒿具有較小的葉面積及比葉面積、排列緊密柵欄組織、發達的貯水組織和致密的表皮毛,以利于其減小光照傷害、維持水分平衡和提高植物光合效率,這與前人研究不同環境下植物葉片形態結構變化趨勢相似[23-24]。但黃土高原豬毛蒿在其適應環境過程中也有獨特的特征,如豬毛蒿有較小的SLA值,同Garnier[25]研究1年生植物SLA顯著大于多年生植物的結果相反,推測可能原因是豬毛蒿葉片特化為針形,其單個葉片面積減小而引起SLA值減小,而這種變化使得葉片對貧瘠的土壤環境中有限資源的利用能力提高[26],有效確保了豬毛蒿在群落演替初期具有寬生態位[27]。陽峁坡和峁頂環境下豬毛蒿氣孔指數較小且氣孔可塑性在3種立地環境下差異不明顯,與前人研究脅迫環境下植物氣孔指數增大的結果相反[28],推測原因是由于豬毛蒿葉片維管組織退化,葉片肉質化使得貯水組織發達,氣孔調節水分平衡的作用相應降低,同時發達的貯水組織可有效耐受演替初期較強的土壤侵蝕對其根系的擾動破壞而造成植物的植物水分吸收困難問題。
3.2豬毛蒿葉片生理特征對立地環境的響應
3.3豬毛蒿葉片形態解剖、生理指標與環境因子的相互關系
豬毛蒿葉片形態與生理指標相關性表明,能增加豬毛蒿抗逆性的形態解剖指標,如小的葉面積、致密的表皮毛、厚的角質層與柵欄組織、小的細胞間隙,與抗逆生理指標低葉片相對含水量、高SOD與AsA含量呈顯著的正相關性。說明豬毛蒿在適應環境的過程中不但通過表型可塑性調節減小個體傷害、提高水分利用效率,同時通過生理可塑性減小組織代謝活動、增加抗逆物質積累來提高自身抗逆性,其形態解剖與生理可塑性相互影響、共同調節來適應立地環境。
豬毛蒿葉片肉質化C3植物出現類似CAM 植物的特性,其維管組織輸送水分的功能大部分為其貯水組織替代[34],因此RDA分析顯示出豬毛蒿體內運送水分的維管組織特征、調節水分平衡的氣孔特性和滲透調節物質可溶性蛋白都與立地環境相關性較小。除水分因子外,豬毛蒿葉片形態、解剖和生理的可塑性對立地有機質含量和光照強度也較為敏感,如陽峁坡其SLA可迅速減小以提高該立地土壤環境中有限資源的利用效率;光照較強時體內抗逆物質SOD和AsA的活性增加效降低光照脅迫。以上葉片形態和生理的可塑性使豬毛蒿對撂荒初期光照強度大、土壤貧瘠但土層干化現象尚未出現[4,22]的立地環境有很好的適應性,加之其r-生活史策略可使其種群在撂荒后裸地快速拓展[35],使其成為黃土高原植被自然演替過程中的先鋒物種。
參考文獻(References):
[1]杜峰, 山侖, 陳小燕, 梁宗鎖. 陜北黃土丘陵區撂荒演替研究-撂荒演替序列. 草地學報, 2005, 13(4): 328- 333.
[2]焦菊英, 張振國, 賈燕鋒, 王寧, 白文娟. 陜北丘陵溝壑區撂荒地自然恢復植被的組成結構與數量分類. 生態學報, 2008, 28(7): 2981- 2997.
[3]鄒厚遠, 焦菊英. 黃土丘陵區生態修復地不同抗侵蝕植物的消長變化過程. 水土保持通報, 2009, 29(4): 235- 240.
[4]郝文芳, 梁宗鎖, 陳存根, 唐龍. 黃土丘陵溝壑區棄耕地群落演替與土壤性質演變研究. 中國農學通報, 2005, 21(8): 226- 231.
[5]Catoni R, Gratani L, Varone L. Physiological, morphological and anatomical trait variations between winter and summer leaves of Cistus species. Flora-Morphology, Distribution, Functional Ecology of Plants, 2012, 207(6): 442- 449.
[6]Pintó-Marijuan M, Munné-Bosch S. Ecophysiology of invasive plants: osmotic adjustment and antioxidants. Trends in Plant Science, 2013, 18(12): 660- 669.
[7]Nicotra A B, Atkin O K, Bonser S P, Davidson A M, Finnegan E J, Mathesius U, Poot P, Purugganan M D, Richards C L, Valladares F, van Kleunen M. Plant phenotypic plasticity in a changing climate. Trends in Plant Science, 2010, 15(12): 684- 692.
[8]Miner B G, Sultan SE, Morgan S G, Padilla D K, Relyea R A. Ecological consequences of phenotypic plasticity. Trends in Ecology & Evolution, 2005, 20(12): 685- 692.
[9]Bochet E, García-Fayos P, Poesen J. Topographic thresholds for plant colonization on semi-arid eroded slopes. Earth Surface Processes and Landforms, 2009, 34(13): 1758- 1771.
[10]焦菊英, 王寧, 杜華棟, 王東麗. 土壤侵蝕對植被發育演替的干擾與植物的抗侵蝕特性研究進展. 草業學報, 2012, 21(5): 311- 318.
[11]王勇, 梁宗鎖, 龔春梅, 韓蕊蓮, 于靖. 干旱脅迫對黃土高原4種蒿屬植物葉形態解剖學特征的影響. 生態學報, 2014, 34(16): 4535- 4548.
[12]杜峰, 梁宗鎖, 山侖, 陳小燕. 黃土丘陵區不同立地條件下豬毛蒿種內、種間競爭. 植物生態學報, 2006, 30(4): 601- 609.
[13]胡澍, 焦菊英, 杜華棟, 苗芳. 黃土丘陵溝壑區不同立地環境下植物的抗氧化特性. 草業學報, 2014, 23(5): 1- 12.
[14]Nardini A, Peda G, La Rocca N. Trade-offs between leaf hydraulic capacity and drought vulnerability: morpho-anatomical bases, carbon costs and ecological consequences. New Phytologist, 2012, 196(3): 788- 798.
[15]Yang S J, Sun M, Zhang Y J, Cochard H, Cao K F. Strong leaf morphological, anatomical, and physiological responses of a subtropical woody bamboo (Sinarundinarianitida) to contrasting light environments. Plant Ecology, 2014, 215(1): 97- 109.
[16]Royer D L. Stomatal density and stomatal index as indicators of paleoatmospheric CO2concentration. Review of Palaeobotany and Palynology, 2001, 114(1/2): 1- 28.
[17]Valladares F, Wright S J, Lasso E, Kitajima K, Pearcy R W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology, 2000, 81(7): 1925- 1936.
[18]高俊鳳. 植物生理學實驗技術. 西安: 世界圖書出版公司, 2000.
[19]孫群, 胡景江. 植物生理學研究技術. 楊凌: 西北農林科技大學出版社, 2006.
[20]張憲政. 植物葉綠素含量測定—丙酮乙醇混合液法. 遼寧農業科學, 1986, (3): 26- 28.
[21]張成娥, 陳小利. 黃土丘陵區不同撂荒年限自然恢復的退化草地土壤養分及酶活性特征. 草地學報, 1997, 5(3): 195- 200.
[22]杜峰, 山侖, 梁宗鎖, 譚勇. 陜北黃土丘陵區撂荒演替過程中的土壤水分效應. 自然資源學報, 2005, 20(5): 669- 678.
[23]Ahmad P, Prasad M N V. Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability. New York: Springer, 2012: 21- 39.
[24]李芳蘭, 包維楷. 植物葉片形態解剖結構對環境變化的響應與適應. 植物學通報, 2005, 22(增刊): 118- 127.
[25]Garnier E. Growth analysis of congeneric annual and perennial grass species. Journal of Ecology, 1992, 80(4): 665- 675.
[26]Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335- 380.
[27]王寧, 賈燕鋒, 李靖, 焦菊英. 黃土丘陵溝壑區退耕地自然恢復植被主要物種生態位特征. 水土保持通報, 2007, 27(6): 34- 40.
[28]楊惠敏, 王根軒. 干旱和CO2濃度升高對干旱區春小麥氣孔密度及分布的影響. 植物生態學報, 2001, 25(3): 312- 316.
[29]Martínez J P, Kinet J M, Bajji M, Lutts S. NaCl alleviates polyethylene glycol-induced water stress in the halophyte speciesAtriplexhalimusL.. Journal of Experimental Botany, 2005, 56(419): 2421- 2431.
[30]Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science, 2002, 7(9): 405- 410.
[31]武維華. 植物生理學. 北京: 科學出版社, 2003.
[32]付曉玥. 阿拉善荒漠植物葉片性狀研宄[D]. 呼和浩特: 內蒙古大學, 2012.
[33]Gibson A C. Structure-Function Relations of Warm Desert Plants. Berlin: Springer, 1996.
[34]Fondom N Y, Castro-Nava, S, Huerta A J. Seasonal variation in photosynthesis and diel carbon balance under natural conditions in two Peperomia species that differ with respect to leaf anatomy. The Journal of the Torrey Botanical Society, 2009, 136(1): 57- 69.
[35]杜華棟. 黃土丘陵溝壑區優勢植物對不同侵蝕環境的適應研究—從群落、個體、組織到生理[D]. 北京: 中國科學院大學, 2013.
Adaptability of foliar morphological, anatomical, and physiological characteristics of the pioneer speciesArtemisiascopariagrowing in a hilly-gully Loess Region at different slope sites
DU Huadong1,2, JIAO Juying2,*, KOU Meng2, MIAO Fang3
1CollegeofGeology&Environment,Xi′anUniversityofScienceandTechnology,Xi′an710054,China2InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistryofWaterResources,Yangling712100,China3CollegeofLifeScience,NorthwestAgriculture&ForestryUniversity,Yangling712100,China
Abstract:Soil erosion by water is considered to be a dominant erosion process in a hilly-gully Loess Plateau, and leads to land degradation and desertification. Although many researchers have investigated the dynamics of plant community traits and soil properties caused by erosion, the effects of soil property variation on vegetation succession, especially in terms of ecophysiology, on abandoned cultivated land in a hilly-gully Loess Plateau region have received little attention to date. This study investigated Artemisia scoparia, which is one of the dominant species in early abandoned field communities in hilly-gully Loess Plateau. We tried to find the environmental acclimation and succession niche changes for this species. The foliar morphological, anatomical, and physiological characteristics of A. scoparia at three different sites (namely sunny hilly slope, SH; hilltop, HT; shady hilly slope, HH) were investigated. Relationships between foliar plasticity and environmental factors were also examined. The results demonstrate that (1) A. scoparia has special morphological and anatomical characteristics, such as needle-shaped leaves, trichomes, a ring palisade, water storage parenchyma specialized from spongy tissue, and a secretory cavity, that enable it to adapt to semi-arid environments. When the ability of vascular tissue to transport water decreased, C3 plant leaves exhibit characteristics similar to CAM plants. (2) On the SH and HT sites, which are characterized by dry soil, low air humidity, and strong light intensity, A. scoparia had a smaller leaf area, thicker palisade, denser trichomes, and tighter intercellular air spaces than of those growing on the HH site. Furthermore, obvious differences among site conditions can be observed. However, the redundancy analysis (RDA) results revealed that vascular tissue traits, main vascular bundles, and area ratio of xylem to phloem, have a weak correlation with environmental factors. (3) A. scoparia had a low relative water content (RWC), less chlorophyll, and a high superoxide anion radical content on the SH and HT sites. In contrast, superoxide dismutase (SOD) and ascorbic acid (AsA) increased at these sites to clear the reactive oxygen produced by plants. Soluble protein, which shows the osmotic adjustment ability of a plant, had a weaker relationship with environmental factors. (4) Foliar plasticity had a strong relationship with environmental factors, especially with light intensity, soil moisture, and soil organic matter content. These results implied that A. scoparia had strong adaptability to early abandoned sites in a hilly-gully Loess Region that is subject to high light intensities and soil impoverishment without desiccation. (5) Based on the correlations among foliar morphological, anatomical, and physiological characteristics of A. scoparia, the studied parameters, except for vascular tissue traits and soluble protein, were found to be closely related to each other, especially epidermal hair density, stomatal index, and intercellular air spaces. All the above results suggest that A. scoparia is able to adapt to the microclimate and soil conditions present in early abandoned fields due to its foliar anatomical and physiological characteristics. These properties allow A. scoparia to improve its ability to use limited nutrient resources in poor soil environments and to reduce abiotic stresses (including water, light, nutrient, and erosion interference stresses). This means that A. scoparia is able to attain and dominate an extremely wide ecological distribution on early abandoned cultivated land in hilly-gully Loess Plateau regions.
Key Words:Loess Plateau; ecological adaptation; leaf morphology and anatomy; stress-resistance physiology; Artemisia scoparia
基金項目:國家自然科學基金項目(41371280, 41030532);陜西省教育廳科學研究基金項目(14JK1481)
收稿日期:2014- 08- 19; 網絡出版日期:2015- 09- 28
*通訊作者
Corresponding author.E-mail: jyjiao@ms.iswc.ac.cn
DOI:10.5846/stxb201408191642
杜華棟, 焦菊英, 寇萌, 苗芳.黃土高原先鋒種豬毛蒿葉片形態解剖與生理特征對立地的適應性.生態學報,2016,36(10):2914- 2925.
Du H D, Jiao J Y, Kou M, Miao F.Adaptability of foliar morphological, anatomical, and physiological characteristics of the pioneer speciesArtemisiascopariagrowing in a hilly-gully Loess Region at different slope sites.Acta Ecologica Sinica,2016,36(10):2914- 2925.
