黃東海夏、冬季顆石藻群落及其分布研究
2016-07-06 10:29:53李佳芮紀大偉翟偉康王園君國家海洋信息中心天津300171
海洋科學 2016年2期
張 健, 李佳芮, 紀大偉, 翟偉康, 王園君(國家海洋信息中心, 天津 300171)
?
黃東海夏、冬季顆石藻群落及其分布研究
張 健, 李佳芮, 紀大偉, 翟偉康, 王園君
(國家海洋信息中心, 天津 300171)
2009年作者對中國黃東海海域夏季(7月20日至9月1日)與冬季(12月23日至2月5日)的兩個季度月的顆石藻群落與分布進行調查研究。2009年夏季, 中國黃東海海域調查區共發現21種顆石藻, 其優勢物種分別為赫氏艾密里藻(Emiliania huxleyi)、大洋橋石藻(Gephyrocapsa oceanica)、纖細傘球藻(Umbellosphaera tenuis)和深水花球藻(Florisphaera profunda)。顆石藻細胞豐度為0.23×103~17.62×103個/L, 平均值為2.84×103個/L。2009年冬季, 中國黃東海海域調查區共發現20種顆石藻, 其優勢物種分別為赫氏艾密里藻(E. huxleyi)、大洋橋石藻(G. oceanica)、深水花球藻(F. profunda)和纖細傘球藻(U. tenuis)。顆石藻的細胞豐度為0.12×103~35.35×103個/L, 平均值為3.84×103個/L。本文系統地研究了顆石藻在我國黃、東海陸架海域的分布(特別是垂直分布), 并對其作出了描述與分析, 以期為關于中國海顆石藻群落分布等基礎性研究提供可靠資料。
顆石藻; 群落分布; 黃東海; 夏冬季
[Fundation: National key basic research development planning project (2006CB400605) ]
顆石藻(coccolithophore)指的是生活在現今海洋中, 在生活史的某些階段具有碳酸鈣質的外殼, 并且在海洋生態系統中發揮著重要作用的活的顆石藻類群[1]。顆石藻在整個海洋的碳循環過程中扮演著重要的角色, 是當今海洋生態系統中最為重要的基礎生產者之一, 尤其是在海洋的無機碳循環過程中起到重要作用。它具有獨特的碳酸鹽反向泵(carbonate counter pump)和有機碳泵(organic carbon pump)機制,對于海洋中碳的生物地球化學循環具有重要作用[2]。
顆石藻廣泛地生活于全球范圍內的大洋和近海水域中, 并且更趨向于生活在相對高硝酸鹽、低磷酸鹽、高光和低微型浮游動物豐度的水體中, 它們是大洋和高緯度海洋中發生水華的常見類群, 同時也是十分重要的一個浮游植物類群。到目前為止, 已經有接近70屬的200多種顆石藻物種被記錄下來。在過去的地質年代中, 它們對于大洋底的碳酸鈣沉積具有著非常重大的貢獻。顆石藻每年都會在全球范圍內發生很多次水華(每毫升海水里含有的顆石藻細胞數目超過1000個), 因在水面上漂浮的顆石粒和顆石藻像一面面反射陽光的小鏡子, 使水面光亮無比,因此顆石藻的水華又被稱為白水(white water)[3]。
中國近海的顆石藻群落分布情況(尤其是垂直分布情況)至今并不十分清楚。本文基于2009年黃、東海夏季與冬季的顆石藻群落分布調查研究, 對調查海域真光層0~200 m水體中的顆石藻的物種組成、細胞豐度、優勢種、群落多樣性指數和水柱積分豐度分布及其季節變化進行了分析研究。由于目前對于顆石藻還沒有國際統一的定量采樣和樣品分析方法, 本研究應用在國際上普遍認可并能較真實反映定量信息的偏光顯微鏡分析法[4], 系統的研究了顆石藻在我國黃、東海陸架海域的分布(特別是垂直分布), 并對其作出了描述與分析, 以期為中國近海顆石藻物種多樣性研究, 特別是顆石藻群落對全球氣候變化響應等后續研究提供基礎資料。
1 調查海域與研究方法
1.1 調查海域
本研究基于2009年在中國近海所進行的CHOICE-C(973)項目的走航調查。分別于2009年夏季(7月20日至9月1日)與冬季(12月23至2月5 日)對我國黃東海海域(27.00°~36.50°N, 121.50°~ 127.00°E)進行了兩個季度月的包括水化學、化學和生物在內的綜合嵌套式外業調查。夏季和冬季, 調查區分別設置調查采樣站位17個和19個(圖1, 圖2)。同時, 各調查區設4個斷面(斷面1、斷面2、斷面3、斷面4), 參照《海洋調查規范》中標準層的采樣原則[5],水深小于60 m的站位, 取表層、中層和底層3層; 水深大于60 m的站位, 取表層、次表層、中層和底層4層。各調查區海域實驗站位信息見表1和表2。顆石藻樣品使用調查船上的梅花式海鳥CTD(12個12L 的Go-Flo)進行海水取樣, 然后將取得的海水倒入1L的聚乙烯(PE)瓶中, 立刻加入適量弱的堿性福爾馬林溶液固定, 使樣品中的甲醛濃度為1%~2%[2]。溫度和鹽度由調查船上的CTD現場進行測定。
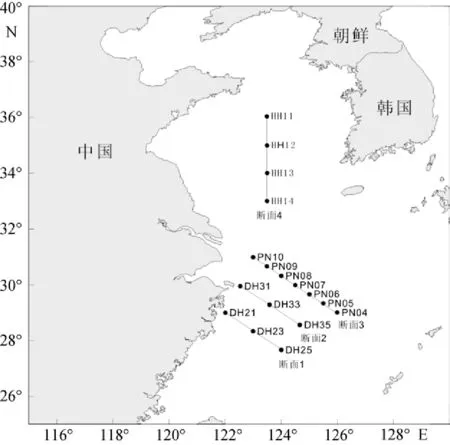
圖1 2009年夏季調查站位圖Fig. 1 Stations surveyed in the summer of 2009
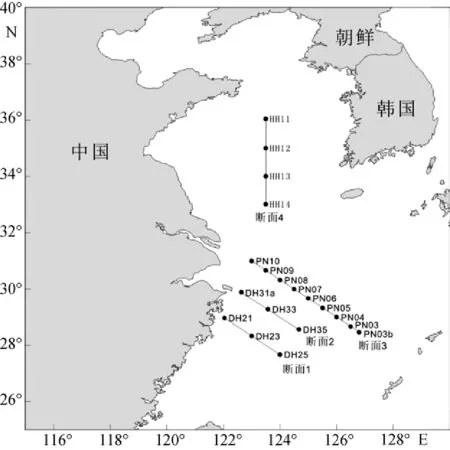
圖2 2009年冬季調查站位圖Fig. 2 Stations surveyed in the winter of 2009
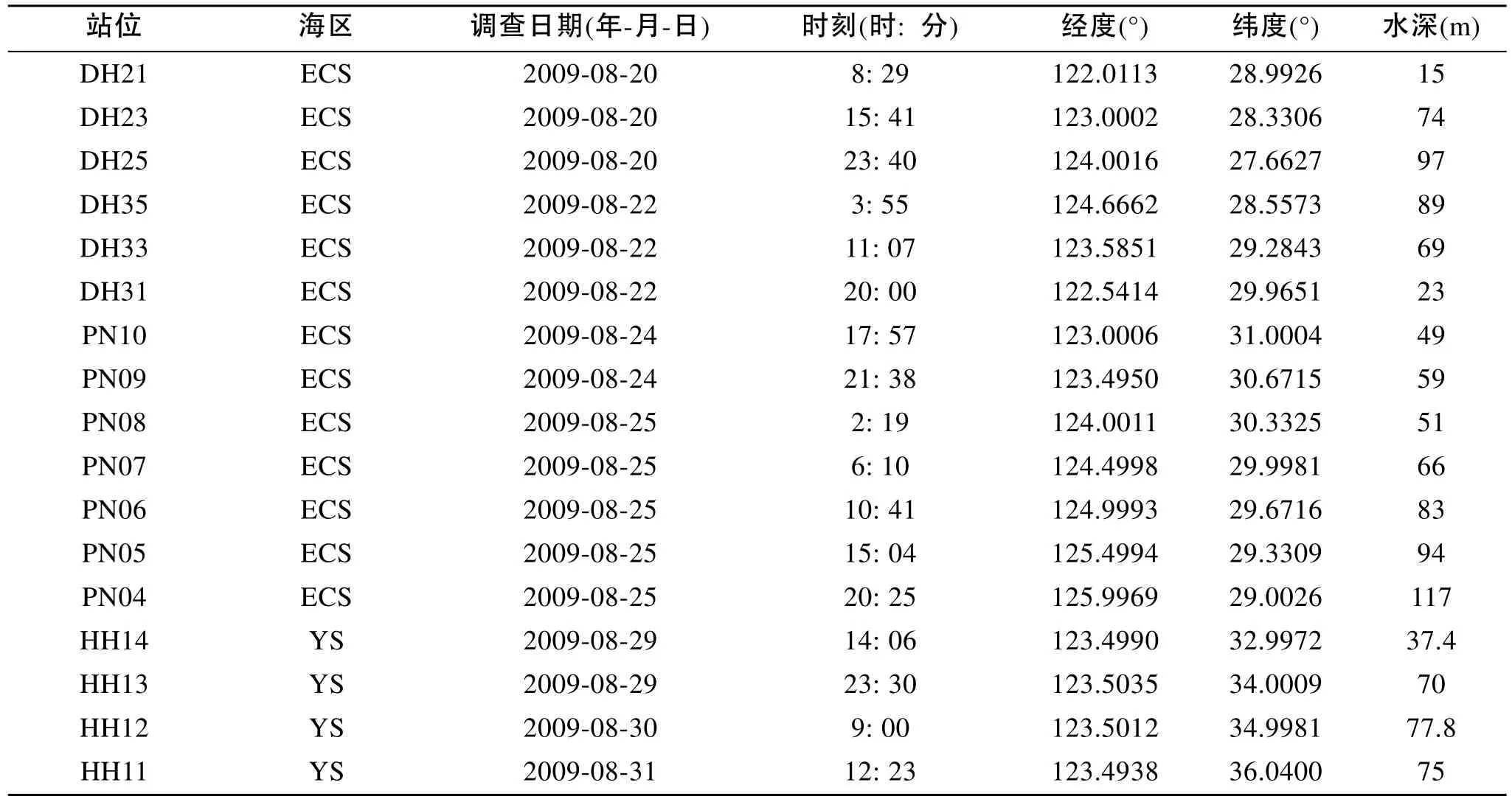
表1 2009年夏季調查區實驗站位基本信息Tab. 1 Basic information of sampling stations in the survey area in the summer of 2009
1.2 研究方法
將取得的樣品帶回實驗室, 從1L的聚乙烯瓶(PE)中取400 mL的顆石藻亞樣品過濾到聚碳酸酯(polycarbonate)濾膜(直徑25 mm, 孔徑0.45 μm)上,過濾壓力小于100 mmHg。樣品過濾后, 立即用弱堿性的蒸餾水潤洗濾膜以除去多余的鹽分。然后, 將濾膜放入塑料培養皿中, 擱置于烘箱中, 于50 ℃下進行烘干處理。最后, 將烘干后的濾膜取出, 用剪刀剪取適當大小, 放到玻璃制的載玻片上, 滴上適量的加拿大中性樹脂(Canada Balsam), 然后用蓋玻片封片。制片完成后的樣品隨即放入烘箱(50℃ )中烘干2~3 d。
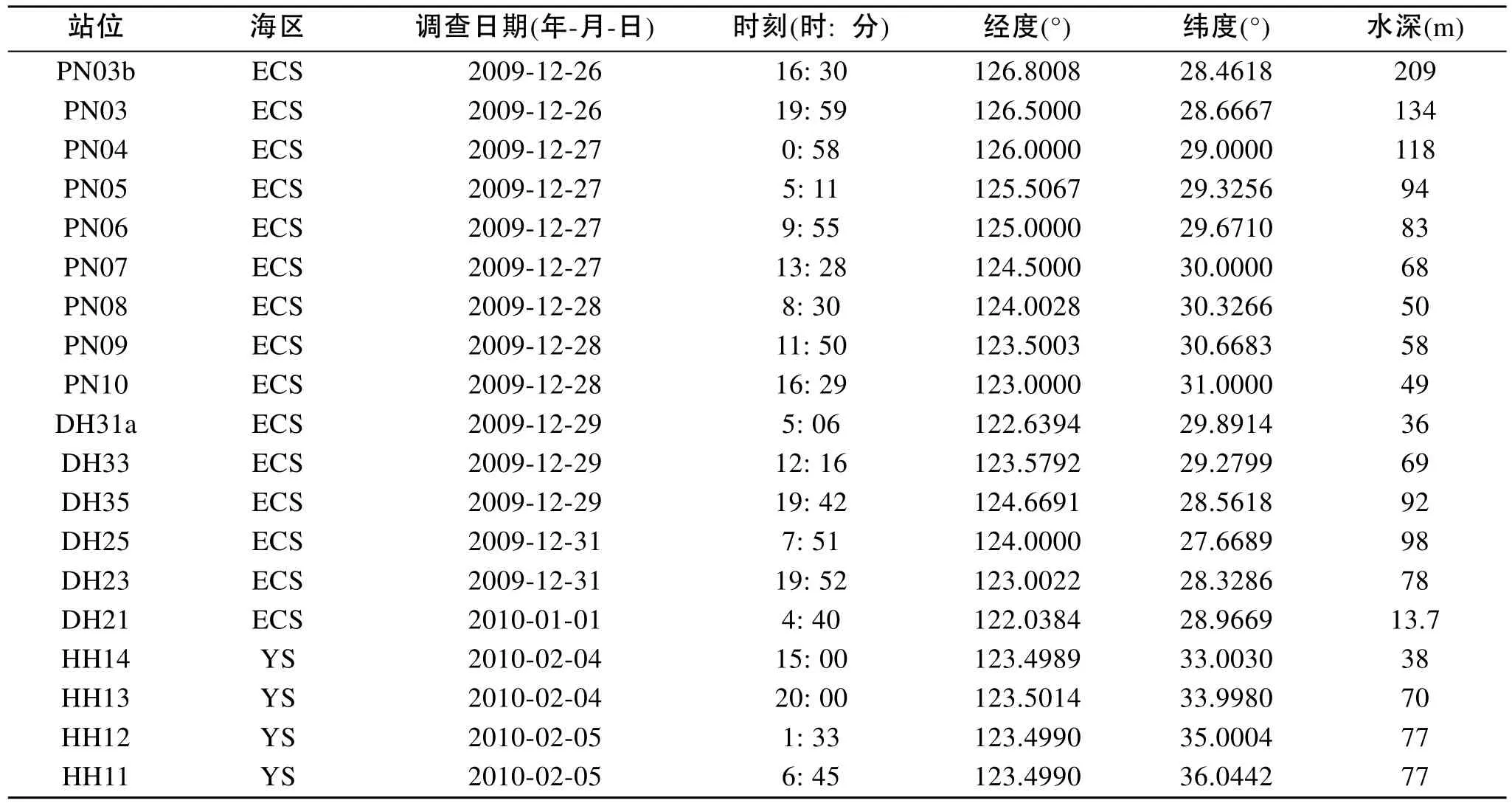
表2 2009年冬季調查區實驗站位基本信息Tab. 2 Basic information of sampling stations in the survey area in the winter of 2009
顆石藻的數量統計是在偏光顯微鏡(Motic BA300pol)1000×下進行的。根據雙折射的特性對游離的顆石粒以及顆石球進行鑒定和計數。按照統計學要求, 隨機選擇視野并且盡可能鏡檢到300個顆石粒或100個顆石球。優勢種的鑒定則根據顆石藻形態學特征[6-7]使用掃描電子顯微鏡(SEM)進行物種鑒定[8], 同時測定其平均最大粒長和平均球徑。
顆石藻的細胞豐度計算公式采用修改自Bollmann的公式:

其中A為顆石藻細胞豐度(個/ L); N為在每個玻片上所觀察到的視野個數; a為N個視野中的顆石藻數目; b為過濾的樣品體積(mL); S為濾膜的有效過濾區域面積; s為偏光顯微鏡1000 ×下的單個視野面積。依據每單位細胞所含顆石粒的數將形成顆石球的各物種顆石粒轉換為細胞數量[7-8]。
顆石藻群落多樣性指數和優勢度分析, 各指數計算如下所示:
物種多樣性指數的計算采用香農–威納指數(H′, Shannon-Wiener index):
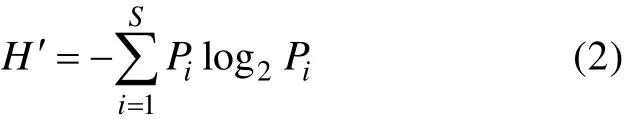
物種均勻度采用Pielou指數(J), 計算公式為:

優勢度指數(Y), 其計算公式為:

式中, N為采集樣品中所有顆石藻的總個體數, S為樣品中的物種總數, ni為第i種的總個體數, Pi= ni/ N為第i種在樣品中的細胞豐度概率, fi為該種在各樣品中出現的頻率。
計算得到的數據運用surfer9.3和CorelDRAW軟件進行制圖作業, 從而獲得顆石藻的分布規律圖。
2 結果與討論
2.1 物種組成與優勢種
2009年夏季航次(圖1), 在黃東海海域調查區共發現顆石藻21種, 其中絕大多數為異晶顆石藻(heterococcolithophore), 只有極少數為同晶顆石藻(holococcolithophore)[9]。黃東海夏季顆石藻的主要優勢種為赫氏艾密里藻(Emiliania huxleyi)、大洋橋石藻(Gephyrocapsa oceanica)、纖細傘球藻(Umbellosphaera tenuis)、深水花球藻(Florisphaera profunda)、卡特旋筏球藻(Helicopontosphaera carteri)以及希布格臍球藻(Umbilicosphaera sibogae)。赫氏艾密里藻與大洋橋石藻為絕對優勢種, 其相對細胞豐度分別為36.77%和32.90 %。兩者的出現頻率均為1.00; 纖細傘球藻的相對細胞豐度為14.87 %, 出現頻率為0.69。2009年夏季黃東海海域顆石藻的豐度百分比、出現頻率和優勢度如表3所示。
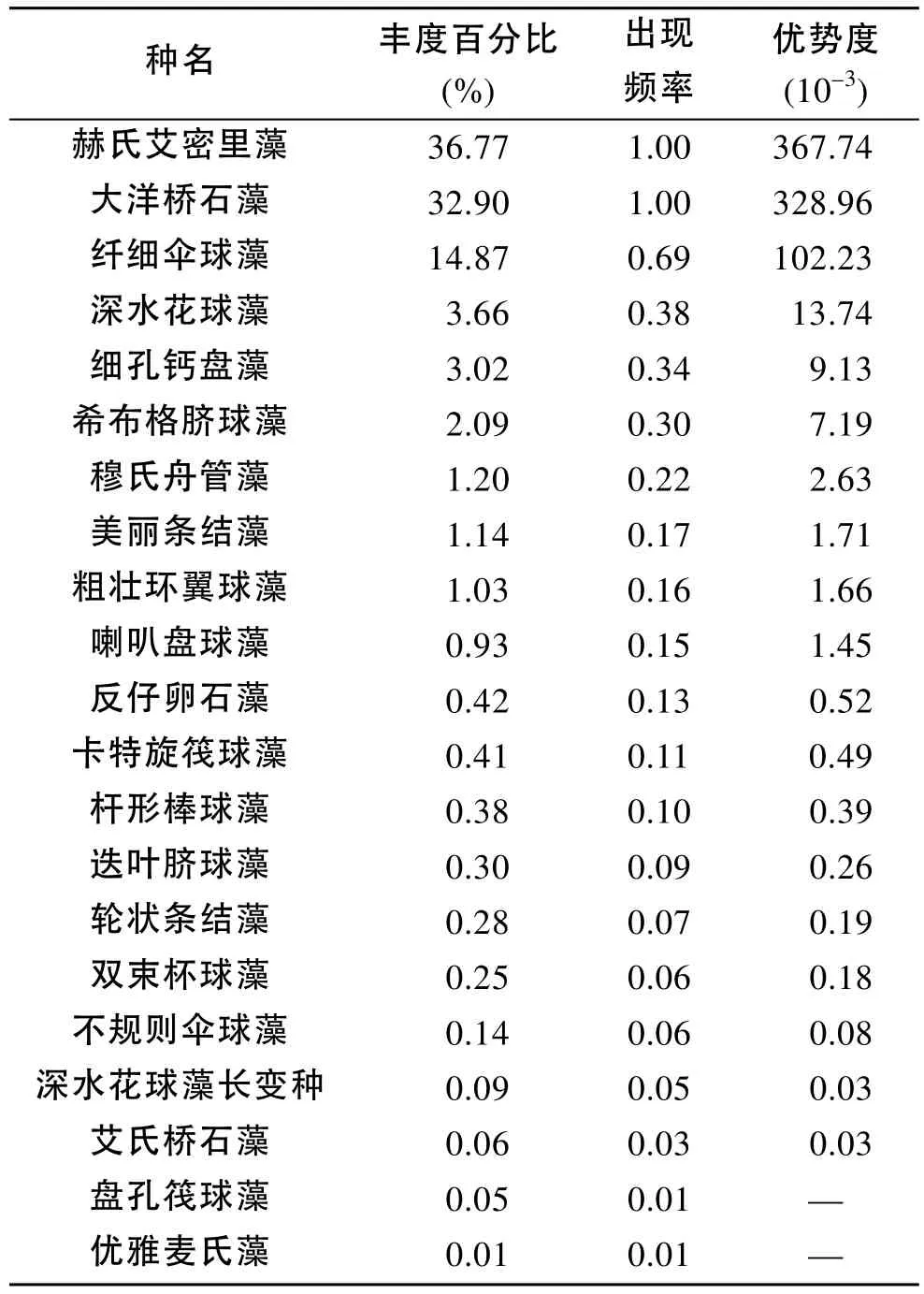
表3 2009年夏季調查區顆石藻物種組成Tab. 3 Composition of coccolithophore species in the summer of 2009
2009年冬季航次(圖2), 在黃東海海域調查區內共發現顆石藻20種。其中絕大多數種類為異晶顆石藻(heterococcolithophore), 少數為同晶顆石藻(holococcolithophore)。冬季黃東海海域顆石藻的主要優勢種為赫氏艾密里藻(E. huxleyi)、大洋橋石藻(G. oceanica)、深水花球藻(F. profunda)、纖細傘球藻(U. tenuis)、美麗條結藻(S. pulchra)和希布格臍球藻(U. sibogae)。赫氏艾密里藻與大洋橋石藻的相對細胞豐度在調查區占有絕對優勢, 分別占到42.97%和32.06%, 兩者的出現頻率分別為1.00和0.98; 而深水花球藻的相對細胞豐度為5.60 %, 出現頻率是0.64。纖細傘球藻的相對細胞豐度與出現頻率則分別為3.54%和0.51。2009年冬季黃東海顆石藻的豐度百分比、出現頻率和優勢度如表4所示。
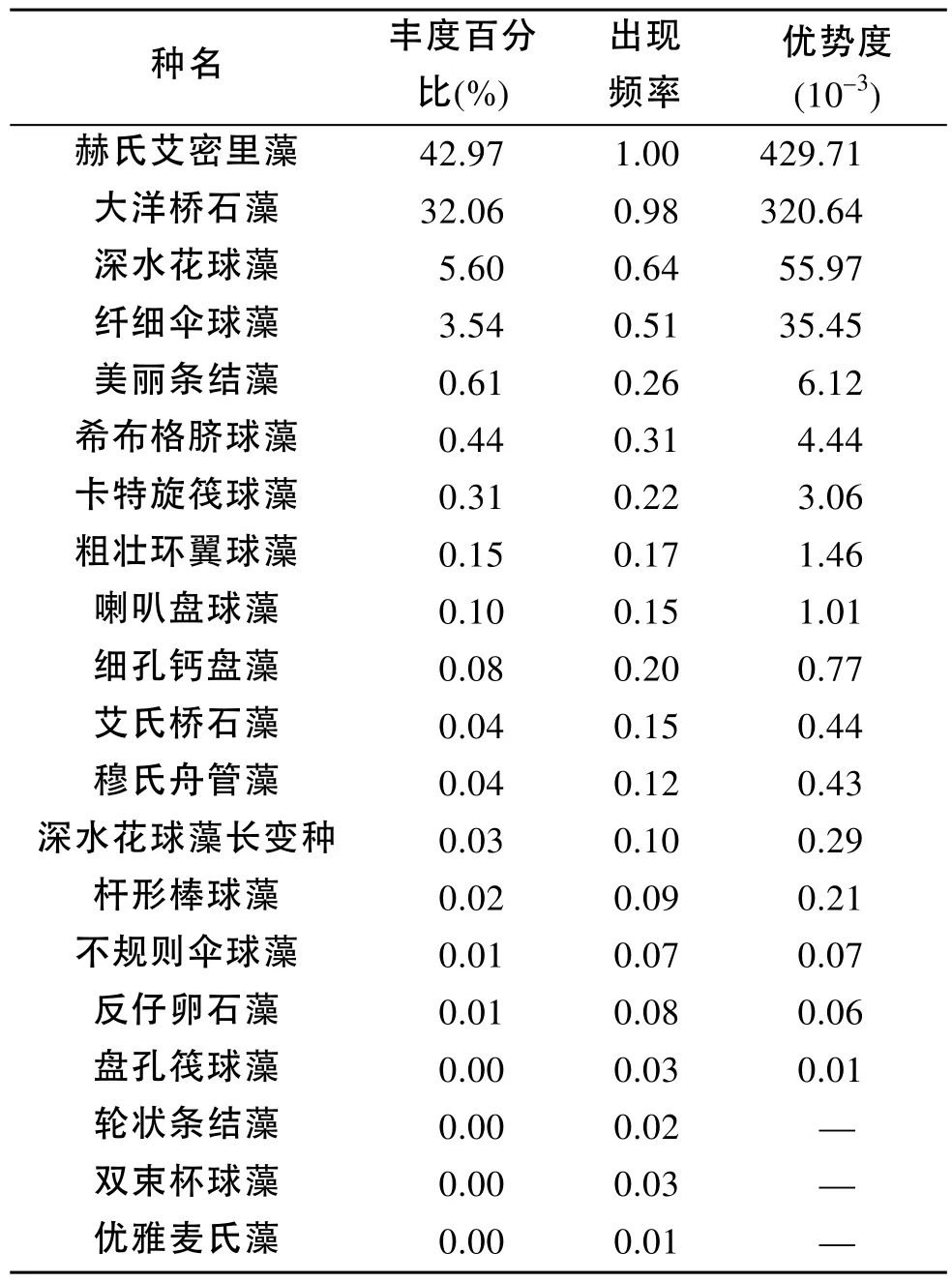
表4 2009年冬季調查區顆石藻物種組成Tab. 4 Composition of coccolithophore species in the winter of 2009
2.2 平面分布
2009年夏季, 黃東海海域調查區總的顆石藻的細胞豐度為0.23×103~17.62×103個/L, 細胞豐度的平均值為2.84×103個/L。赫氏艾密里藻的細胞豐度在0.79×103~7.4×103個/L, 平均值為1.04×103個/L。大洋橋石藻的細胞豐度為0.29×103~7.6×103個/L, 平均值為0.93×103個/L。纖細傘球藻的細胞豐度為在0 ~ 2.22×103個/L, 平均值為0.42×103個/L(圖3)。
夏季整個研究海域中, 顆石藻的分布呈現不均勻的特點, 其在調查區的某一站點會突然出現高值。這是由于夏季黃東海顆石藻的分布不光受到光和營養鹽的影響, 其受到南方來的黑潮暖流以及北方的黃海冷水團的交互影響也十分明顯, 同時, 作為中國沿海最大的沖淡水徑流—長江, 它對顆石藻群落分布的影響也是不容小視的[10]。
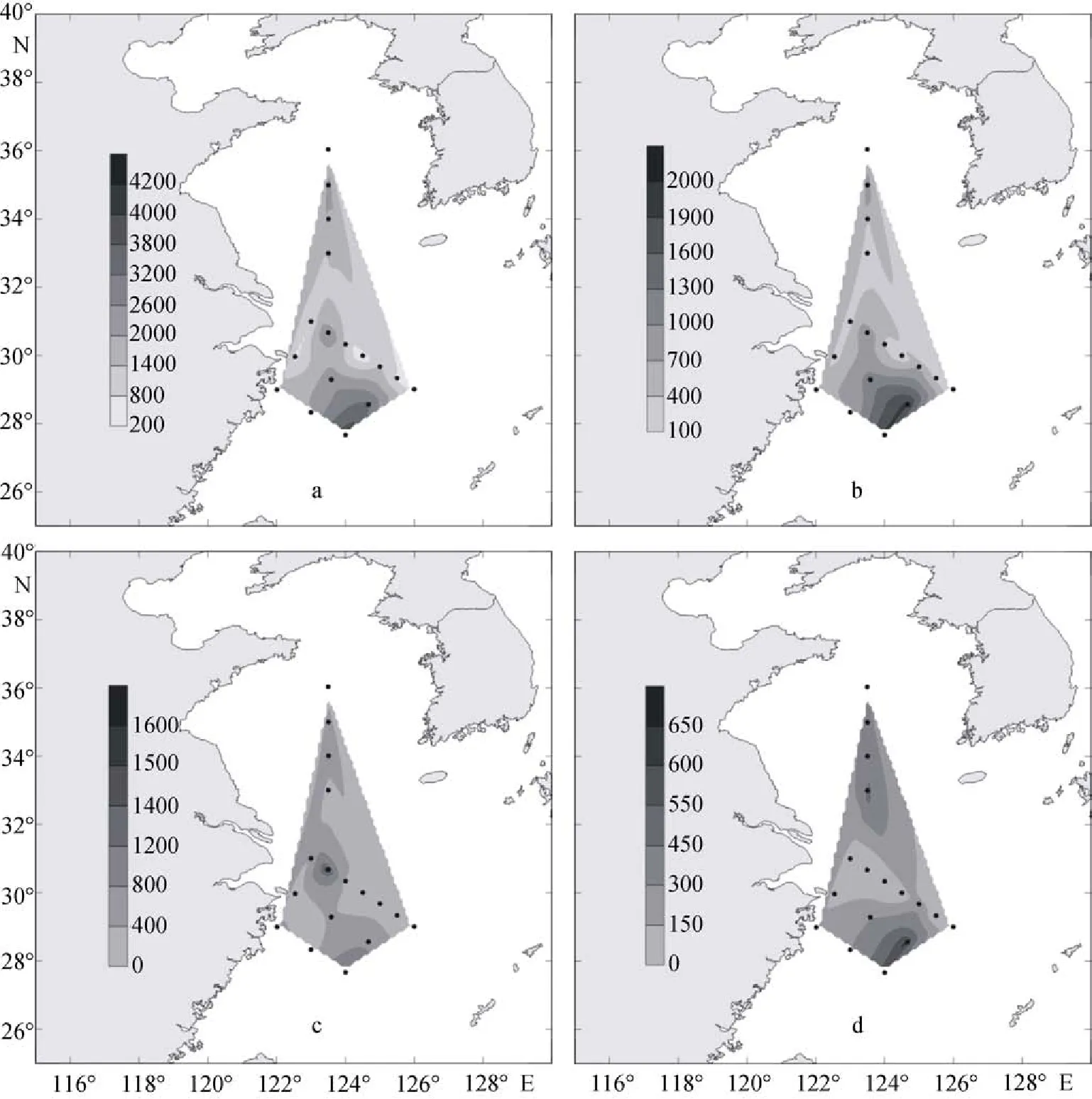
圖3 2009年夏季黃東海表層顆石藻細胞豐度分布(個/L)Fig. 3 Distribution of coccolithophore abundance in surface water in the summer of 2009 (cells/L)
冬季黃東海總的顆石藻的細胞豐度為0.12×103~ 35.35×103個/L, 平均值為3.84×103個/L, 其數值范圍相比夏季有較大的提高。赫氏艾密里藻的細胞豐度在0.06×103~16.6×103個/L, 平均值為1.23×103個/L。大洋橋石藻的細胞豐度在0~14.35×103個/L, 平均值為1.93×103個/L。深水花球藻的細胞豐度為0~ 2.20×103個/L, 平均值為0.248×103個/L(圖4)。
在整個冬季調查海域中, 顆石藻的細胞豐度較夏季顆石藻的細胞豐度要提高很多, 但是其高值區的分布范圍卻相對比較狹小, 只在調查區中部的PN斷面的一些站位出現。這是因為其受到冬季黃東海陸架區上升流、長江沖淡水和黑潮水的交互影響作用[10]。
2.3 垂直分布
2009年夏季調查區的顆石藻多分布在35 m、50 m 和75 m這3個水層當中, 細胞豐度的最大值出現在斷面1的DH25站(圖5)的50 m層水中, 達到了17.62× 103個/L, 而在該站位的35 m層和75 m層中也分別達到了14.83×103個/L和12.46×103個/L的較高值。斷面2如圖6所示。同時, 在斷面3(圖7)的PN09站也出現10.28×103個/L的較高值。夏季調查區的垂直分布呈現一種斑狀(patchy)和“牛眼”(bull’s eyes)的分布模式[11],在某些站位的某一水層中會突然出現豐度高值。
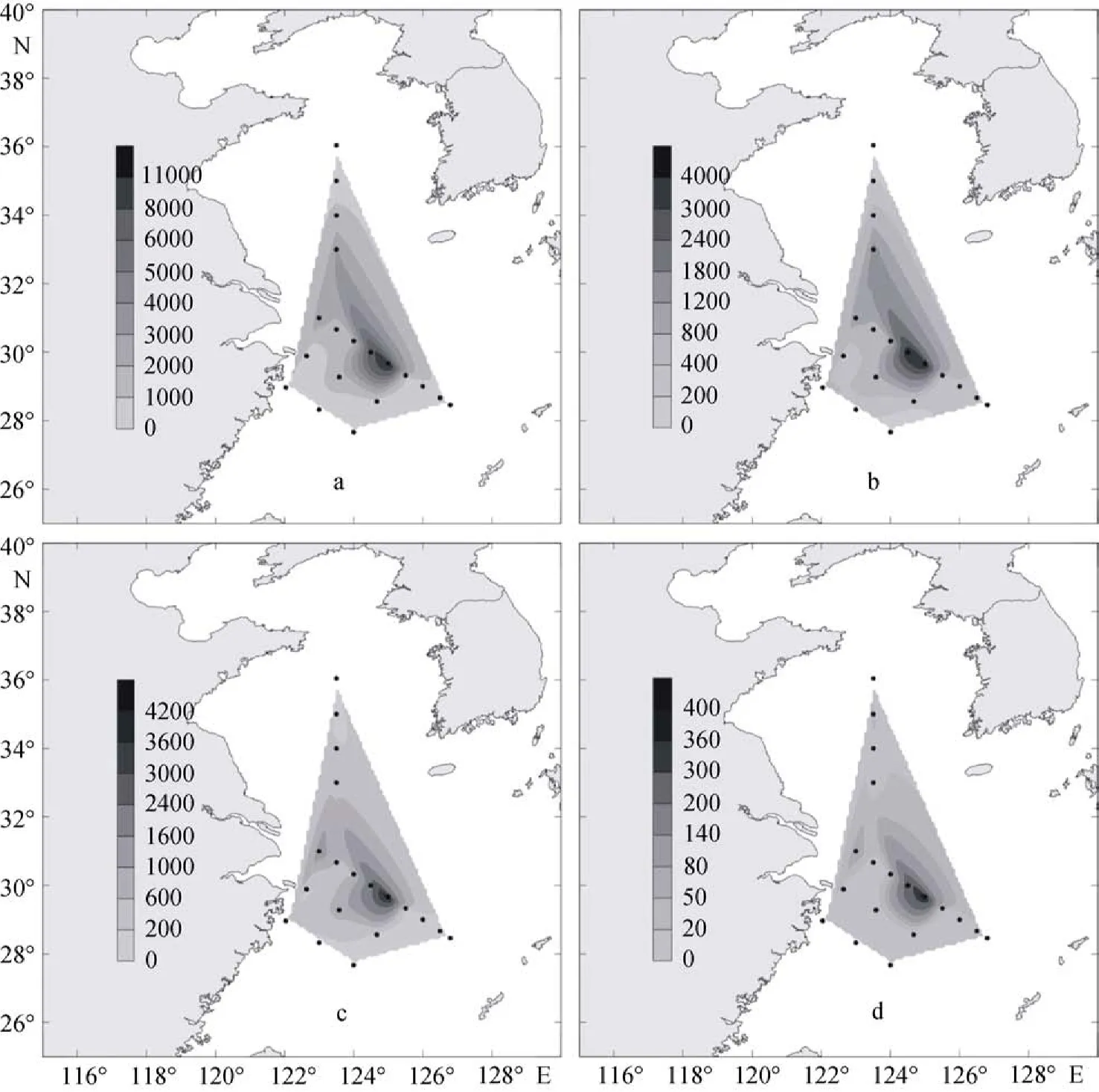
圖4 2009年冬季黃東海表層顆石藻細胞豐度分布(個/L)Fig. 4 Distribution of coccolithophore abundance in surface water in the winter of 2009 (cells/L)
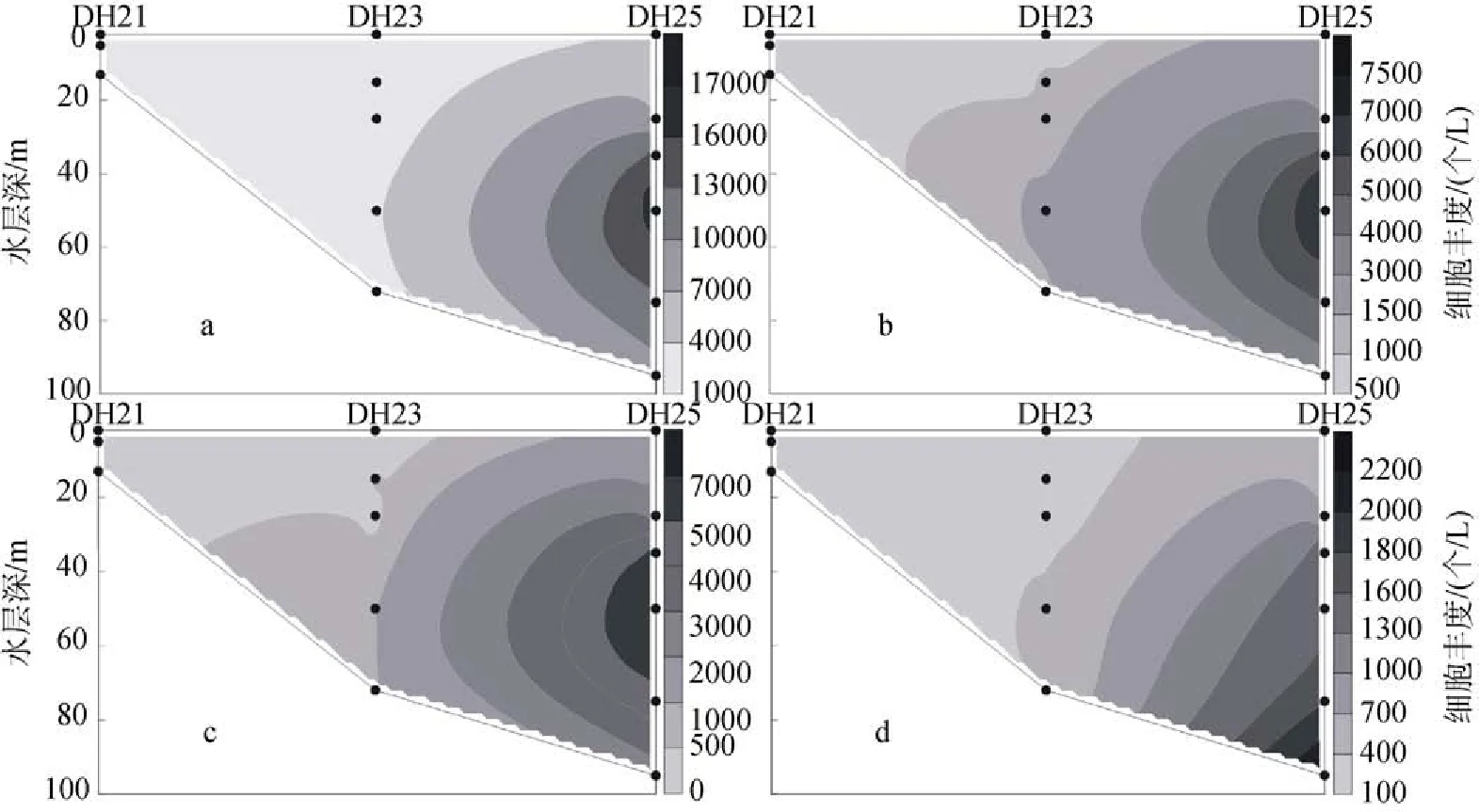
圖5 2009年夏季黃東海海域斷面1顆石藻細胞豐度分布Fig. 5 Vertical distribution of coccolithophore abundance in section 1 in the summer of 2009
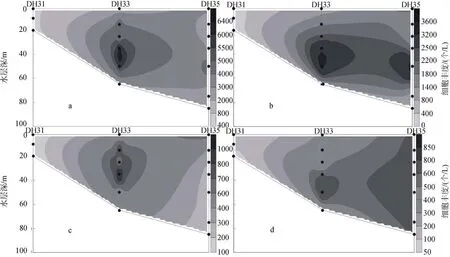
圖6 2009年夏季黃東海海域斷面2顆石藻細胞豐度分布Fig. 6 Vertical distribution of coccolithophore abundance in section 2 in the summer of 2009
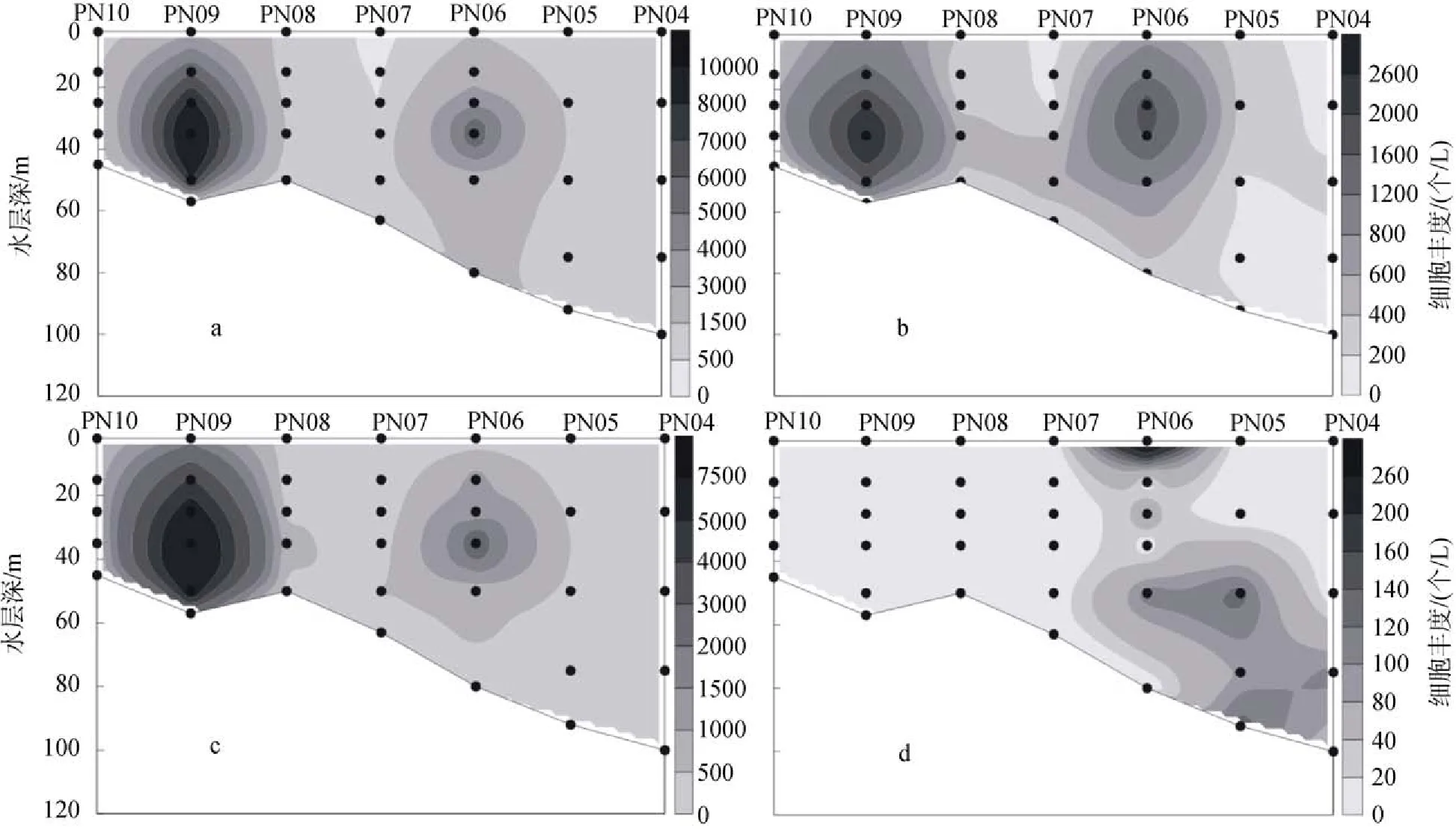
圖7 2009年夏季黃東海海域斷面3顆石藻細胞豐度分布Fig. 7 Vertical distribution of coccolithophore abundance in section 3 in the summer of 2009
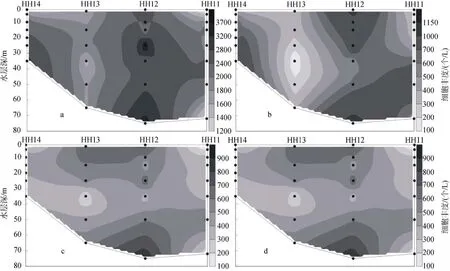
圖8 2009年夏季黃東海海域斷面4顆石藻細胞豐度分布Fig. 8 Vertical distribution of coccolithophore abundance in section 4 in the summer of 2009
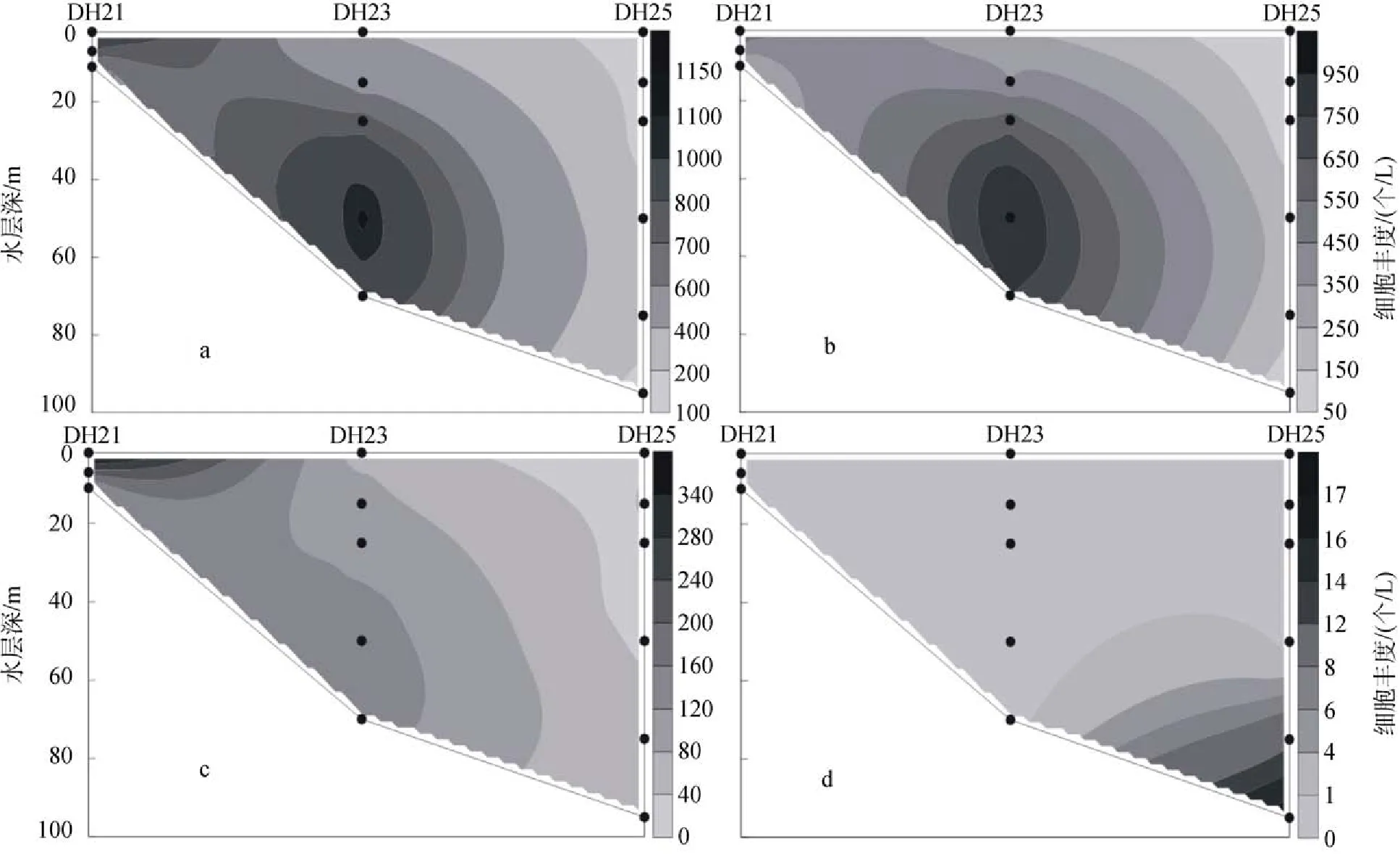
圖9 2009年冬季黃東海海域斷面1顆石藻細胞豐度分布Fig. 9 Vertical distribution of coccolithophore abundance in section 1 in the winter of 2009
2009年冬季黃東海調查區的顆石藻多分布在25 m、50 m和75 m這3個水層當中, 顆石藻細胞豐度的最大值出現在斷面3的PN06站(圖11)的50 m層中,達到了35.35×103個/L, 同時, 在該站位的25 m層和75 m層中也分別達到了26.45×103個/L和20.67×103個/L的較高值。除了PN06站以外, 在PN09站的55 m層和PN07的65 m層中也分別達到了8.66×103個/L和 8.47×103個/L的較高值。與夏季不同, 在調查區的斷面4(圖12)也出現了顆石藻細胞豐度的較高值, 其分別出現在HH13站的50 m層和HH14站的35 m層, 數值分別達到6.97×103個/L 和5.56×103個/L。冬季黃東海調查區顆石藻的垂直分布特點與夏季顆石藻的垂直分布特點較為類似, 也呈現出不均勻分布的特點, 會在某些站位的某一水層中突然出現豐度高值。
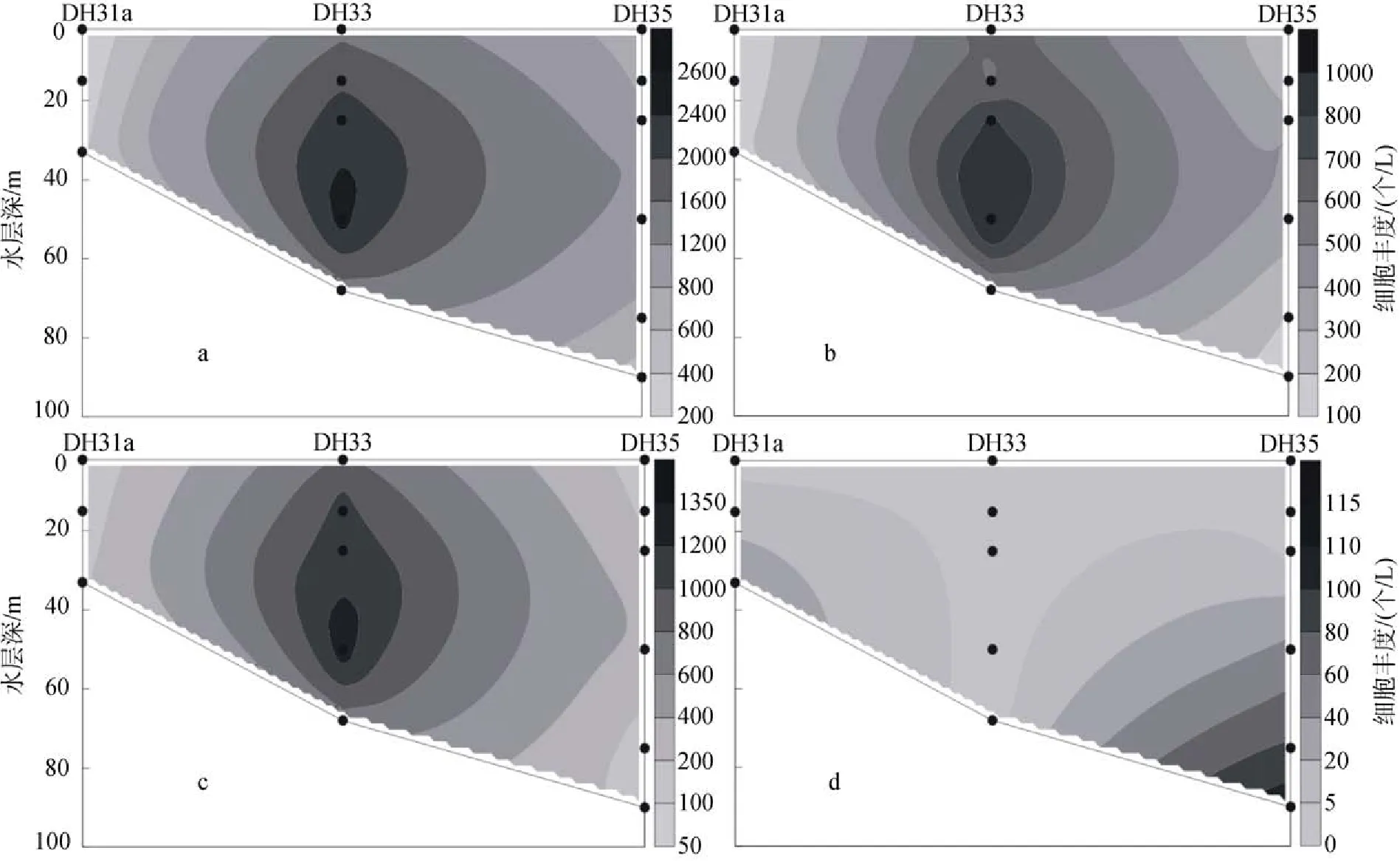
圖10 2009年冬季黃東海海域斷面2顆石藻細胞豐度分布Fig. 10 Vertical distribution of coccolithophore abundance in section 2 in the winter of 2009
2.4 多樣性指數與均勻度
2009年夏季顆石藻群落的物種多樣性指數(圖13a)為0.72~2.35, 平均值為1.84。多樣性指數在調查區的北部與南部及其鄰近海域較高, 在中部的PN08站位也存在較高的數值。而均勻度指數(圖13b)為0.49~0.99, 平均值為0.82, 其在調查區的中部與東部海域具有較高值。
2009年冬季黃東海顆石藻群落的物種多樣性指數(圖14a)為1.11~ 2.62, 平均值為1.85。物種多樣性指數的最高值出現在HH12站及其鄰近海域。整個調查區的北部、東部與南部及其鄰近海域多樣性指數較高, 較高值的出現站位分別為HH12、PN06和 DH35。調查區顆石藻的Pielou均勻度與物種多樣性指數的分布呈現出較為一致的特征, 其數值為0.57~ 0.95, 平均值為0.78(圖14b), 在調查區的中部及其鄰近海域具有較高值。
2.5 調查區表層溫鹽分布
2009年夏季, 黃東海海域調查區的表層溫鹽分布如圖15a和圖15b所示。調查區的表層溫度值為25.52~29.99℃ , 平均值為28.34℃。溫度的高值區出現在調查區的東部和南部, 最高值出現在DH33站,數值為29.99℃。而在最靠近長江口的PN10站, 溫度達到最低值, 為25.52℃。從圖15a中可以看出, 夏季黃東海調查區表層溫度分布趨勢是從北向南, 由近岸到遠海, 溫度值越來越高。
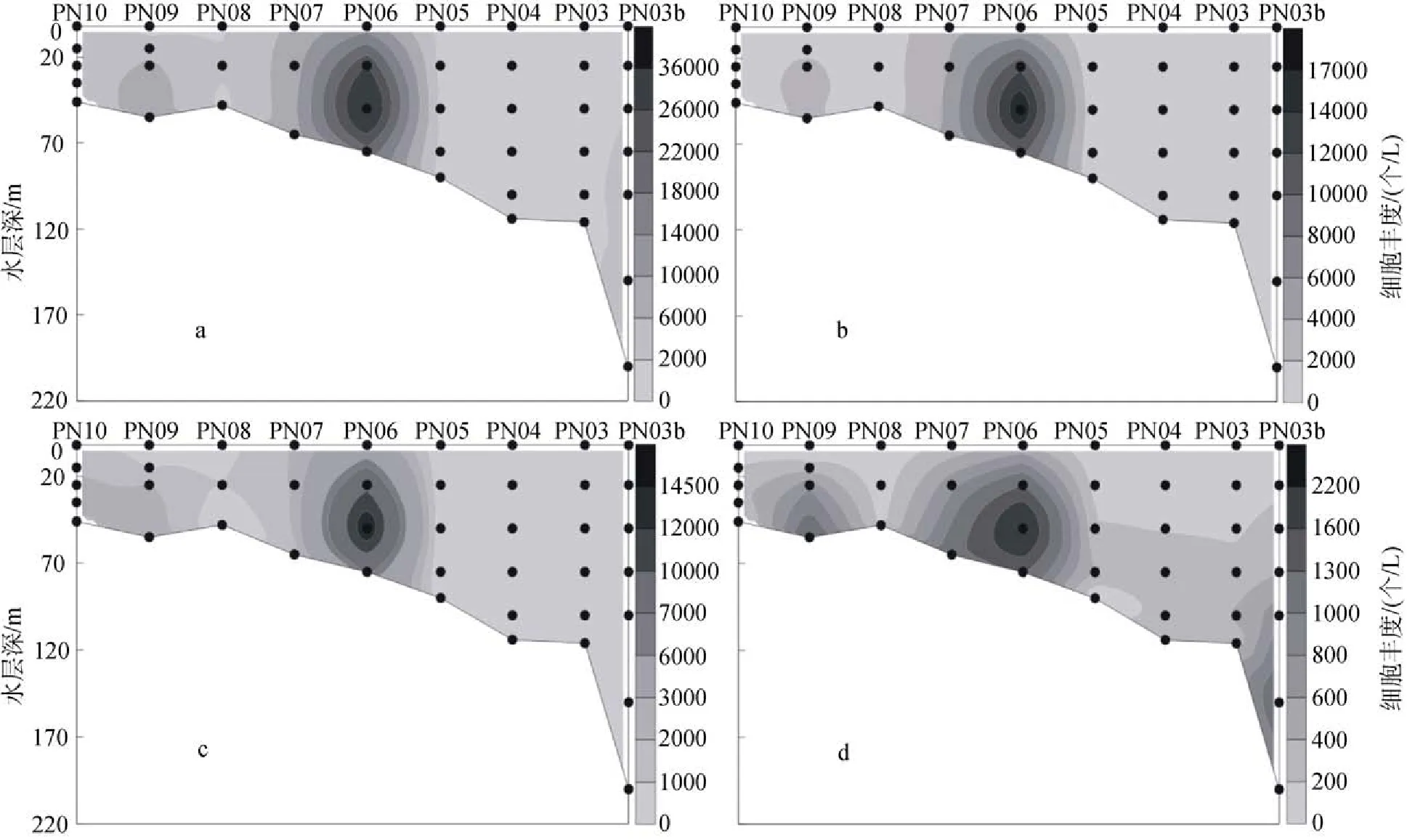
圖11 2009年冬季黃東海海域斷面3顆石藻細胞豐度分布Fig. 11 Vertical distribution of coccolithophore abundance in section 3 in the winter of 2009
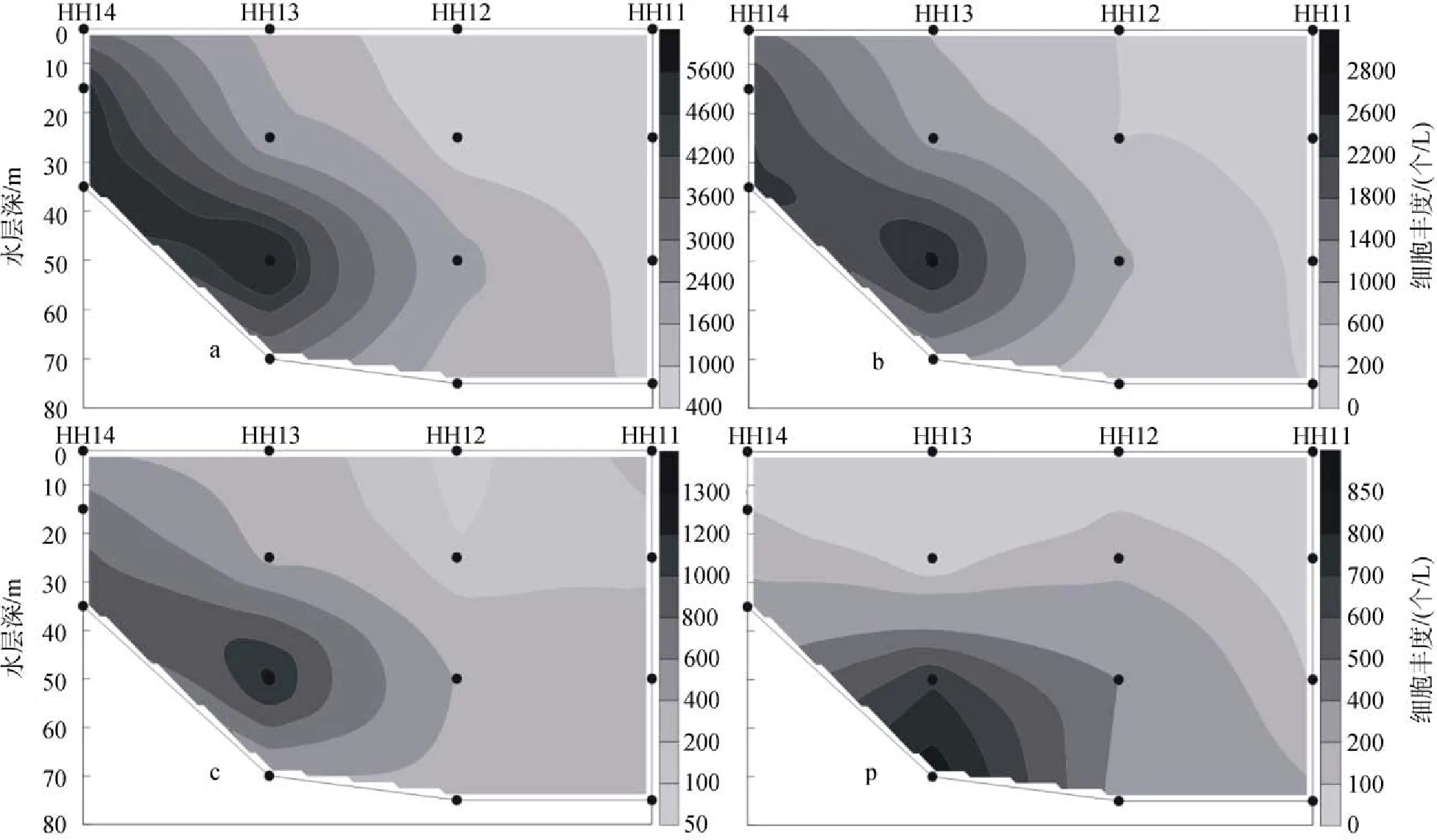
圖12 2009年冬季黃東海海域斷面4顆石藻細胞豐度分布Fig. 12 Vertical distribution of coccolithophore abundance in section 4 in the winter of 2009
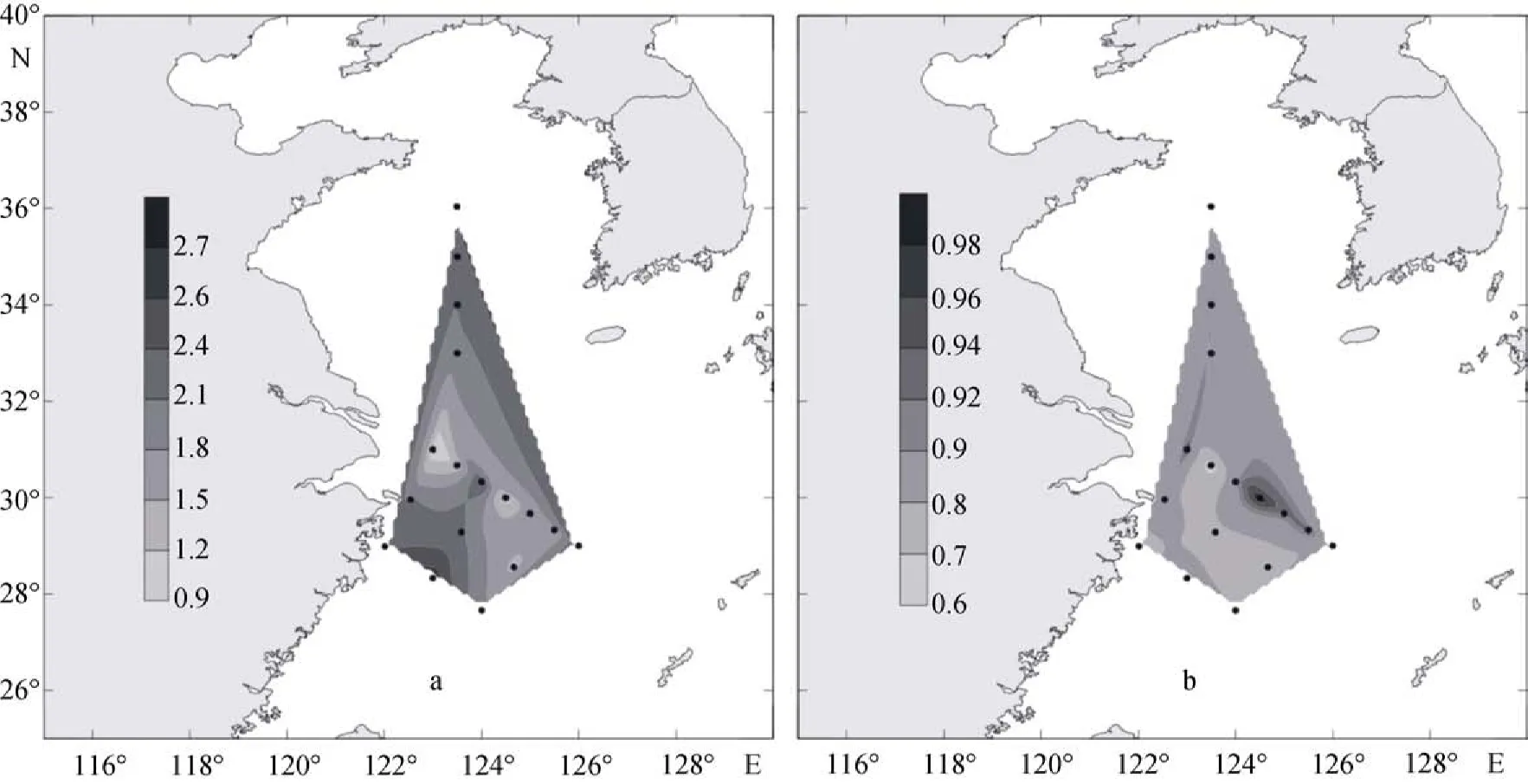
圖13 2009年夏季黃東海海域表層多樣性和均勻度的分布Fig. 13 Distribution of Shannon–Wiener diversity index and Pielou evenness index in surface water of the survey area in thesummer of 2009
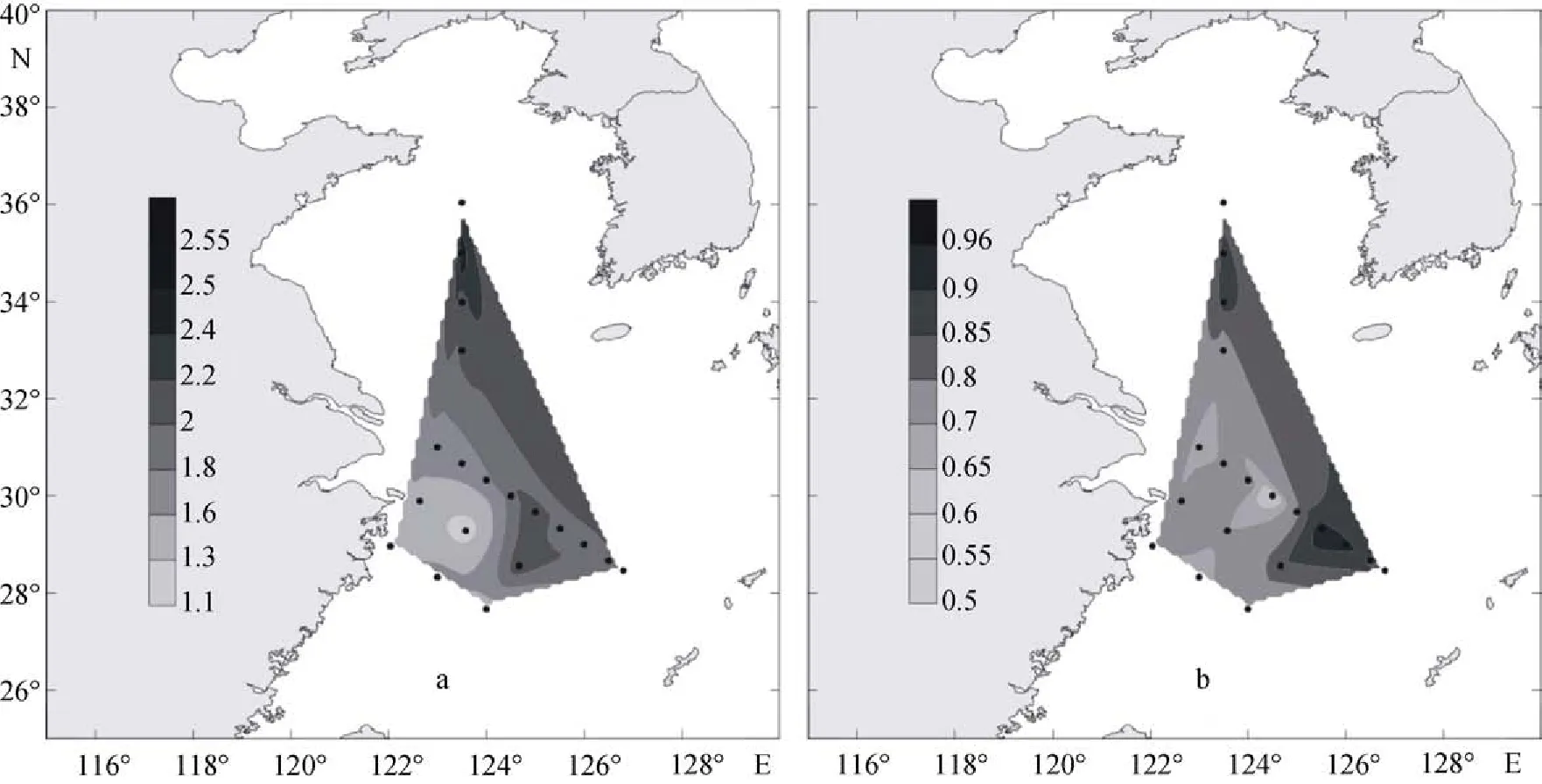
圖14 2009年冬季黃東海海域表層多樣性和均勻度的分布Fig. 14 Distribution of Shannon–Wiener diversity index and Pielou evenness index in surface water of the survey area in thewinter of 2009
夏季調查區表層的鹽度值為22.39~33.69, 平均值為29.96。鹽度的高值區出現在調查海域的東部,最高值出現在PN04站, 達到33.69。而在南部的DH35站, 鹽度達到最低值, 數值為22.39。從圖15b中可以看出, 夏季黃東海表層鹽度受長江沖淡水的影響較為顯著, 近岸表層鹽度較低, 沿沖淡水方向至外海遞增。
2009年冬季, 黃東海海域調查區的表層溫鹽分布如圖16a和圖16b所示。調查區的表層溫度值為7.63~22.87℃ , 平均值為15.73℃。溫度的高值區出現在調查區的東南部, 最高值出現在PN03b站, 數值為22.87℃。同時, 在最靠北的HH11站, 溫度達到最低值, 為7.63℃。從圖16a中可以看出, 冬季黃東海表層溫度分布趨勢同夏季的相類似, 都是由北向南, 由近岸到遠海, 溫度值越來越高。
冬季調查區表層的鹽度值為27.06~34.58, 平均值為33.24。鹽度的高值區出現在調查海域的東部, 最高值出現在PN03站, 達到34.58。而在靠近杭州灣的DH21 站, 鹽度達到最低值, 數值為27.06。從圖16b中可以看出, 冬季黃東海表層鹽度依舊受到內陸徑流的影響, 在近岸表層鹽度較低, 沿沖淡水方向至外海遞增。
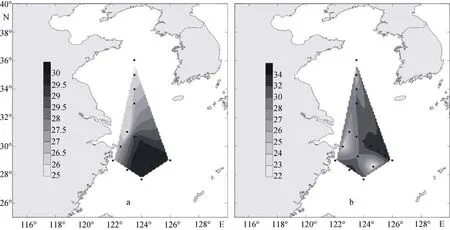
圖15 夏季調查區表層溫鹽分布Fig. 15 Distributions of temperature and salinity in surface water of the survey area in the summer of 2009
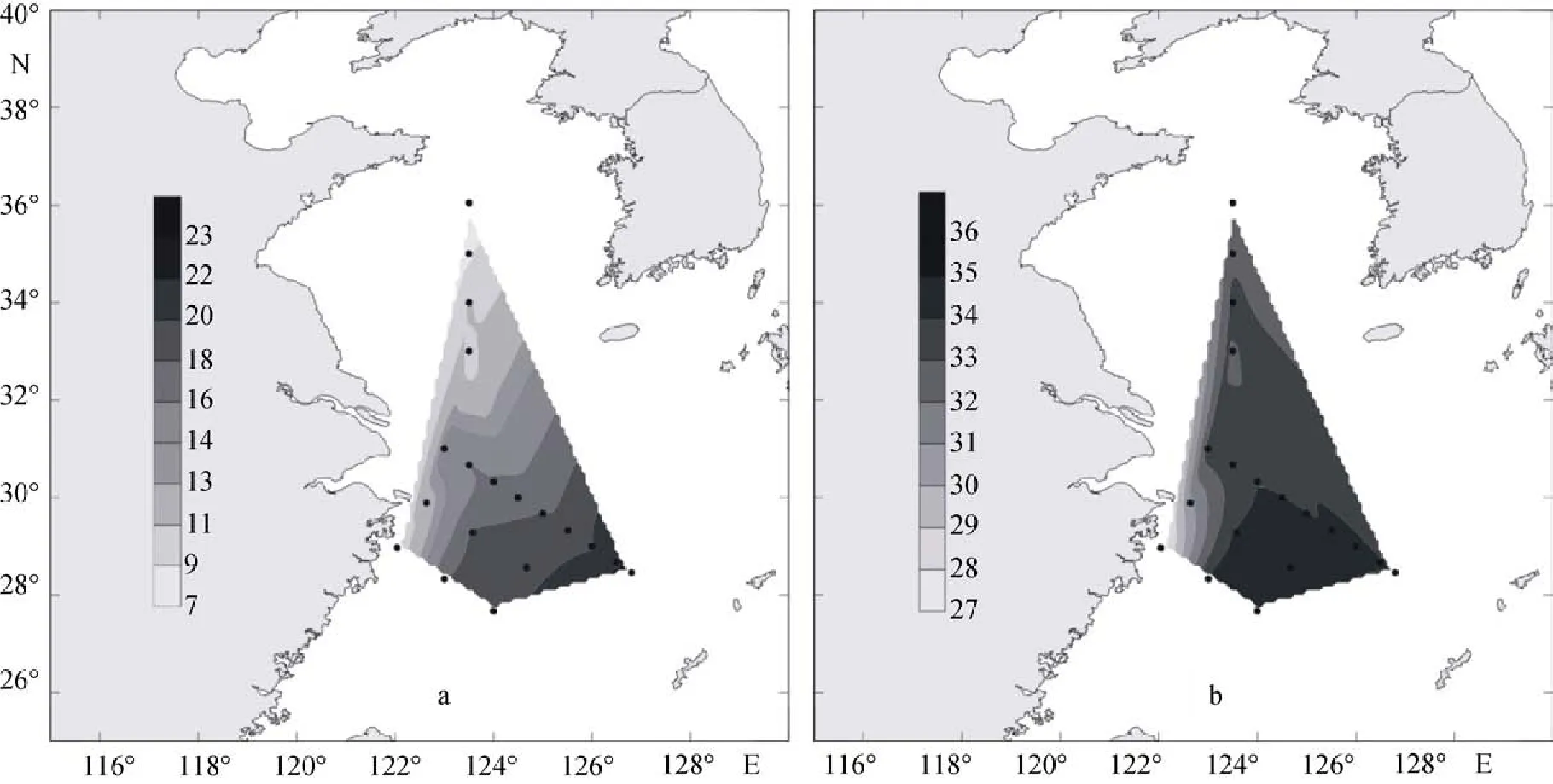
圖16 冬季調查區表層溫鹽分布Fig. 16 Distribution of temperature and salinity in surface water of the survey area in the winter of 2009
3 討論
本文主要研究了中國黃東海海域2009年夏季(7 月20日至9月1日)與冬季(12月23日至2月5日)兩個季度月的顆石藻群落與分布。
2009年夏季, 調查區共鑒定顆石藻21種, 其中絕大多數為異晶顆石藻, 而少數為同晶顆石藻。主要優勢種為赫氏艾密里藻(E. huxleyi)、大洋橋石藻(G. oceanica)、纖細傘球藻(U. tenuis)、深水花球藻(F. profunda)、卡特旋筏球藻(H. carteri)以及希布格臍球藻(U. sibogae)。顆石藻的細胞豐度為0.23×103~17.62× 103個/L, 平均值為2.84×103個/L。這些優勢種的分布高值區一般都出現在調查海域的南部及其周邊水域。這是由于夏季黃東海顆石藻的分布不光受到光和營養鹽的影響, 其受到南方來的黑潮暖流以及北方的黃海冷水團的交互影響也十分明顯[12], 同時,作為中國沿海最大的沖淡水徑流—長江, 其對顆石藻分布的影響也是十分顯著的。
2009年冬季, 調查區共發現顆石藻20種, 與夏季相同, 發現的顆石藻大多數為異晶顆石藻, 只有少數為同晶顆石藻。主要優勢種為赫氏艾密里藻(E. huxleyi)、大洋橋石藻(G. oceanica)、深水花球藻(F. profunda)、纖細傘球藻(U. tenuis)、美麗條結藻(S. pulchra)和希布格臍球藻(U. sibogae)。顆石藻的細胞豐度為0.12×103~35.35×103個/L, 平均值為3.84×103個/L。與夏季相比, 顆石藻的豐度值有很大的提高, 但是優勢種的種類變化不大。冬季顆石藻細胞豐度高值區出現在中部及東部海域, 其豐度值覆蓋的范圍要比夏季寬廣許多, 但是均勻度分布卻不如夏季顯著。
與夏季不同的原因主要是由于冬季調查海域受到強烈季風的作用, 黃東海陸架區的上升流作用效果顯著, 上升流在某一區域將海水底部的營養鹽輸送到上層水體[13], 同時, 溫暖的黑潮水也在很大程度上改變著顆石藻的生存環境, 它們的共同作用給予了顆石藻生長的必要營養物質和生長環境, 從而使其產生較高的細胞豐度。
我國黃東海陸架寬廣、水系眾多, 與西、北太平洋相接, 環境條件復雜而多變。發源于北赤道流的黑潮與東海陸架水相互作用, 其混合水形成獨特的棲息環境, 對于顆石藻的快速生長與繁殖具有重要作用[14]。今后對顆石藻的研究除了走航式觀測外, 還應加強定點調查, 尤其是通過連續站觀測其晝夜變化,并且關注其季節動態。此外, 由于顆石藻存在不同的生態適應策略, 其在物種組成、多樣性以及區域分布等方面也具有更加多樣化的特征。因此, 在緯度較高的海區, 如渤海, 也應該開展顆石藻的調查工作, 以期更加全面地了解顆石藻在中國海區的分布狀況。
[1] Billard C and Inouye I. What is new in coccolithophore biology? In: Coccolithophore: from molecular processes to global impact. Thierstein H R, Young J R, eds. Berlin: Springer-Verlag Berlin, 2004. 35-37.
[2] 孫軍.球石藻的有機碳泵和碳酸鹽反向泵. 地球科學進展, 2007, 22(12): 1231-1239.
Sun Jun.Organic carbon pump and carbonate counter pump of living coccolithophorid. Advances in earth science, 2007, 22(12): 1231-1239.
[3] Paasche E. A review of the coccolithophorid Emiliania huxleyi(Prymnesiophyceae), with particular reference to growth, coccolith formation, and calcification photosynthesis interactions. Phycologia, 2002, 40(6): 503-529.
[4] Bollmann J, Cortés M Y, Haidar A T. Techniques for quantitative analyses of calcareous marine phytoplankton. Marine Micropaleontology, 2002, 44: 163-185.
[5] 國家質量技術監督局. GB/T 12763. 4-91. 海洋調查規范—海水化學要素觀測. 北京: 中國標準出版社, 1991: 1-54.
National quality and Technical Supervision Bureau of.GB/12763.4-91. T oceanographic survey standard seawater chemical element observation. Beijing: China Standard Press, 1991: 1-54.
[6] The state bureau of quality and technical supervision. GB/T 12763. 4-91. Ocean survey specification—Water chemical elements.Beijing: Standards Press of China, 1991: 1-54
[7] Heimdal B R. Modern coccolithophorid. In: Tomas C R, eds. Identifying Marine Phytoplankton. United Kingdom: Academic Press, 1993. 147-247.
[8] Jordan R W, Kleijne A. A classification system for coccolithophores. In: Winter A, Siesser W G, eds. Coccolithophores. United Kingdom: Cambridge University Press, 1994. 83-105.
[9] Winter A and Siesser G. Atlas of coccolithophores. In: Winter A and Siesser W G, eds. Coccolithophores. Cambridge: Cambridge University Press, 1994. pp: 107-159.
[10] Honjo S. Coccoliths production, transportation, sedimentation. Marine Micropaleontology, 1976, 1: 65-79.
[11] 鄒娥梅, 郭炳火, 湯毓祥等. 南黃海及東海北部夏季若干水文特征和環流分析[J]. 海洋與湖沼, 2001, 32(3): 340-348.
Zou Emei, Guo Binghuo, Tang Yuxiang, et al.A number of hydrological characteristics and circulation analysis in Southern Yellow Sea and Northern East China Sea in summer[J]. Oceanologia et Limnologia Sinica, 2001, 32(3): 340-348.
[12] Yang T N and Wei K Y. How many coccoliths are therein a coccosphere of the extant coccolithophorids? A compilation. Journal of Nannoplankton Research, 2003, 25: 7-15.
[13] 于非, 張志欣, 刁新源等. 黃海冷水團演變過程及其與鄰近水團關系的分析[J]. 海洋學報, 2006, 28(5): 26-34.
Yu Fei , Zhang Zhixin, Diao Xinyuan, et al.Analysis of evolution of the Huanghai Sea Cold Water Mass and its relationship with adjacent water[J]. Acta Oceanologica Sinica, 2006, 28(5): 26-34.
[14] 鐘權偉. 南海北部海域鈣板金藻群落的時空變異[D].臺灣: 中山大學海洋生物研究所, 2005.
Zhong Quanwei. The Spatial and temporal variation of coccolithophorids in the north of the South China Sea. National sun yat-sen university institute of Marine organisms, 2005.
[15] Yang T N, Wei K Y, Chen M P. Summer and winter distribution and malformation of coccolithophores in the East China Sea. Micropaleontology, 2004, 50(s1): 157~170.
(本文編輯: 梁德海)
Distribution of coccolithophores in the Yellow Sea and East China Sea in summer and winter
ZHANG Jian, LI Jia-rui, JI Da-wei, ZHAI Wei-kang, WANG Yuan-jun
(National Marine Data and Information Service, Tianjin 300171, China)
Jul., 5, 2015
coccolithophores; community and distribution; Yellow Sea and East China Sea; summer and winter
A community-based survey of the monthly distribution of coccolithophores in the Yellow Sea was conducted in two seasons of 2009: summer (July 20–September 1) and winter (December 23–February 5). In the summer of 2009, 21 species of coccolithophores were identified in the survey area of the Yellow Sea, which were dominated by Emiliania huxleyi, Gephyrocapsa oceanica, Umbellosphaera tenui, and Florisphaera profunda. The abundance of coccolithophores was between 0.23×103and 17.62×103cells/L, with an average of 2.84×103cells/L. In winter 2009, 20 species of coccolithophores were identified in the survey area of the Yellow Sea, which were dominated by E. huxleyi, G. oceanica, F. profunda, and U. tenuis. The abundance of coccolithophores was between 0.12×103and 35.35×103cells/L, with an average of 3.84×103cells/L. The distribution of coccolithophores in the Yellow Sea and East China Sea were systematically studied to determine the community distribution of coccolithophores in the Yellow Sea and East China Sea.
Q948.1
A
1000-3096(2016)02-0041-14
10.11759/hykx20140710003
2015-07-05;
2015-09-11
國家重點基礎研究發展規劃項目(2006CB400605)
張健(1986-), 男, 山東青島人, 工程師, 碩士, 主要從事海洋生態學研究, E-mail: 889900hhhhhh@163.com
