合浦珠母貝3個養殖群體雙列雜交F1代的生長和遺傳分析
2016-07-06 10:29:49黃桂菊陳飛飛劉寶鎖范嗣剛刑孔武喻達輝中國水產科學研究院南海水產研究所農業部南海漁業資源開發利用重點實驗室南海資源開發與保護協同創新中心廣東廣州50300上海海洋大學水產與生命學院上海20306
海洋科學 2016年2期
關鍵詞:生長
黃桂菊, 陳飛飛, 2, 劉寶鎖, 范嗣剛, 刑孔武, 喻達輝(. 中國水產科學研究院 南海水產研究所 農業部南海漁業資源開發利用重點實驗室, 南海資源開發與保護協同創新中心, 廣東 廣州 50300; 2. 上海海洋大學 水產與生命學院, 上海 20306)
?
合浦珠母貝3個養殖群體雙列雜交F1代的生長和遺傳分析
黃桂菊1, 陳飛飛1, 2, 劉寶鎖1, 范嗣剛1, 刑孔武1, 喻達輝1
(1. 中國水產科學研究院 南海水產研究所 農業部南海漁業資源開發利用重點實驗室, 南海資源開發與保護協同創新中心, 廣東 廣州 510300; 2. 上海海洋大學 水產與生命學院, 上海 201306)
為開展合浦珠母貝(Pinctada fucata)遺傳改良和培育優質新品種, 作者利用合浦珠母貝北海養殖群體(B)、徐聞養殖群體(X)和三亞養殖群體(S)進行雙列雜交, 獲得3個種群內自繁群體及6個種群間雜交群體。對8月齡F1代的殼長、殼高、殼寬和體質量4個生長性狀進行了測定, 計算了各組合的一般配合力、特殊配合力及雜種優勢。結果表明, 雜交組合的各性狀值普遍高于自繁群體, 其中S(♀)×B(♂)和S(♀)×X(♂)雜交組合的各性狀值極顯著高于其他組合。三亞群體各性狀的一般配合力最大, S(♀)×X(♂)組合各性狀特殊配合力最大, 其次是S(♀)×B(♂)組合。雜交子一代4個生長指標均表現出一定程度的雜種優勢(0.24%~52.62%)。不同雜交組合間和不同性狀間的雜種優勢存在差異, 其中, S(♀)×B(♂)雜交組合雜種優勢明顯, 其反交組合B(♀)×S(♂)雜交優勢程度不高, 且在殼寬上表現出較低的雜種優勢率(0.24%); S(♀)×X(♂)雜交組合雜種優勢率比反交組合X(♀)×S(♂)高。體質量的平均雜種優勢率均高于其他3個性狀。該研究的結果表明雜交群體性狀的改進為進一步選育奠定了基礎。
合浦珠母貝(Pinctada fucata); 雙列雜交群體; 生長; 配合力; 雜交優勢
[Foundation: the Earmarked Fund for China Agriculture Research System(CARS-48); Guangdong Provincial Marine Fisheries Science & Technology Promotion Special Projects(A201301A02, A201301A08, Z2014003)]
中國自1965年合浦珠母貝(Pinctada fucata)人工育苗成功以來, 海水珍珠養殖經歷了半個世紀的發展, 由于近親繁殖嚴重, 導致養殖性狀退化、貝體變小、死亡率高、病害增多, 同時生產珍珠顆粒小、珠層薄等種質退化現象。因此, 解決這個問題的關鍵是開展合浦珠母貝的遺傳改良和新品種培育。
目前對合浦珠母貝的研究已開展了殼型、殼色、殼厚、生長速度的選育[1-3]、不同地理群體間的雜交育種[4]和家系選擇育種[5]、貝殼珍珠質顏色分析[6]、加性遺傳效應估計[7]、遺傳多樣性與性狀相關性分析[8]、生長性狀和閉殼肌拉力的比較分析[9]、中藥對插核育珠的影響[10]、卵子電擊脈沖參數的初步研究[11]、生長性狀的相關分析[12]等研究。黎火金等[13]通過測量以海南三亞和廣東徐聞2 個地理群體合浦珠母貝為親本構建的33個全同胞家系的殼長、殼高、殼寬和體質量4 個生長性狀對其進行遺傳參數分析, 發現合浦珠母貝4 個生長性狀均屬中等遺傳力, 可以用選擇育種進行遺傳改良。Wada等[2-3]對合浦珠母貝的殼型參數的遺傳力進行了研究, 結果表明合浦珠母貝的殼寬、殼凸度遺傳力較大, 殼厚則次之; 不同地理種群和不同品系的合浦珠母貝雜交和回交, 雜交后代在成活率和殼寬上均表現出明顯的雜種優勢。王愛民等[4, 14]利用廣西北海野生種的合浦珠母(BW)、廣東大亞灣野生合浦珠母貝(DW)和海南三亞野生合浦珠母貝(SW)開展了不同地理種群內自繁和種群間雜交的工作, 運用方差分析對6個子一代群體的殼長、殼寬、殼厚、總體質量和殼質量5個主要性狀進行了比較, 發現3個雜交群體未表現出雜種優勢, 但獲得的6個群體子一代的變異性增加, 為進一步選育奠定了基礎。谷龍春等[15]用廣西北海(B)、廣東徐聞(X)和海南三亞(S)親貝構建了90個雙列雜交家系。分析表明, 北海群體的殼長、殼高的一般配合力最大; 三亞-北海殼長、殼高的特殊配合力最大。
雜種優勢是指雜交子代在生長、成活、繁殖能力或生產性能等方面均優于雙親均值的現象, 雜交育種是培育新品種的重要途徑之一。目前, 在貽貝(Mytilus edulis)[16]、櫛孔扇貝(Chlamys farreri)[17]、太平洋牡蠣(Crassostrea gigas)[18]、海膽(Echinometra spp.)[19-20]和其他蝦類[21-23]的雜交育種已取得巨大進展。作者開展了合浦珠母貝3個不同地理養殖群體的雜交試驗, 在相同養殖管理條件下測定合浦珠母貝不同群體自繁和雜交子代的生長性狀差異, 計算各組合的一般配合力、特殊配合力及雜種優勢, 為利用雜種優勢培育合浦珠母貝新品種奠定了基礎。
1 材料和方法
1.1 親本來源
合浦珠母貝廣西北海群體(用B表示)采自廣西北海一養殖場, 廣東徐聞群體(X)采自廣東徐聞一養殖場, 海南三亞群體(S)采自海南省三亞陵水新村養殖場。經海上暫養, 使其達到同步性成熟, 選擇發育良好、性腺成熟的個體作為親貝。
1.2 雙列雜交群體的構建
2011年2月17日, 開展了3個不同地理養殖群體內自繁和群體間雜交試驗, 獲得3個自繁群體(BB、XX和SS)和6個正反交群體(BX、XB、SB、BS、SX、XS)(表1)。3個自繁群體按照雌雄比例為10∶6的方式通過陰干刺激后自然排放受精而獲得, 6個雜交群體按照雌雄比例為10∶6的方式采用人工授精而得, 人工授精過程中利用0.6‰的氨海水刺激精卵。待受精完成后, 轉入10 m3的水池內培育。培育用海水為過濾后的自然海水。
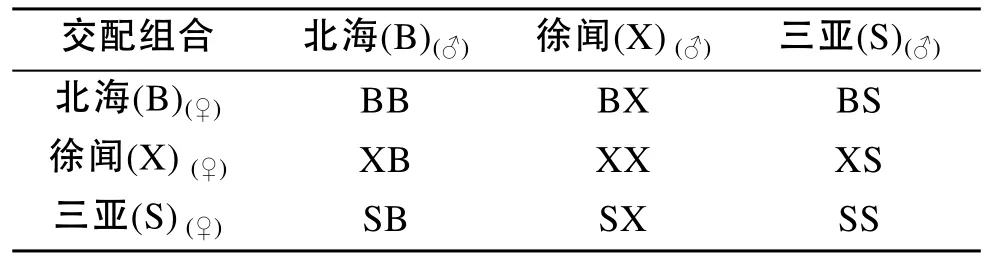
表1 不同地理來源群體的交配設計Tab. 1 Mating design with parental populations from Beihai (B), Xuwen (X), and Sanya (S)
1.3 苗種培育與養殖
9個群體的培育采用常規方法進行[1], 早期以湛江等鞭金藻(Isochrysis zhanjiangensis)為主要餌料,適當混合投喂亞心形扁藻(Platymonas subcordiformis)和酵母等, 后期以亞心形扁藻為主要餌料。每天換水1/3, 連續充氣培養。
2011年4月23日幼苗達到3.0 mm左右后將幼苗移至中國水產科學研究院南海水產研究所陵水新村珍珠貝養殖基地進行固定排架吊養。分別用80、60、40、20目貝苗籠, 以及網目為1和3 cm單圈籠養殖, 先用網目密的80目貝苗籠, 隨著貝的生長逐漸換用網眼大的籠。每個群體的養殖密度和管理方法相同[24]。
1.4 數據測定
2011年10月從每個群體中隨機抽樣, 用于生長測量。測量前先清貝, 然后每個群體隨機抽取100個樣本測量其殼長、殼高、殼寬和體質量4個數量性狀。殼長、殼高和殼寬用游標卡尺行測量, 精確到0.01cm。體質量用電子天平稱量, 精確到0.01 g。
1.5 數據處理
各數據用Excel和SPSS16.0軟件分析和處理,差異顯著性分析采用最小顯著差法。一般配合力、特殊配合力和雜種優勢的計算按文獻方法[25-26]。
一般配合力估算應用公式為:
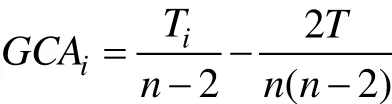
式中: Ti為某種群為共同親本的所有雜交組合的總和, T為所有雜交組合的總和,, n為種群數。特殊配合力估算應用公式為:
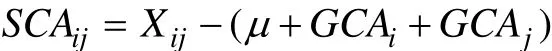
式中: Xij為兩個種群特定組合實測值, μ為所有雜交組合的平均值, GCAi為親本種群i的一般配合力, GCAj為親本種群j的一般配合力, 雜交子代的雜種優勢率H(%)計算公式為:
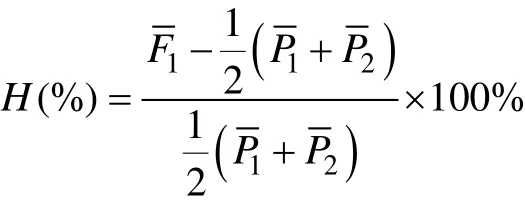
式中:F1、P1、P2分別代表雜交群體和兩個親本群體內的自繁群體的平均值, H(%)代表F1的雜種優勢率。
2 結果
2.1 9個群體各生長性狀的分布
群體BSF1、XSF1、SSF1、XXF1、BBF1的殼長主要分布在22.11~38.60 mm, 分別占87%、86%、90%、92%、74%。群體BXF1、XBF1的殼長主要分布在27.61~38.60 mm, 分別占69%、76%。群體SBF1、SXF1的殼長主要分布在33.11~44.10mm, 分別占70%、78%。9個群體的殼高、殼寬、體質量與殼長類似, 群體BSF1、XSF1、SSF1、XXF1、BBF1、BXF1和XBF1各性狀在較低范圍內分布頻率較高, 而群體SBF1、SXF1在較高范圍內分布頻率較高(圖1)[24]。
2.2 9個群體各生長性狀的比較
9個群體各組的平均殼長為26.72~37.95 mm, 平均殼高為25.89~38.34 mm, 平均殼寬為8.91~12.18 mm,平均體質量為7.39~11.65 g, 雜交組合的各性狀值普遍高于自繁群體(表2)。方差分析表明, 組間4個性狀表型值均存在極顯著差異(表3)[24]。兩兩多重比較表明, SB和SX的4個性狀值都極顯著高于其他組合, 而XS顯著高于BX和SB雜交組合, 與BS差異不顯著(表2)。此外, 除XX組合外, 自繁組合的變異系數高于雜交組合, 體質量的變異系數遠遠大于其他性狀(表2)。
部分雜交組合中正反交之間存在顯著差異(P< 0.05, 表2)。正交組合SB和SX各性狀值分別顯著高于反交組合BS和XS, 而正交組合BX各性狀值與反交組合XB不存在顯著差異。
2.3 雜交組合配合力分析
雙列雜交組合各性狀的一般配合力和特殊配合力分析結果分別見表4和表5[24]。三亞群體雌性各性狀的一般配合力遠遠高于其他群體, 其次為徐聞雌性。但雄性個體各性狀存在差異, 三亞雄性殼長和體質量一般配合力最大, 北海雄性的殼高和殼寬最大, 徐聞群體遠遠小于其他群體(表4)。S×X和S×B組合各性狀的特殊配合力遠遠高于其他組合, 其中S×X組合殼高和體質量特殊配合力最大, S×B組合殼長和殼寬的特殊配合力最大, 此外X×S組合的體質量特殊配合力也較高(表5)。
2.4 雜交子一代的雜種優勢率
各雜交組合在4個生長指標中均表現出一定的雜種優勢(0.24%~52.62%)(圖2)。各雜交組合之間殼長的雜種優勢率大小順序為SX>SB>XS>BS>BX> XB, 殼高的雜種優勢率大小順序為SX>SB>BS> XS>BX>XB, 殼寬的雜種優勢率大小順序為SB> SX>XS>XB>BX>BS, 體質量的雜種優勢率大小順序為SX>SB>XS>BX>XB>BS。
雜種優勢在不同雜交組合間和不同性狀間存在很大差異。SB和SX組合的4個性狀的雜種優勢率均高于其他雜交組合, 除BS組合外, 體質量的雜種優勢率均高于其他3個性狀的雜種優勢率(圖2)[24]。
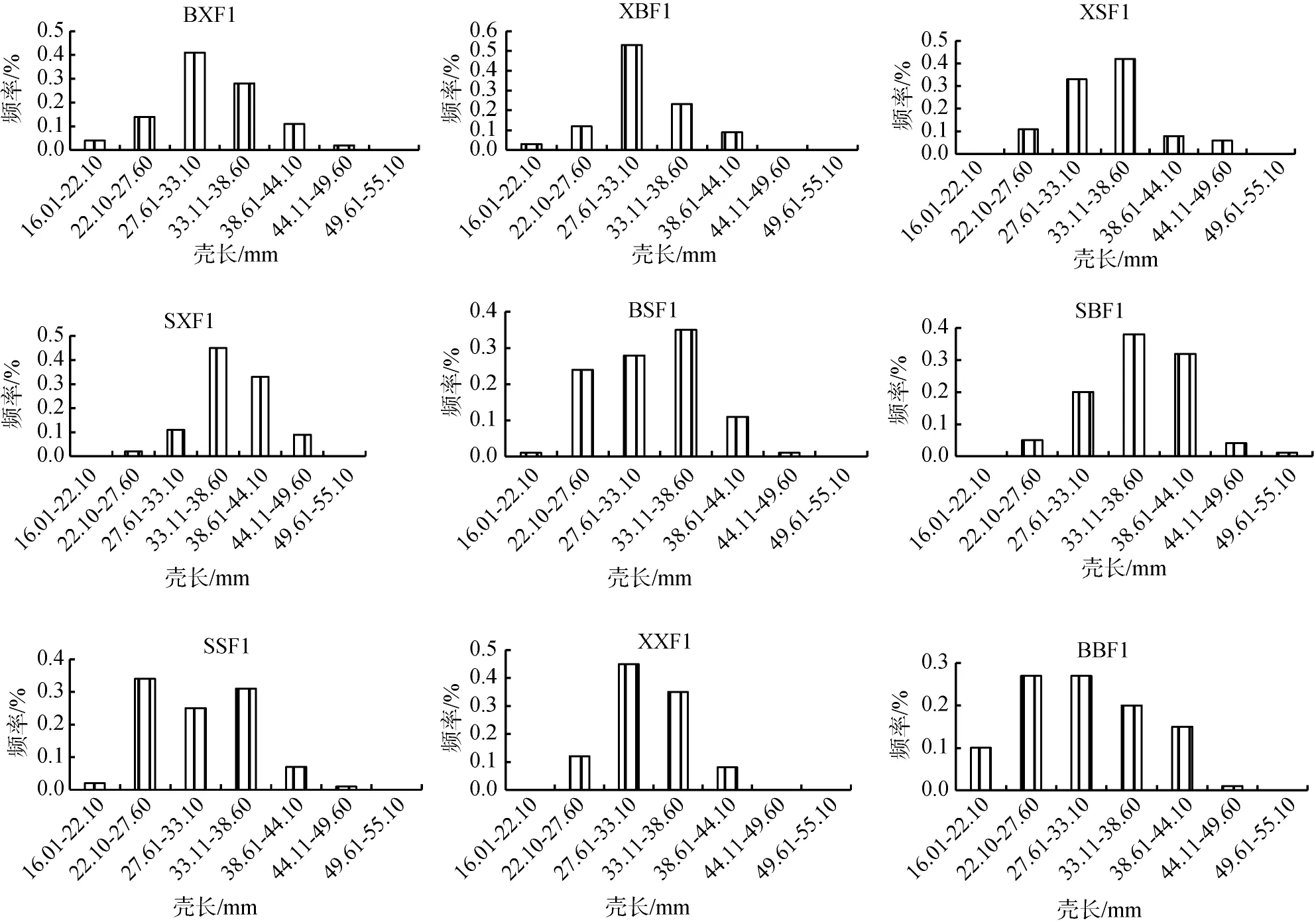
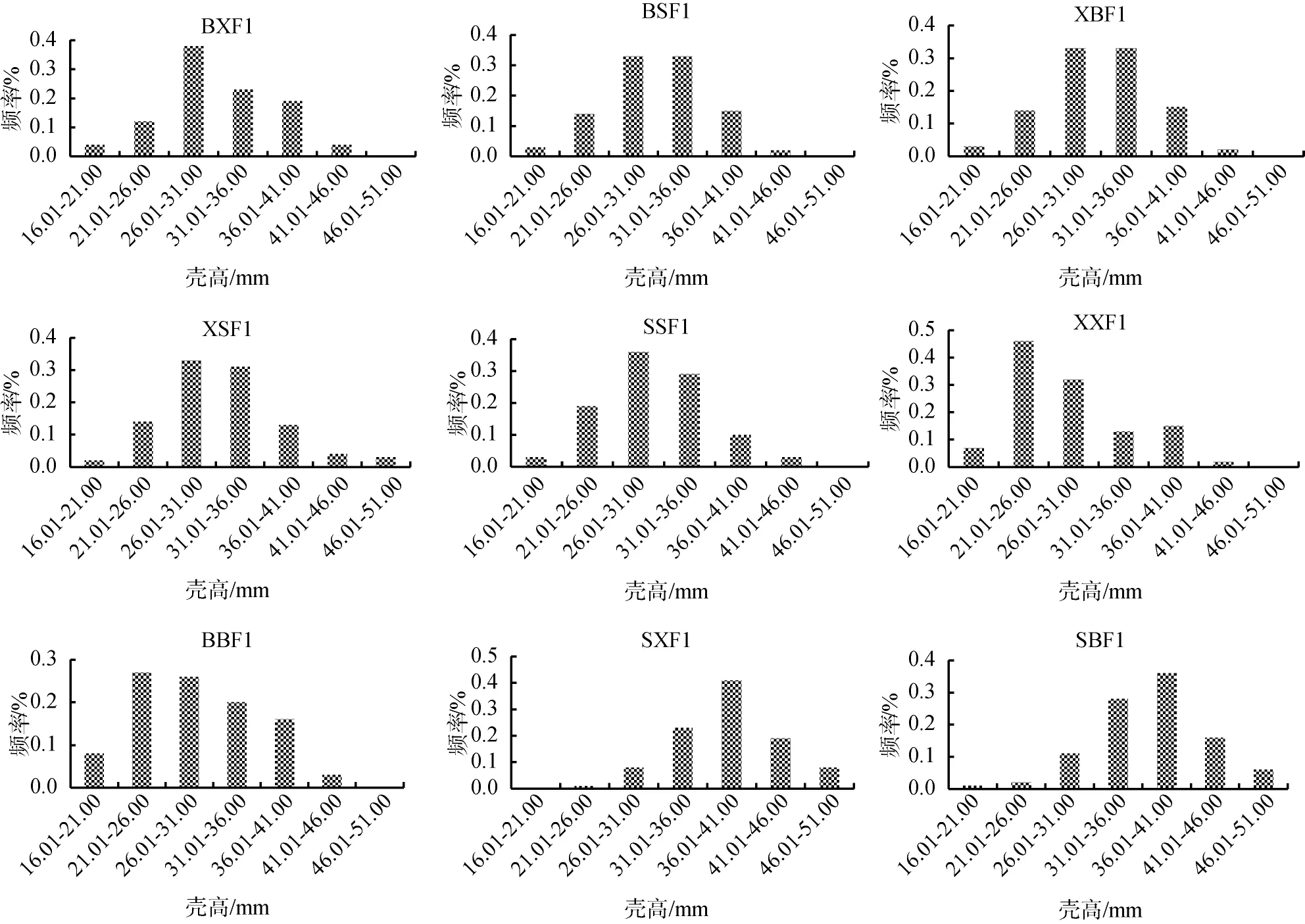
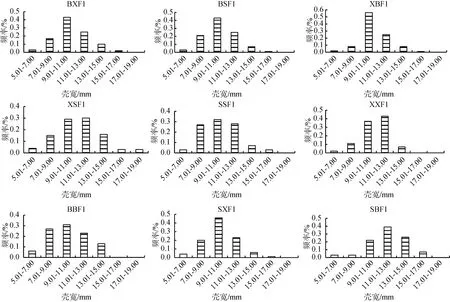
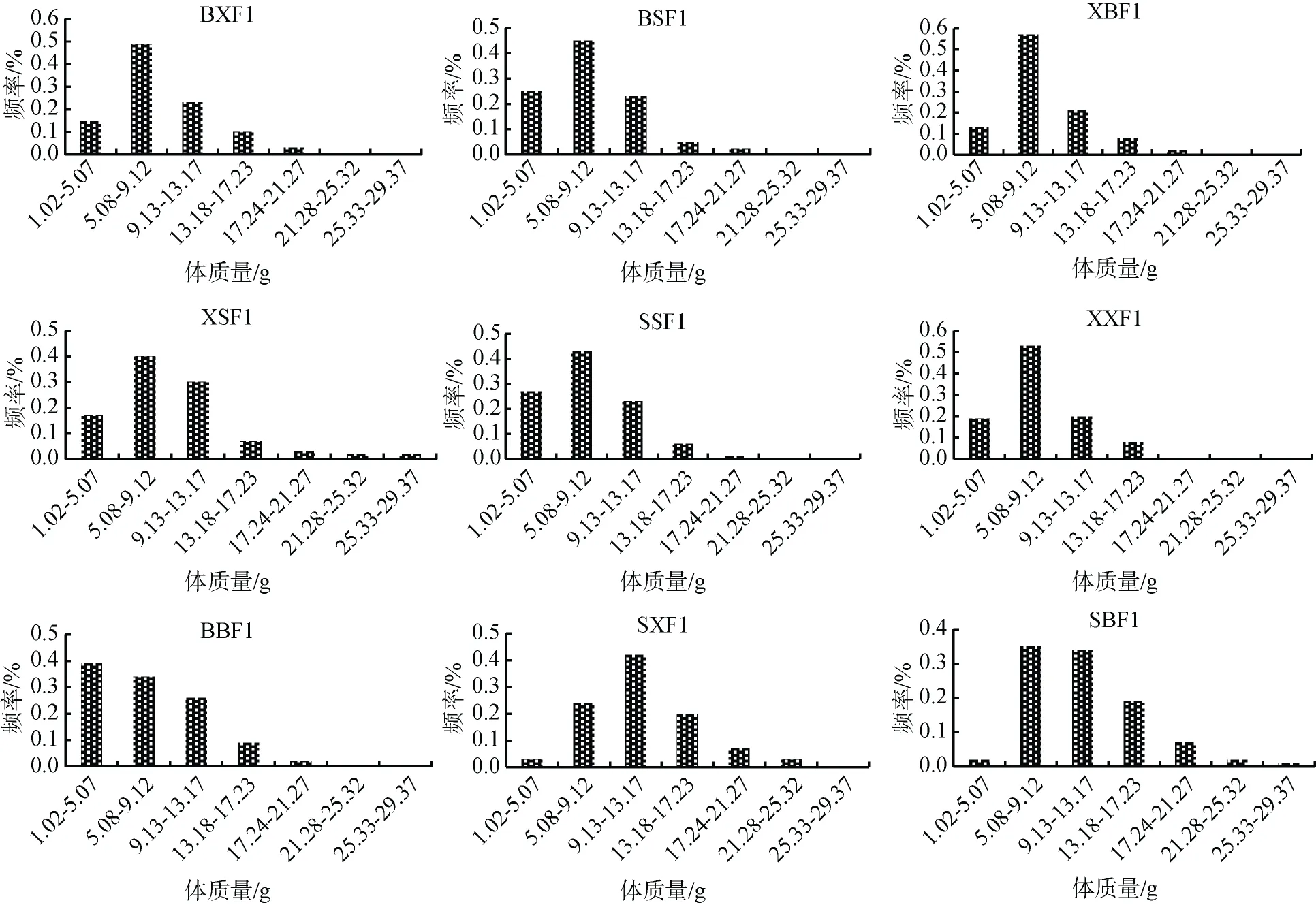
圖1 合浦珠母貝9個群體各性狀分布頻率Fig. 1 Frequency distribution of the four traits in nine crosses of P. fucata
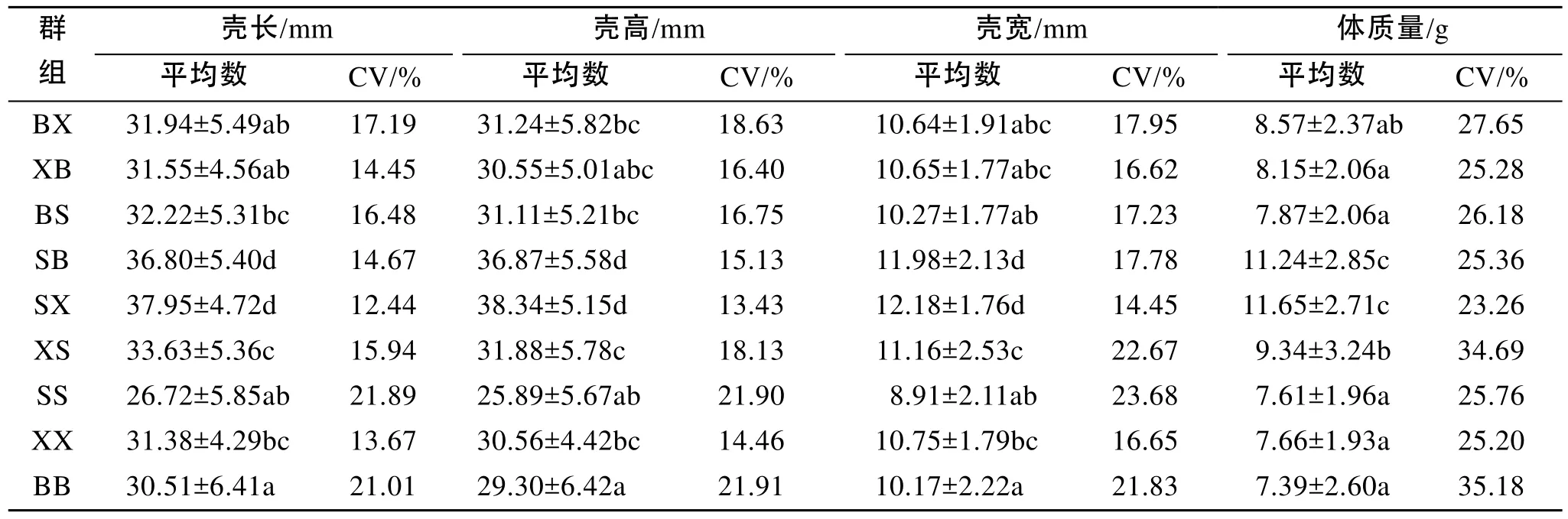
表2 合浦珠母貝雙列雜交子一代4個性狀的平均值和變異系數(CV)Tab. 2 Means of the four traits in F1groups and the coefficient of variations for P. fucata
3 討論
雙列雜交是育種中構建選育基礎群體的重要手段之一。該研究構建的9個雙列雜交群體中, 6個雜交群體各性狀的表現普遍高于3個自繁群體, 表明存在不同程度的雜交優勢。其中, S×B與S×X的生長表現顯著高于其他4個雜交群體, 說明在殼長、殼高、殼寬、體質量4個指標性狀上, 以三亞群體為母本的雜交組合要優于其他雜交組合。X×S組合雖然平均值不高, 但從各性狀的分布情況來看, 上限值卻較高。王愛民等[27]以三亞野生合浦珠母貝和大亞灣養殖貝雜交, 也發現殼高和總體質量方面表現出了明顯的提高,而且雜交子代插核育珠表現出了優良的育珠效果。在變異度方面, 各個組合不盡相同, 自繁群體BB和SS變異系數較大, 雜交群體SX的變異系數最小, 而王愛民等[4]、丁小雷等[28]研究表明不論是種群內自繁后代還是雜交子一代在各性狀指標上變異性較大。

表3 合浦珠母貝各性狀在雜交組合子一代間的方差分析Tab. 3 ANOVA of growth traits in hybrids of all combina-tions of P. fucata
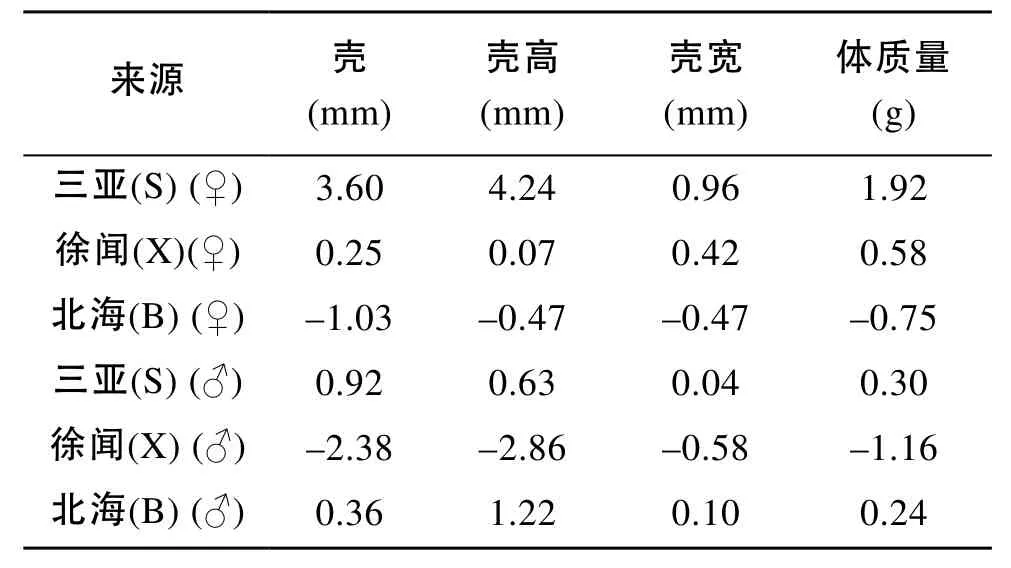
表4 合浦珠母貝雙列雜交各群體的一般配合力Tab. 4 General combining ability of diallel crosses of P. fucata
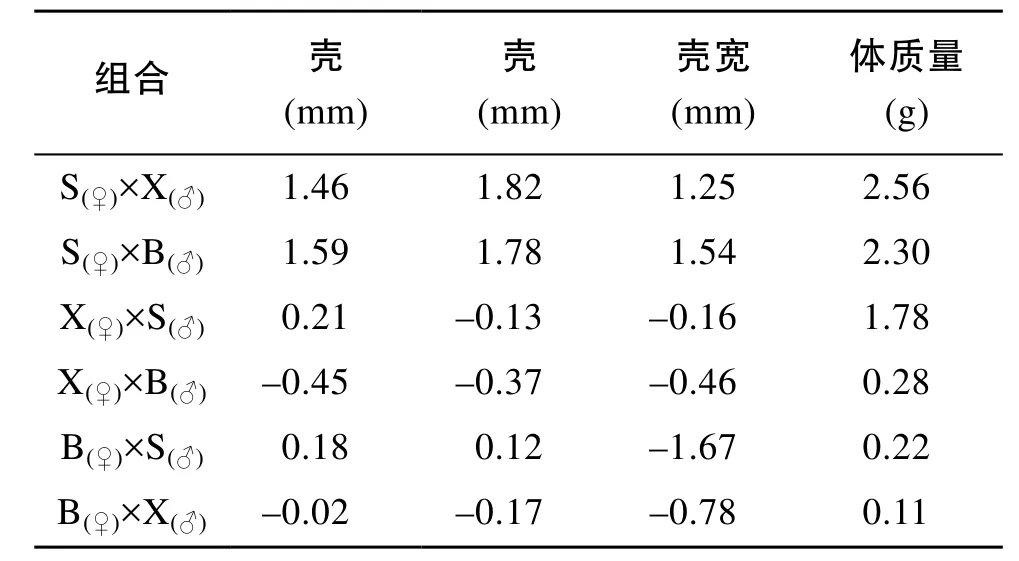
表5 合浦珠母貝雙列雜交各組合的特殊配合力Tab. 5 Specific combining ability of diallel crosses for P. fucata
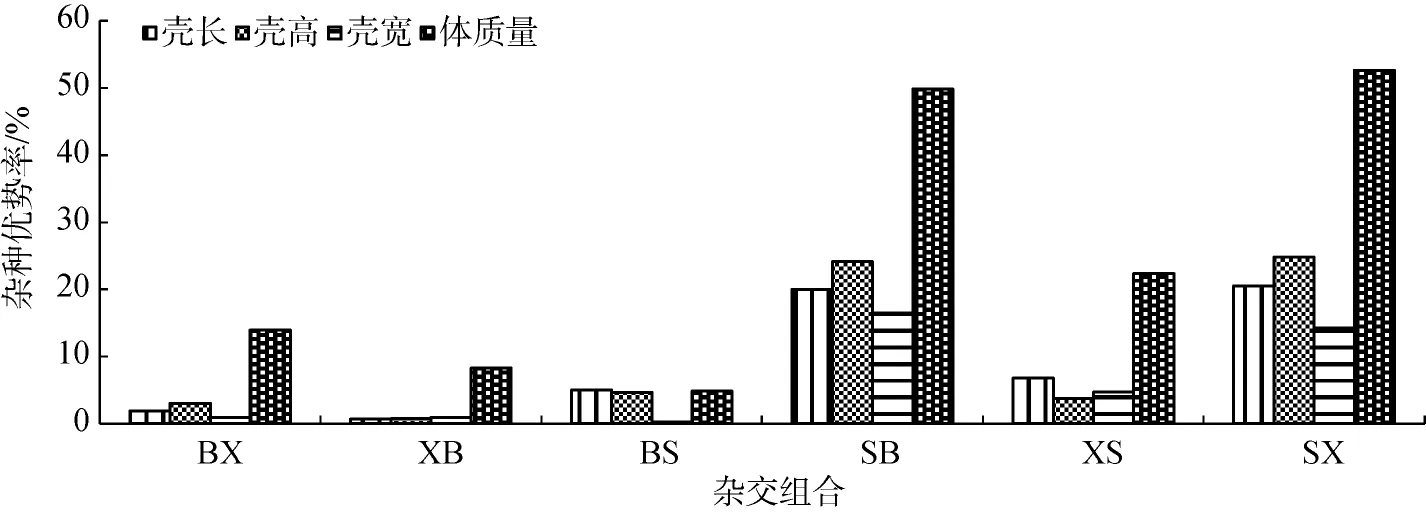
圖2 合浦珠母貝不同群體雜交子一代雜交優勢率Fig. 2 Heterosis in hybrids of P. fucata
雜交育種的前提是各品種配合力的測定, 對雜交組合的設計和親本的選育有重要指導意義。一般配合力是指某一種群和其他種群雜交后, 雜交后代在某個數量性狀上表現的平均值, 一般配合力高的親本才能產生均值高的子代。本研究結果表明, 三亞親本具有較高的一般配合力。特殊配合力是指兩個特定種群雜交時, 雜種后代在某個數量性狀上的表現能力, 主要是來自兩個親本之間基因的非加性效應。這一效應一般只在雜交后才表現出來, 較難在上下代之間穩定遺傳。通過育種比較容易操縱和積累的是GCA效應, 而SCA只能通過保持父母本兩個雜種優勢群的辦法來保持或提高。本研究結果表明, 三亞群體雌性各性狀的一般配合力最大, 而北海雄性的殼高和殼寬一般配合力最大, 而谷龍春[15]等在研究這三個地理種群雙列雜交家系(出苗60日齡)時發現北海的殼長和殼高的一般配合力最大, 但沒有區分雌雄的差異。在特殊配合力方面, 該研究中, S×X組合殼高和體質量的特殊配合力最大, S×B組合的殼長和殼寬特殊配合力最大, 谷龍春[15]等對雙列雜交家系的研究表明三亞-北海的殼長、殼高特殊配合力最大。兩者有相同也有相異之處, 可能與貝的生長階段不同有關, 因為貝月齡越小, 受環境影響越大。
雜交優勢是指2個或2個以上不同遺傳類型的物種、品種或自繁或雜交產生的雜種子一代, 在生長、成活、抗逆性、適應性、生產性能等方面均優于雙親的現象。該研究的6個雜交群體在4個生長指標中均表現出一定程度的雜種優勢(0.24%~ 52.62%)。作物配合力的研究表明組合間雜種優勢的大小與其特殊配合力效應有關, 該研究中S×X和S×B組合的特殊配合力最高, 其雜交優勢也較其他組合高, 與作物方面的情況相似。谷龍春等[15]也發現, S×X組合的殼長雜交優勢較高, 而S×B組合的殼高雜交優勢較高。此外, 各性狀雜種優勢表現不盡相同, 其中體質量的雜種優勢率最高。有研究表明, 正交與反交效果不一樣[27]。筆者研究也發現, S×B和S×X雜交組合在殼長、殼高、殼寬和體質量4個性狀指標上表現出很高的雜種優勢, 而其反交組合的雜交優勢程度都較低, 且在殼寬上BS組合表現出最低的雜種優勢率(0.24%)。由此可見, 以三亞群體為母本的雜交組合均表現出很大程度的雜種優勢, 與配合力的分析結果一致。
該研究用不同來源的養殖群體親本, 采用雙列雜交方式構建基礎群體, 一次共用雌性親本90個,雄性親本54個, 樣本量具有一定的代表性。此次試驗中共獲得6個具有良好的可供進一步選育的雜交組合, 與家系選育相比, 群體選育具有子代數量大、組合少、易管理等優點, 為下一步采用旋轉交配開展后續選育奠定了良好基礎。
[1] Wada K T. Genetic selection for shell traits in the Japanese pearl oyster, Pinctada fucata martensii[J]. Aquaculture, 1986, 57: 171-176.
[2] Wada K T, Komaru A. Color and weight of pearls produced by grafting the mantle tissue from a selected population for white shell color of the Japanese pearl oyster Pinctada fucata martensii (Dunker) [J]. Aquaculture, 1996, 142: 25-32.
[3] Wada K T, Komaru A. Effect of selection for shell coloration on growth rate and mortality in the Japanese pearl oyster, Pinctada fucata martensii[J]. Aquaculture, 1994, 125: 59-65.
[4] 王愛民, 閻冰, 葉力, 等. 馬氏珠母貝不同地理種群內自繁和種群間雜交子一代主要性狀的比較[J]. 水產學報, 2003, 27(3) : 200-206. Wang Aimin, Yan Bing, Ye Li, et al. Comparison on main traits of F1 from matings and crosses of different geographical populations in Pinctada martensii[J]. Journal of Fisheries of China, 2003, 27(3) : 200-206.
[5] 湯健, 劉文廣, 林堅士, 等. 9 個馬氏珠母貝家系的中期生長性狀評估[J]. 南方水產科學, 2011, 7(5): 30-36. Tang Jian, Liu Wenguang, Lin Jianshi, et al. Evaluation on mid-term growth of 9 families of pearl oyster Pinctada facata[J]. South China Fisheries Science, 2011, 7(5): 30-36.
[6] 吳曼, 劉寶鎖, 黃桂菊, 等. 合浦珠母貝全同胞家系貝殼珍珠質顏色分析[J]. 南方水產科學, 2014, 10(6): 44-50. Wu Man, Liu Baosuo, Huang Guiju, et al. Analysis of shell nacre color in full-sib families of pearl oyster (Pinctada fucata)[J]. South China Fisheries Science, 2014, 10(6): 44-50.
[7] 郭華陽, 張殿昌, 李恒德, 等. 合浦珠母貝幼體生長性狀的遺傳力及其相關性分析[J]. 湖北農業科學, 2011, 50(21): 4441-4444. Guo Huayang, Zhang Dianchang, Li Hengde, et al. Estimates of heritability and genetic correlations for the growth of larvae of Pinctada fucata[J]. Hubei Agricultural Sciences, 2011, 50(21): 4441-4444.
[8] 許成帥, 范嗣剛, 黃桂菊, 等. 合浦珠母貝家系遺傳多樣性與性狀相關性[J]. 廣東農業科學, 2013, 40(12): 156-158. Xu Chengshuai, Fan Sigang, Huang Guiju, et al. The correlation between genetic diversity and phenotypes of Pinctada fucata families[J]. Guangdong Agricultural Sciences, 2013, 40(12): 156-158.
[9] 牛志凱, 劉寶鎖, 張東玲, 等. 合浦珠母貝3個地理群體雜交后代生長性狀和閉殼肌拉力的比較分析[J].南方水產科學, 2015, 11(1): 26-32. Niu Zhikai, Liu Baosuo, Zhang Dongling, et al. Comparative analysis of growth traits and shell-closing strength among hybrid populations from three geographical groups of pearl oyster (Pinctada fucata)[J]. South China Fisheries Science, 2015, 11(1): 26-32.
[10] 張希, 邢孔武, 喻達輝, 等. 中藥對合浦珠母貝插核手術傷口愈合效果和珍珠質量的影響[J]. 海洋科學, 2011, 35(6): 25-29. Zhang Xi, Xing Kongwu, Yu Dahui, et al. Impact of traditional Chinese medicines on recuperation of nucleus inserting cut and pearl quality in pearl oyster Pinctada fucata[J]. Marine Sciences, 2011, 35(6): 25-29.
[11] 喻達輝, 賈曉平, 陳素文, 等. 合浦珠母貝卵子電擊脈沖參數的初步研究[J]. 海洋科學, 2005, 29(8): 17-21. Yu Dahui, Jia Xiaoping, Chen Suwen, et al. Optimization of pulse parameters in electroporation of Pinctada fucata eggs[J]. Marine Sciences, 2005, 29(8): 17-21.
[12] 何毛賢, 史兼華, 林岳光, 等. 馬氏珠母貝生長性狀的相關分析[J]. 海洋科學, 2006, 30(11): 1-4. He Maoxian, Shi Jianhua, Lin Yueguang, et al. The growth trait correlated responses in pearl oyster Pinctada martensi[J]. Marine Sciences, 2006, 30(11): 1-4.
[13] 黎火金, 劉寶鎖, 羅會, 等. 合浦珠母貝幼貝生長性狀的遺傳參數估計[J]. 中國水產科學, 2013, 20(6):1182-1187. Li Huojin, Liu Baosuo, Luo Hui, et al. Estimates of genetic parameters for growth traits in juvenile pearl oyster Pinctada fucata[J]. Journal of Fishery Sciences of China, 2013, 20(6): 1182-1187.
[14] 王愛民, 石耀華, 周志剛.馬氏珠母貝不同地理種群內自繁和種群間雜交子一代形態性狀參數及相關性分析[J] .海洋水產研究, 2004, 25(3) : 39-45. Wang Aimin, Shi Yaohua, Zhou Zhigang. Morphological trait parameters and their correlations of the first generation from matings and crosses of geographical populations of Pinctada martensii(Dunker)[J]. Marine Fisheries Research, 2004, 25(3) : 39-45.
[15] 谷龍春, 李金碧, 喻達輝, 等.合浦珠母貝雙列雜交家系的建立與遺傳分析[J]. 水產學報, 2010, 34(1) : 26-31. Gu Longchun, Li Jinbi, Yu Dahui, et al. Establishment and genetic analysis of complete diallel cross families of pearl oyster(Pinctada fucata)[J]. Journal of Fisheries of China, 2010, 34(1) : 26-31.
[16] Miguel A, Rio-portilla D, Andy R B. Larval growth, juvenile size and heterozygosity in laboratory reared mussels, Mytilus edulis[J]. Exp Mar Biol Ecol, 2000, 254(1): 1-17.
[17] Liu X L, Chang Y Q, Xiang J H. Studies on hybridization effects of different geographic population of Chlamys farreri[J]. Acta Oceanologica Sinica, 2003, 25(2): 255-263.
[18] English L J, Maguire G B, Ward R D. Genetic variation of wild and hatchery stocks of the Pacific oyster, Crassostrea gigas(Thunberg), in Australia[J]. Aquaculture, 2000, 187(3/4): 283-298.
[19] Rahman M A, Uehara T, Pearse J S. Hybrids of two closely related tropical sea urchins (Genus Echinometra): Evidence against postzygotic isolating mechanisms[J]. Biol Bull, 2001, 200(1): 97-106.
[20] Rahman M A, Uehara T, Lawrence J M. Growth and heterosis of hybrids of two closely related species of Pacific sea urchins (Genus Echinometra) in Okinawa[J]. Aquaculture, 2005, 245(1/2/3/4): 121-133.
[21] Cruz P, Ibarra A M. Larval growth and survival of two catarina scallop (Argopecten circularis, Sowerby, 1835) populations and their reciprocal crosses[J]. Expl Mar Biol Ecol, 1997, 212(1): 95-110.
[22] Misamore M, Browdy C L. Evaluating hybridization potential between Penaeus setiferus and Penaeus vannamei through natural mating, artificial insemination and in vitro fertilization[J]. Aquaculture, 1997, 150(1/2): 1-10.
[23] Bierne N, Beuzart I, Vonan V. Microsatellite-associated heterosis in hatchery-propagated stocks of the shrimp Penaeus stylirostris[J]. Aquaculture, 2000, 184(3/4): 203-219.
[24] 陳飛飛. 合浦珠母貝群體選育及鰓結構研究[D]. 上海: 上海海洋大學, 2012: 29-43. Chen Feifei. Studies on mass selectiand gill microstructure of Pinctada fucata[D].Shanghai: Shanghai Ocean University, 2012: 29-43.
[25] 毛友純, 徐慶國, 胡志明. 雜交早稻農藝性狀的配合力研究[J]. 湖南農業大學學報(自然科學版), 2005, 2: 115-119. Mao Youchun, Xu Qingguo, Hu Zhiming. Studies on the combining ability of agronomic characters in early-seasoned hybrid rice[J]. Journal of Hunan Agricultural University(Natural Sciences), 2005, 2: 115-119.
[26] 盛志廉, 陳瑤生.數量遺傳學[M].北京: 科學出版社, 2001: 319-320. Sheng Zhilian, Chen Yaosheng. Quantitative genetics[M]. Beijing: The Science Publishing Company, 2001: 319-320.
[27] 王愛民, 丁小雷, 鄧鳳嬌, 等.馬氏珠母貝大亞灣和三亞野生群體內自繁及群體間雜交一代的RAPD分析[J].海洋水產研究, 2003, 24(4): 19-25. Wang Aimin, Ding Xiaolei, Deng Fengjiao, et al. The genetic diversity of the first filial generation from matings and crosses of two wild populations(DayaBay, Guangdong and Sanya, Hainan) in Pinctada martensii (Dunker) [J]. Marine Fisheries Research, 2003, 24(4): 19-25.
[28] 丁小雷, 鄧鳳嬌, 王愛民, 等.野生馬氏珠母貝子一代的遺傳多樣性分析[J].動物學雜志, 2003, 38 (1) : 2-7. Ding Xiaolei, Deng Fengjiao, Wang Aimin, et al. Genetic diversity of two wild Pinctata martensi populations′F1 generations[J].Chinese Journal of Zoology, 2003, 38 (1) : 2-7.
(本文編輯: 譚雪靜)
Growth performances and genetic analysis of complete diallel crosses of Pinctada fucata based on three cultured populations
HUANG Gui-ju1, CHEN Fei-fei1, 2, LIU Bao-suo1, FAN Si-gang1, XING Kong-wu1, YU Da-hui1
(1. Key Laboratory of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture, South China Sea Resource Exploitation and Protection Collaborative Innovation Center(scs-REPIC), Chinese Academy of Fishery Sciences, Guangzhou 510300, China; 2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China)
Aug., 19, 2014
Pinctada fucata; diallel population; growth; combining ability; heterosis
To improve the genetic quality of pearl oysters (Pinctada fucata) and to develop new species, in this study, we developed complete diallel crosses using three stocks of pearl oysters from Beihai (B), Xuwen (X), and Sanya (S). We then evaluated the growth performances of the eight-months-old F1generation with respect to shell length, shell height, shell width, and body weight. In addition, we analyzed hybrid vigor and general and specific combining abilities with respect to these four growth traits. The results showed that the growth performances of the crossbreds were significantly better than those of the purebreds, as measured by the four traits. of the crossbreds, the growth performances of S(♀)×B(♂)and S(♀)×X(♂)were significantly higher than any other crosses with respect to the four traits. ANOVA results showed no significant differences among the three purebreds. The Sanya stock had the best general combining abilities, while the S(♀)×X(♂)cross had the best specific combining abilities. All the crossbreds showed heterosis in the four growth traits to some extent, ranging from 0.24% to 52.62%. The heterosis differed for different combinations or different traits. While the heterosis in S(♀)×B(♂)and S(♀)×X(♂)were greater than those of the reverse crosses B(♀)×S(♂)or X(♀)×S(♂)for the four growth traits, the shell width of the cross B(♀)×S(♂)had the lowest heterosis rate (0.24%). The heterosis related to body weight were the highest of the four traits for all crosses. These six genetically improved crossbreds can be used as base populations to further breed new varieties.
S968.31+6.1
A
1000-3096(2016)02-0026-09
10.11759//hykx20140819003
2014-08-19;
2014-12-19
現代農業產業技術體系建設專項(CARS-48); 廣東省海洋漁業科技推廣專項(A201301A02, A201301A08, Z2014003)
黃桂菊(1978-), 女, 廣東肇慶人, 碩士, 副研究員, 從事水產生物技術研究, E-mail: xaxhgj@126.com; 喻達輝, 通信作者, 研究員, E-mail: pearlydh@163.com
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
